
环境温度对达乌尔黄鼠冬眠模式和能量消耗的影响
2014-05-16 07:28宋士一
沈阳师范大学学报(自然科学版) 2014年3期
于 超,宋士一,彭 霞,杨 明
(1.沈阳师范大学 化学与生命科学学院,沈阳 110034;2.沈阳师范大学 实验教学中心,沈阳 110034)
0 引 言
许多内温动物通过蛰眠(Torper)来度过寒冷的冬季[1],冬眠是蛰眠的一种类型,通常指动物在麻痹或昏睡状态下越冬[2]。恒温动物维持高体温需要消耗大量的能量,而温带和极地的哺乳动物在冬季面临着食物短缺的问题,为了节约能量,有些动物会采取降低代谢率和体温的冬眠策略。然而动物在整个冬眠季节并不是一直都处于蛰眠状态,冬眠期间体温降低,自身代谢率处于较低水平,且这一状态能够维持几天或几周,然后被重复性的复温和简短的持续性产热所打断[3-4],这样动物的整个冬眠季节会被分割成若干个冬眠阵。已有的研究表明,同一物种的冬眠模式可以依据冬眠季节的长短和冬眠阵类型分为深冬眠型、少冬眠型和不冬眠型[5]。深冬眠型长而深的冬眠阵会使动物长时间处于低体温低代谢的状态下,更有利于节约能量[6]。
深冬眠状态下,动物的体温会在一定范围内随环境温度的变化而变化,其冬眠模式也随着温度的改变而改变。所以,环境温度会通过影响深冬眠阶段的体温以及阵间觉醒的频率等参数,进而影响冬季的能量消耗[7]。有报道显示,全球气候变暖,动物园和野外生活的冬眠动物提前出眠,从而对其生存和繁殖造成了严重的影响。但环境温度如何影响动物的冬眠模式和能量消耗尚缺乏系统的研究报道。动物能否适应逐渐变暖的冬季,温暖的冬季会对冬眠动物的越冬有何影响仍有待进一步研究。
达乌尔黄鼠(Spermophilusdauricus)是一种典型的贮脂类小型冬眠哺乳动物,分布于古北界蒙新区。该物种在冬眠前期会在体内大量贮存脂肪,以应对漫长的冬季,入眠阶段是一个从储存能量向消耗能量转变的过程[8]。在入眠前的体重可以达到出眠后的两倍左右。大量的脂肪储备为该物种的冬眠提供了充足的能量。而黄鼠在深冬眠阶段几乎不取食。体内贮存的脂肪是其越冬的主要能源。本文在实验室不同环境温度条件下,通过连续长时间的监测动物的体温[9]和能量收支,记录达乌尔黄鼠在不同环境温度下的冬眠模式和能量消耗,为进一步分析气候变化对动物冬眠的影响有提供支持。
1 材料与方法
1.1 实验动物
实验用达乌尔黄鼠27只,于2011年五月捕自内蒙古通辽(43.9°N,122.5°E),于沈阳师范大学动物房单笼饲养,饲养笼大小48cm×35cm×20cm,用碎刨花做巢材。动物房内自然温度(随外界环境气温波动的范围为16~28℃)和光照,动物喂以沈阳市于洪区前民动物饲料厂生产的标准鼠饲料,水、食自取。9月份,当动物出现蛰眠状态(卷曲身体、体温降低、行动迟缓)时,将动物随机分为3组,分别置于(5±2)℃,(15±2)℃,(25±2)℃下饲养,笼内加适量的棉花作为补充巢材。低温房恒黑(模拟洞穴内条件),动物水、食仍自取,饲料成分不变。
1.2 体温和环境温度的记录
动物移入低温房前半个月腹腔内埋植半导体温度记录元件(Maxim/Dallas公司生产的DS1921LThermochron®iButton,温度记录范围为-40~85℃,灵敏度为0.5℃),每小时记录一次体核温度,以准确测定冬眠季节及其前后体温的变化和冬眠的表达[7,10]。动物术后恢复一周。另取3个iButton,参数设定同上,分别置于3号自封袋内,悬挂于黄鼠越冬的低温房,用来记录不同低温房的环境温度。
1.3 冬眠模式
