
牡丹试管苗与扦插苗生根过程中相关酶活性的变化
2014-03-26 07:39王照路王雪玲何松林
西北农林科技大学学报(自然科学版) 2014年10期
王 政,王照路,申 萍,王雪玲,何松林
(河南农业大学 林学院,河南 郑州 450002)
牡丹(PaeoniasuffruticosaAndr.)别名木芍药、富贵花、鼠姑,为芍药科芍药属宿根木本花卉,是我国著名的观赏植物与药用植物[1]。其野生种主要分布于陕西、河南、四川、西藏等地,花姿典雅,雍容华贵,被誉为“花中之王”,是我国的传统名花,常采用播种、分株和嫁接等传统方法繁殖。但在实际生产中,传统繁殖方法的繁殖系数低,生长速度慢,难以满足牡丹苗木产业化和商品化生产的需要。而组织培养技术能大大缩短牡丹的繁殖周期,繁殖系数高,利于大规模生产。但牡丹试管苗根系发生困难、根系质量不高成为生根培养的主要瓶颈之一[2]。
无根试管苗与扦插苗生根过程相似,在其不定根的形成过程中,均伴随着插穗或试管苗茎基部组织进行细胞脱分化、根原基形成以及酶活性变化等一系列生理活动。有研究表明,牡丹试管苗由于根系由基部愈伤组织形成且无维管束与茎中木质部连接,导致其根系质量不高且成活率低[3];扦插苗不定根中的维管束与插穗木质部直接相连,根系质量好且移栽成活率较高。因此,研究牡丹试管苗和扦插苗生根过程中生理指标的变化规律具有重要意义。研究表明,过氧化物酶(POD)、多酚氧化酶(PPO)和吲哚乙酸氧化酶(IAAO)等酶活性的变化与植物生根过程密切相关。PPO对苹果[4-5]、梨[5]插穗和葡萄[6]试管苗生根有影响;POD与乐昌含笑[7]试管苗生根过程中根原基的形成和发育密切相关,与垂丝海棠、楸子[8]插穗生根过程中愈伤组织的形成有关;在山杏[9]插穗生根过程中IAAO活性与不定根的形成有关。目前,有关牡丹生根过程中POD、PPO、IAAO等酶活性的研究,主要集中在试管苗生根过程中的变化规律[2-3,10-11]方面,而对扦插苗和试管苗生根前后酶活性变化情况的研究尚未见报道。因此,本试验拟通过对比大田扦插苗和组培试管苗生根过程中上述3种酶活性的变化规律,探讨酶活性变化与牡丹生根的关系,分析牡丹试管苗生根难的可能内在原因,以期为提高牡丹试管苗生根率提供理论基础和技术依据。
1 材料与方法
1.1 供试材料
于2013-01-25,在洛阳土桥花木种苗有限公司牡丹苗木基地采集牡丹品种‘凤丹白’鳞芽,取样植株生长状况良好,无病虫害,常规田间管理。将所采牡丹鳞芽灭菌后接种到含MS+6-BA 0.5 mg/L+NAA 0.5 mg/L+蔗糖30 g/L+琼脂7 g/L (pH=5.8)的固体培养基上,在常规条件下[温度(24±1) ℃,光照强度40 μmol/(m2·s),光照时间12 h/d]进行诱导培养,培养30 d后获得供试试管苗。
牡丹品种‘凤丹白’插穗采自河南农业大学三区实验基地,母株为3年龄。插穗为当年生半木质化嫩枝,长10~12 cm,基部直径0.5~0.8 cm。每条插穗留2~3个芽,剪去下部叶片,将插穗下端剪成平滑的斜面,上端留2~3片叶片,并剪去叶片的 1/3 以减少蒸腾,备用。
1.2 试验方法
1.2.1 组织培养诱导生根 将供试牡丹试管苗在超净工作台上,接种到含木本植物培养基(WPM)+IBA 4.0 mg/L+聚乙烯吡咯烷酮(PVP)1.0 g/L+Vc 50 mg/L+植物凝胶(Gellan gum)2.0 g/L+蔗糖30 g/L (pH=5.8)的固体培养基中,进行生根培养,每瓶接种1株。
1.2.2 扦插生根 将修剪好的牡丹插穗用1 g/L IBA处理,然后扦插在V(沙)∶V(土)=1∶1的混合基质中,置于盆中培养,插入深度为插穗长的1/2~2/3,压实基质,浇透水,之后保持湿润。
1.3 测定指标及方法
贺丹等[3]、徐盼盼[10]对牡丹试管苗生根过程的观察发现,其生根时间为诱导培养后3~15 d;本研究预试验发现,牡丹扦插苗在第18天有根原基形成,因此将试验取材时间定为0~21 d。于生根培养的0,1,2,3,4,5,7,9,12,15,18,21 d取材,试管苗随机取10株,取其茎基部;扦插苗随机取10株,取插穗基部1~2 cm;各自混匀后分别测定其POD、PPO、IAAO酶活性,均重复3次。
POD活性测定参照李合生[12]的方法;PPO活性测定参照朱广廉等[13]的方法;IAAO活性测定参照张志良[14]的方法。