体温(Tb)<30℃时定义为异温状态,首次记录到Tb<30℃时为动物的入眠时间,最后一次记录到Tb从<30℃回复到≥30℃为动物的出眠时间。冬眠季节即为从入眠到出眠的时间跨度。动物冬眠季节内,所有异温状态的时长之和为异温累积时间。取不同环境温度下达乌尔黄鼠连续且平稳的3个最长冬眠阵,计算连续3个冬眠阵的平均天数作为该温度下冬眠阵的长度。根据冬眠季节的长短和冬眠阵的类型,将冬眠分为深冬眠型、少冬眠型和不冬眠型,深冬眠型冬眠季节长,主要由若干持续多天的深冬眠阵组成;少冬眠型冬眠季节短,冬眠阵中深冬眠阵较少,短冬眠阵和日眠阵较多;不冬眠型个体整个冬季都不表达深冬眠阵和短冬眠阵,只表达日眠阵或一直处于常温产热状态[7]。
1.4 能量消耗的测定
动物冬眠期间的总能量消耗由食物消耗和体脂消耗两部分组成。食物消耗提供的能量通过食物平衡法测定能量收支来计算。体脂消耗提供的能量通过监测动物入眠与出眠的体重差,作为体脂消耗。
1)能量收支测定。用改进的饲养笼法测定动物的能量收支[11]。采用全自动氧弹量热仪(YX-ZR/Q型,长沙友欣实业有限生产公司)测定食物和粪便热值。用电子天平监测动物体重,每周1次。体重的变化为动物体脂的变化。
2)体脂消耗测定。动物入眠和出眠时的体重差当做动物在冬眠过程中的体脂消耗。1g脂肪组织大约含有0.8g脂质(39kJ/g)[12-13]。得出动物脂肪供能公式为:体脂能量=体脂消耗质量×0.8×39(kJ)。
1.5 数据统计
数据统计利用SPSS 13.0软件统计包,进行统计分析。1-Sample-K-S检验后,证明所有用于统计的数据均符合正态分布。黄鼠在冬眠过程中体脂的消耗,冬眠模式的差异,总能量消耗,均采用独立样本T检验(Independent-Samples T test),分析后再采用单因素方差分析(One-way ANOVA)和post hoc多重比较。文中所用数据均以平均值±标准误(Mean±SE)表示。P<0.05为差异显著,P<0.01为差异极显著。
2 结 果
2.1 冬眠模式
1)冬眠类型。深冬眠型、少冬眠型和不冬眠型3种冬眠模式在不同环境温度下的冬眠动物中均有表达。不同的环境温度下的动物通过蛰眠均成功越过冬季,但越冬模式具有一定差异(图1)。5℃环境温度下9只动物全部为深冬眠型(图1a);15℃环境温度下6只动物为深冬眠型(图1b),2只动物为少冬眠型(图1c);25℃环境温度下,7只动物为少冬眠型(图1d),1只动物为不冬眠型(图1e)。

图1 不同环境温度下达乌尔黄鼠的冬眠模式
2)冬眠季节长度。5,15,25℃动物的冬眠季节长度分别为(168.33±6.17)d,(146.75±11.45)d,(88.38±10.39)d(图2),差异显著(F(2,22)=19.431,P=0<0.01)。
3)平均冬眠阵长度。5,15,25℃环境温度下达乌尔黄鼠连续3个冬眠阵长度分别为(27.4±3.3)d,(15.3±1.2)d,(3.88±0.58)d,差异显著(F(2,22)=28.838,P=0<0.01)。
4)异温累积时长。5,15,25℃环境温度下总的异温累积时长分别为(110.45±4.38)d,(78.50±6.00)d,(14.09±3.98)d(图4),显著差异(F(2,22)=94.669,P=0<0.01)。
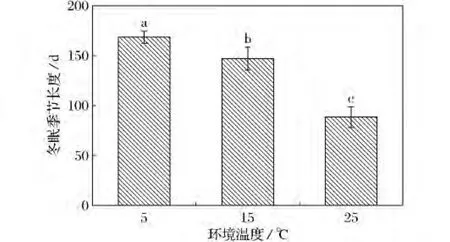
图2 不同环境温度下达乌尔黄鼠冬眠季节的长度

图3 不同环境温度下平均3个冬眠阵长度(Day)

图4 不同环境温度下达乌尔黄鼠冬眠异温累积时长
2.2 冬眠期间摄食量