1.4 数据统计与分析
试验数据采用Excel软件和DPS 3.01对数据进行处理分析;采用邓肯氏新复极差法(SSR法)检测其差异显著性,显著水平为P<0.05。
2 结果与分析
2.1 试管苗与扦插苗生根过程中过氧化物酶(POD)活性的变化
从图1可以看出,生根过程中,牡丹试管苗的POD活性在0~3 d整体呈下降趋势,其中第3天降到最低值(6.67 U/g);在3~7 d呈上升趋势,并在5 d达到第1峰值,然后在第9天下降至18.67 U/g;9~12 d急剧上升,于第12天达到第2峰值,12~15 d下降,而后呈上升趋势;且试管苗的POD活性始终显著高于扦插苗。与试管苗相比,扦插苗的POD活性变化趋势较平缓,其中0~2 d呈上升趋势,第3天下降至1.67 U/g,第4天上升至最大值(20.00 U/g)后下降,在5~15 d整体呈上升趋势,于第15天达到第2峰值后下降,在18~21 d又呈上升趋势。试管苗与扦插苗POD活性均在第3天降到最低值,第1个峰值出现时间分别为5和4 d,分别于12和15 d达到第2个峰值;第2个低值出现时间分别是第9和第5天。
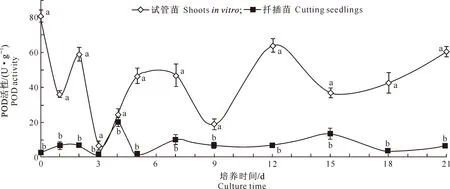
图1 牡丹试管苗和扦插苗生根过程中过氧化物酶(POD)活性的变化
2.2 试管苗与扦插苗生根过程中多酚氧化酶(PPO)活性的变化
在本试验中,牡丹品种‘凤丹白’无根试管苗与扦插苗生根过程中多酚氧化酶(PPO)活性的变化结果见图2。
如图2所示,转入生根培养基后,牡丹试管苗的PPO活性在0~1 d迅速上升,在第1天达到最高峰(21.0 U/g),1~5 d整体呈下降趋势,于第5天降到最低值(1.0 U/g),5~12 d呈波浪上升趋势,于第12天达到峰值后下降,18~21 d再次上升;扦插苗的PPO活性在0~3 d上升后下降,于第4天降到1.0 U/g,4~9 d逐步上升,第9天出现第2个峰值,9~15 d下降后再次上升,于第21天达到最大值(7.0 U/g)。整个生根过程中试管苗与扦插苗PPO活性在第1,2和12天差异显著。与试管苗相比,扦插苗PPO活性第1个峰值的出现时间推迟2 d,第2个峰值提前3 d;试管苗与扦插苗PPO活性分别在5和4 d降到最低值。
2.3 试管苗与扦插苗生根过程中吲哚乙酸氧化酶(IAAO)活性的变化
由图3可知,在生根培养过程中,试管苗IAAO活性变化较小,在0~7 d呈波浪上升趋势,在第7天达到峰值(33.47 U/g)后下降并趋于平稳,18~21 d上升,于第21天达到最大值(36.59 U/g);扦插苗在0~5 d呈下降趋势,第5天降至27.82 U/g,然后上升,于第9天达到第1个峰值(50.04 U/g),9~15 d下降,于第15天降到最低值(27.23 U/g),15~21 d迅速上升,第21天达到最大值(67.39 U/g)。在生根过程中,除第4天外,二者IAAO活性均达显著差异,扦插苗IAAO活性变化幅度较试管苗大。试管苗与扦插苗IAAO活性分别在第7和第9天出现第1个峰值,试管苗较扦插苗提前2 d,二者均在21 d达到最大值;试管苗在第3天出现低值,扦插苗在第5天出现低值,第15天降到最低。
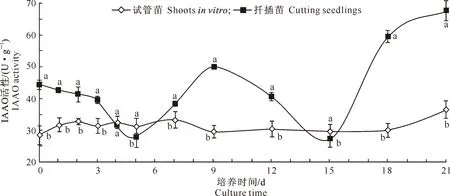
图3 牡丹试管苗和扦插苗生根过程中吲哚乙酸氧化酶(IAAO)活性的变化
3 结论与讨论
本试验探讨了牡丹品种‘凤丹白’试管苗与扦插苗生根过程中POD、PPO、IAAO 3种酶活性的变化,结果表明,在生根过程中,牡丹试管苗POD活性呈“下降-上升-下降-上升”的趋势,PPO和IAAO活性均呈“上升-下降-上升”趋势;而扦插苗POD和PPO活性均呈“上升-下降-上升”趋势,IAAO活性呈“下降-上升-下降-上升”趋势;牡丹试管苗POD活性在第3天降到最低值,第1个峰值出现时间较扦插苗推迟1 d;PPO活性第1个峰值较扦插苗提前2 d,最低值推迟1 d;IAAO活性第1个峰值出现时间较扦插苗提前2 d,均在第21天达到最大值,第1个低值出现时间较扦插苗提前2 d。
近年来,有研究认为,POD、PPO及IAAO活性与植物不定根的发生和生长均有着密切关系,其变化因生根时期不同而异[15]。相关研究表明,在不定根诱导期和表达期,POD 活性升高是有生根能力的标志[16-17]。贺丹等[3,11]、徐盼盼[10]对牡丹试管苗进行生根细胞学观察发现,根原基的诱导发生于3~5 d,9~15 d根原基进一步分化,逐渐突破表皮。本试验发现,牡丹试管苗POD活性在生根培养的3~5 d迅速上升后下降,9~12 d继续上升。由此推测,第3天开始POD活性升高,可能是因为产生了某些物质进而促进了试管苗不定根的发生,9~12 d POD活性迅速升高有利于根原基的进一步分化;15~21 d POD活性继续升高,可能与茎基部木质素的成熟度有关[18];扦插苗POD活性变化较平缓, 3~4 d上升达最大值后下降,5~15 d整体呈上升趋势,于第15天达到峰值后下降,18~21 d再次升高,这与宋金耀等[19]的研究结果类似,可能起始期高活性的POD对不定根的发生起诱导作用。在生根过程中,试管苗与扦插苗POD活性均在第3天降到最低值,而后呈上升趋势,可能有利于根原基的诱导。试管苗POD活性第2个峰值出现在第12天,而扦插苗出现在第15天,较试管苗推迟3 d,可能与扦插苗根系发生较晚有关。
PPO是一种含铜酶,可能会催化酚类物质与IAA形成一种“IAA-酚酸复合物”,这种复合物可能是一种生根辅助因子,可促进不定根的形成[17]。另外,PPO活性还与植物组织培养外植体褐变有密切关系,接种时由于切割作用,PPO作用于天然底物酚类物质而形成褐色物质醌,导致外植体的褐化,直接影响组培苗生根及正常生长[6,20],因此,PPO对生根具有双重效应。本试验中,试管苗0~2 d PPO活性变化剧烈,第1天达到最大值,可能是由于切割作用,PPO催化多酚类物质发生褐化,导致PPO活性迅速升高,2~12 d呈波浪上升趋势,可能有利于“IAA-酚酸复合物”的形成以促进根原基的诱导;扦插苗PPO活性在0~3 d上升后下降,从第4天开始继续上升,于第9天达到峰值,可能有利于根原基的诱导,在15~21 d逐步升高,于第21天达到最大值,可能是因为催化生成的“IAA-酚酸复合物”增多,从而促进根的形成。在整个生根过程中,与试管苗相比,扦插苗PPO活性第1个峰值出现的时间推迟2 d,第2个峰值提前3 d,2个峰值均低于试管苗,第21天扦插苗PPO活性达到最大值且较试管苗高,可能与扦插苗根系发生较试管苗晚有关。
IAAO可以降解植物体内的吲哚乙酸(IAA),调节植物体内的IAA水平,从而影响试管苗生根[21-22]。Gaspar等[23]试验表明,离体生根过程中,高活性的IAAO使内源IAA水平降低,是生根诱导期的特点之一。低浓度的IAA有利于诱导生根,之后在表达期要求较高浓度的IAA以促进根的生长。本试验中,试管苗IAAO活性变化不显著,在0~7 d呈波浪上升式变化,第7天出现峰值,可能有利于降低内源IAA水平从而促进根原基的诱导,之后变化趋于平缓,于第21天达到最大值,可能与本研究观察到该时期的侧根形成现象有关。扦插苗IAAO活性在0~5 d下降,5~9 d呈上升趋势,第9天出现第1个峰值。有研究表明,扦插后前10 d是根的诱导阶段[24],期间IAAO活性升高可能会促进IAA分解,有利于根原基诱导生根;9~15 d逐渐下降至最低值,可能有利于愈伤组织的形成,随后迅速上升,第21天达到最大值,其变化趋势与宋金耀等[19]研究的月季、葡萄等植物在扦插生根过程中IAAO活性的变化趋势相似。在生根过程中,除第4、5和15天外,扦插苗IAAO活性均较试管苗高,且扦插苗IAAO活性第1个峰值和低值的出现时间均较试管苗推迟2 d,可能是扦插苗根系发生晚的原因之一。扦插苗IAAO活性变化幅度较大,在根的诱导阶段(扦插后0~10 d)先下降后迅速上升,有利于降低内源IAA水平,从而促进根原基的诱导;而试管苗IAAO活性则变化较平缓,可能在调节内源IAA水平方面作用不显著。
本研究发现,与试管苗相比,牡丹扦插苗生根过程中POD、PPO及IAAO酶活性的一些极值出现时间均出现推迟现象,这与扦插苗生根时间较晚有关,但其作用机理还有待于进一步研究。
[参考文献]
[1] 包满珠.花卉学 [M].2版.北京:中国农业出版社,2003:215.
Bao M Z.Floriculture [M].2nd ed.Beijing:China Agriculture Press,2003:215.(in Chinese)
[2] 李 航,王永伟,何松林.牡丹试管苗生根研究进展和展望 [J].安徽农业科学,2007,35(12):3499-3513.