5,15,25℃动物的摄食量分别为(38.00±5.46)g,(557.70±104.20)g,(913.59±60.45)g(图5),差异显著(F(2,22)=42.525,P=0<0.01)。
2.3 体脂的消耗
在5,15,25℃的环境温度下冬眠季节体脂消耗分别为(83.16±6.38)g,(85.63±15.21)g,(123.55±13.55)g,这些体脂分别为动物提供了(2 594.59±199.06)kJ,(2 671.66±474.55)kJ,(3 854.76±422.38)kJ能量。组间差异显著(F(2,22)=3.408,P=0.05)(图6)。25℃环境下体脂消耗量显著高于5℃(P=0.025<0.05)和15℃(P=0.039<0.05)。
2.4 冬眠期间总能量消耗
5,15,25℃动物在冬眠过程中消耗的总能量分别为(3 722±217.82)kJ,(18 907±2 895.07)kJ,(30 542±2 146.40)kJ(图7),差异显著(F(2,22)=45.198,P=0<0.01)。

图5 不同环境温度下达乌尔黄鼠冬眠季节的摄食量

图6 不同环境温度下达乌尔黄鼠冬眠季节体脂的消耗

图7 不同环境温度下达乌尔黄鼠越冬总的能量
3 讨 论
3.1 环境温度对达乌尔黄鼠冬眠模式的影响
根据达乌尔黄鼠冬眠季节的长短和冬眠阵的长度,可以把达乌尔黄鼠的冬眠模式分为深冬眠型、少冬眠型和不冬眠型[5]。冬眠阵的长短在一定程度上取决于环境温度,环境温度降低会延长动物冬眠阵的长度[14],5℃环境温度下达乌尔黄鼠的冬眠阵长度显著长于15和25℃,而拥有最长最深的冬眠阵正是最理想的冬眠模式[15],蝙蝠(VespertilioLinnaeus)和睡鼠(G.glis)中都曾观察到超过30d的冬眠阵[16]。而通过观测达乌尔黄鼠不同环境温度下冬眠季节长度发现,随着环境温度的升高导致动物的冬眠季节变短,高的环境温度严重影响动物冬眠季节长度。在旱獭(Marnotamarmota)的研究中发现由于气候变暖,阿拉斯加旱獭冬眠时长较23年前减少38d[17]。动物在冬眠过程中,体温略高于环境温度几度[18],不同环境温度会影响动物体温,进而影响冬眠模式。不同环境温度下动物表达出的异温时长具有显著差异,这与不同环境温度下,动物的代谢率不同相适应,体温随着代谢率的降低而降低[16]。很多物种往往都有一个最适冬眠温度[16],在这个温度范围之内,动物可表现最长冬眠阵,最低代谢率和最适体温。旱獭(Marnotamarmota)冬眠最适冬眠体温一般在5~10℃[19],而啮齿动物一般在1~6℃[18]。通过3个环境温度下达乌尔黄鼠越冬的对比,5℃环境温度下有最适达乌尔黄鼠的冬眠模式。但在相同的温度下动物并不一定表现出相同的冬眠模式。在15℃环境温度下,有2只动物表现出少冬眠型,在25℃环境温度下有1只动物表现出不冬眠型。某些动物在15,25℃环境温度下只表现少冬眠或者不冬眠,这种现象可能是由动物自身非常规冬眠阵而并非环境温度的原因,本实验室在此前对连续3个冬眠季节的研究中都发现在低温中不冬眠或极少冬眠的个体[20]。还有研究在实验室条件恒定不变的情况下,27只花鼠(Eutamiassibiricus)中有26%一生都没有表达冬眠[21]。
3.2 环境温度对达乌尔黄鼠能量消耗的影响
达乌尔黄鼠属于贮脂类冬眠动物,贮脂类冬眠动物在常规环境温度下冬眠过程中停止摄食,利用体内贮存的脂肪分解供能[22]。本实验5℃环境温度下,动物在冬眠过程中取食量38.00g。通过分析实验数据,发现其中81.2%摄食发生在出眠前一个月的时间内。出眠前期,动物冬眠阵时间缩短,此时有较多阵间觉醒,阵间觉醒和阵间产热时间虽短,但消耗能量较高。旱獭(Marnotamarmota)在阵间觉醒期间的耗能占冬眠季节的72%,其中产热阶段占57%[23]。随着环境温度的升高,动物的冬眠阵变短,拥有更多的阵间觉醒[14],高温环境下冬眠的动物在阵间觉醒期间消耗了更多的能量。育肥期间动物在体内贮存一定量的脂肪,这些脂肪能够维持动物在低温下越冬的能量需求,当动物因为阵间觉醒增多而导致能量消耗增高时,动物就需要从外界获取可利用的食物,来补充能量缺失。近百年来,世界正以一种异乎寻常的速度变暖,气候变化对生物的影响甚为直接,高的环境温度导致动物具有更短的冬眠阵,更多的阵间觉醒,更多的能量消耗,这些能量均来自于体脂消耗的供给和食物摄入的补充。
冬眠是达乌尔黄鼠生活史中的一个环节,动物不仅要衡量能量与冬眠之间的关系,还要衡量能量与出眠后繁殖的问题。有研究表明,蟾蜍在暖冬下死亡率会增加,并且产卵数会减少[24]。随着全球气候的变暖,冬眠季节变短,动物在可利用食物资源还不够丰富的时候出眠,出眠后直接面临能量供给不足,自身体脂消耗过多,体内剩余的能量是否能够支撑动物在繁殖期间的需求,还有待于进一步的研究。
4 结 语
在冬眠期间,达乌尔黄鼠的冬眠季节长度,冬眠阵长度,冬眠期间异温累积时长均随着环境温度的升高而变短,因此从冬眠模式上看,高的环境温度不利于动物的冬眠。
在冬眠期间,达乌尔黄鼠的摄食量,体脂消耗,和冬眠期间总能量消耗均随着环境温度的升高而增大,因此从能量学角度来看,高的环境温度不利于动物的冬眠。
[1]WILZ M,HELDMAIER G.Comparison of hibernation,estivation and daily torpor in the edible dormouse,Glis glis[J].J Comp Chem Physiol B,2000,170(7):511-521.
[2]蔡益鹏,黄钦恒,赵海青,等.脑室注射6-羟多巴胺对黄鼠冬眠入眠的影响[J].生理学报,1992,44(2):175-180.
[3]FRENCH A R.Allometries of the durations of torpid and euthermic intervals during mammalian hibernation:a test of the theory of metabolic control of the timing of changes in body temperature[J].J Comp Chem Physiol B,1985,156(1):13-19.
[4]HUMPHRIES M M,THOMAS D W,KRAMER D L.The role of energy availability in mammalian hibernation:a cost-benefit approach[J].Physiol Biochem Zool,2003,76(2):165-179.
[5]杨明,王德华.哺乳动物的蛰眠:类型,物种分布与模式[J].兽类学报,2011,31(2):195-204.
[6]HELDMAIER G,ORTMANN S,ELVERT R.Natural hypometabolism during hibernation and daily torpor in mammals[J].Respiratory physiology &neurobiology,2004,141(3):317-329.