Li H,Wang Y W,He S L.Research progress and prospect of root-inducing ofPaeoniasuffruticosa[J].Journal of Anhui Agri Sci,2007,35(12):3499-3513.(in Chinese)
[3] 贺 丹,王 政,何松林.牡丹试管苗生根过程解剖结构观察及相关激素与酶变化的研究 [J].园艺学报,2011,38(4):770-776.
He D,Wang Z,He S L.Adventitious root generating process and hormone and enzyme changesinvitroPaeoniasuffruticosa[J].Acta Horticulturae Sinica,2011,38(4):770-776.(in Chinese)
[4] Bassuk N L,Hunter L D,Howard B H.The apparent involvement of polyphenol oxidese and phloridzin in the production of apple rooting cofactors [J].Hortic Sci,1981,56:313-315.
[5] Poapst P A,Durkee A B.Root differentiating properties of so-me simple aromatic substances of the apple and pear fruit [J].J Hort Sci,1967,42:429-438.
[6] 宋士任,王 华.葡萄多酚含量和多酚氧化酶(PPO)活性与组培苗生根关系的初步研究 [J].中国农学通报,2005,21(9):70-73.
Song S R,Wang H.The effect of polyphenol content and polyphenol oxidase activity on rooting of tissue cultured seedlings inVitisL. [J].Chinese Agricultural Science Bulletin,2005,21(9):70-73.(in Chinese)
[7] 吴秋峰,金 玲,吴月燕.乐昌含笑组织培养根的诱导及其POD和PPO活性的变化 [J].科技通报,2009,25(4):460-464.
Wu Q F,Jin L,Wu Y Y.Activity changes of POD and PPO during root inducing in tissue culture ofMicheliachapensisDandy [J].Bulletin of Science and Technology,2009,25(4):460-464.(in Chinese)
[8] 许晓岗,丁芳芳,童丽丽.垂丝海棠、楸子扦插生根的生理机制 [J].东北林业大学学报,2013,41(5):91-97.
Xu X G,Ding F F,Tong L L.Basic physiological indices in cuttings ofMalushallianaandMalusprunifolia[J].Journal of Northeast Forestry University,2013,41(5):91-97.(in Chinese)
[9] 董胜君,刘明国,戴 菲,等.2种激素处理的山杏插穗生根过程中营养物质及酶活性的变化 [J].西部林业科学,2012,41(6):26-30.
Dong S J,Liu M G,Dai F,et al.Variation of nutrients and enzyme activities inArmeniacasibiricasoftwood cuttings during adventitious root formation under treatments of two hormones [J].Journal of West China Forestry Science,2012,41(6):26-30.(in Chinese)
[10] 徐盼盼.环境因子对牡丹试管苗生根的影响 [D].郑州:河南农业大学,2011.