[7]杨明,邢昕,管淑君,等.达乌尔黄鼠冬眠期间体温的变化和冬眠模式[J].兽类学报,2011,31(4):387-395.
[8]HUMPHRIES M M,THOMAS D W,Hall C L,et al.The energetics of autumn mast hoarding in eastern chipmunks[J].Oecologia,2002,133(1):30-37.
[9]MICHENER G R.Sexual differences in over-winter torpor patterns of Richardson's ground squirrels in natural hibernacula[J].Oecologia,1992,89(3):397-406.
[10]MASAKI M,KOSHIMOTO C,TSUCHIYA K,et al.Body temperature profiles of the Korean field mouse Apodemus peninsulae during winter aggregation[J].Mammal Study,2005,30(1):33-40.
[11]刁颖,杨明,迟庆生.用改进的代谢笼方法测定黑线姬鼠的能量收支[J].沈阳师范大学学报:自然科学版,2007,25(2):241-244.
[12]SPEAKMAN J R,STUBBS R J,MERCER J G.Does body mass play a role in the regulation of food intake?[J].Proc Nutrition Soc,2002,61(4):473-487.
[13]赵志军.繁殖经历对黑线仓鼠哺乳期能量收支的影响[J].兽类学报,2012,32(1):33-41.
[14]GEISER F,KENAGY G J,WINGFIELD J C.Dietary cholesterol enhances torpor in a rodent hibernator[J].J Comp Chem Physiol B,1997,167(6):416-422.
[15]HARLOW H J,FRANK C L.The role of dietary fatty acids in the evolution of spontaneous and facultative hibernation patterns in prairie dogs[J].J Comp Chem Physiol B,2001,171(1):77-84.
[16]HELDMAIER G,ORTMANN S,ELVERT R.Natural hypometabolism during hibernation and daily torpor in mammals[J].Respiratory physiology &neurobiology,2004,141(3):317-329.
[17]DAVID A ,PETERW ,WILLAM R F.Effects of climate change on A antarctic sea ice and penguins[C]∥GREEN E R,HARLE M,SPALDING M,et al.Imp acts of C lim ate Chang e on W ild lif e.RSPU,U K,2001:26-27.
[18]BARNES B M.Freeze avoidance in a mammal:body temperatures below 0degree C in an Arctic hibernator[J].Science,1989,244(4912):1593-1595.
[19]RUF T,ARNOLD W.Mechanisms of social thermoregulation in hibernating alpine marmots(Marmota marmota)[M]∥Life in the Cold.Springer Berlin Heidelberg,2000:81-94.
[20]王有,袁丽丽,彭霞,等.达乌尔黄鼠冬眠模式年龄性别差异的实验研究[J].沈阳师范大学学报:自然科学版,2009,27(3):351-355.
[21]KONDO N,SEKIJIMA T,KONDO J,et al.Circannual control of hibernation by HP complex in the brain[J].Cell,2006,125(1):161-172.
[22]MUNRO D,THOMAS D W.The role of polyunsaturated fatty acids in the expression of torpor by mammals:a review[J].Zoology,2004,107(1):29-48.
[23]HELDMAIER G,ORTMANN S,KRTNER G.Energy requirement of hibernating alpine marmots[M]∥CAREY C,FLORANT G L,WUNDER B A,et al.Life in the Cold-Ecological,Physiological,and Molecular Mechanisms.Boulder:Westview Press,1993:175-183.
[24]READING C J.Linking global warming to amphibian declines through its effects on female body condition and survivorship[J].Oecologia,2007,151(1):125-131.
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
体育科技文献通报(2022年3期)2022-05-23
现代畜牧科技(2021年6期)2021-07-16
作文中学版(2020年1期)2020-11-25
中外文摘(2019年20期)2019-10-24
计算机与网络(2018年4期)2018-09-10
世界博览(2016年6期)2016-04-26
微型计算机(2016年4期)2016-03-29
现代妇女(2015年12期)2015-12-08
电影(2015年12期)2015-01-05