Xu P P.The effect of environmental factor on rooting culture ofPaeoniasuffruticosainvitro[D].Zhengzhou:Henan Agricultural University,2011.(in Chinese)
[11] 贺 丹.牡丹试管苗生根调控研究 [D].郑州:河南农业大学,2009.
He D.The control on rooting culture ofPaeoniasuffruticosainvitro[D].Zhengzhou:Henan Agricultural University,2009.(in Chinese)
[12] 李合生.植物生理生化实验原理和技术 [M].北京:高等教育出版社,2003:164-165.
Li H S.Principle and technique of plant physiological and biochemical experiment [M].Beijing:Higher Education Press,2003:164-165.(in Chinese)
[13] 朱广廉,钟海文,张爱琴.植物生理实验 [M].北京:北京大学出版社,1990:37-39.
Zhu G L,Zhong H W,Zhang A Q.Principle of plant physiological experiment [M].Beijing:Beijing University Press,1990:37-39.(in Chinese)
[14] 张志良.植物生理学实验指导 [M].2版.北京:高等教育出版社,1990.
Zhang Z L.Principle of plant physiological experiment guidance [M].2nd ed.Beijing:Higher Education Press,1990.(in Chinese)
[15] 扈红军,曹帮华,尹伟伦,等.不同处理对欧榛硬枝扦插生根的影响及生根过程中相关氧化酶活性的变化 [J].林业科学,2007,43(12):70-75.
Hu H J,Cao B H,Yin W L,et al.Effects of different treatments on hardwood-cutting rooting and related oxidase activity changes during rooting ofCorylusavellana[J].Scientia Silvae Sinicae,2007,43(12):70-75.(in Chinese)
[16] Moncousin C,Gaspar T H.Peroxidase as a marker for rooting improvement ofCynarascolymusL.culivatedinvitro[J].Biochemie und Physiologieder Pflanzen,1983,178:263-271.
[17] Haissig B E.Influence of auxins and auxin synergists on adventitious root primordium initiation and development [J].NZ J For Sci,1974,4:311-323.
[18] Conzalez A,Tames S R,Rodriguez R.Ethylene in relation to protein,peroxidase and polyphenol oxidase activities during rooting inhazelnut cotyledons [J].Physiol Plant,1991,83:611-620.
[19] 宋金耀,宋 刚,李 辉,等.几种园艺植物扦插生根过程中生化指标的变化 [J].江苏农业科学,2010(3):211-214.
Song J Y,Song G,Li H,et al.The changes of several horticultural plants physiological index during rooting process [J].Journal of Jiangsu Agri Sci,2010(3):211-214.(in Chinese)
[20] 崔堂兵,郭 勇,张长远.植物组织培养中褐变现象的产生机理及克服方法 [J].广东农业科学,2001(3):16-18.
Cui T B,Guo Y,Zhang C Y.Occurring mechanism and overcoming ways of browning in plant tissue culture [J].Journal of Guangdong Agri Sci,2001(3):16-18.(in Chinese)
[21] Jackson M B.New root formation in plant and cuttings [M].Lancaster:Martinus Nijhoff Publishers,1986:223-253.
[22] Gebhardt K.Activation of indole-3-acetic acid oxidase from ho-rseradish andPrunusby phenols and H2O2[J].Plant Growth Reg,1982,1:73-84.
[23] Gaspar T,Kevers C,Hausman J F,et al.Practical uses of peroxidase activity as a predictive marker of rooting performance of micropropagated shoot [J].Agronomic,1992(12):757-765.
[24] 宋丽红,曹帮华.光叶楮扦插生根的吲哚乙酸氧化酶、多酚氧化酶、过氧化物酶活性变化研究 [J].武汉植物学研究,2005,23(4):347-350.
Song L H,Cao B H.Studies on activities of indoleacetic acid oxidase,polyphenol oxidase and peroxidase in cuttings ofBroussonetiapapyriferaduring rooting process [J].Journal of Wuhan Botanical Research,2005,23(4):347-350.(in Chinese)
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26
河北农机(2020年10期)2020-12-14
计算机应用(2018年10期)2018-11-22
中成药(2018年10期)2018-10-26
华南师范大学学报(自然科学版)(2017年4期)2017-09-11
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
上海农业学报(2017年4期)2017-04-10
好孩子画报(2016年7期)2016-12-12
中国粮油学报(2016年5期)2016-01-23
中小学实验与装备(2014年2期)2014-09-21
