
杉木苦竹混交林生长效应与凋落物分解研究
2014-09-16 03:22连华萍
福建林业科技 2014年3期
连华萍
(三明市竹业开发站,福建 三明 365000)
杉木苦竹混交林生长效应与凋落物分解研究
连华萍
(三明市竹业开发站,福建 三明 365000)
分析10年生杉木林经间伐后留养苦竹形成的混交复层林的生长效应及凋落物数量及分解状况。结果表明:SJK处理(杉木林进行间伐,保留杉木密度900株·hm-2,留养苦竹,满园后,保留苦竹6000株·hm-2)能够形成层次明显的复层林分,既有利于杉木培育大径材,又可以收获苦竹竹产品,具有较高的生长效应。杉木苦竹混交复层林,不仅苦竹凋落物量大,分解快,而且形成的林分结构,凋落物混合状态,有利于杉木凋落物的分解。SJK处理中凋落物周转期:苦竹2.27 a,杉木3.86 a,苦竹凋落物周转期比杉木短580 d;杉木纯林(杉木林不间伐,现存密度2075~2110株·hm-2)凋落物周转期为4.94 a,比SJK处理中杉木凋落物周转期长394 d。
杉木;苦竹;混交复层林;生长效应;凋落物;分解
杉木(Cunninghamialanceolata)是优良珍贵用材树种,速生丰产,材性好,用途广,在我国南方林区广为栽培。但杉木固有生物学特性所表现出的自肥能力较差的缺陷,导致地力衰退,引起了生产力的下降[1-4],在一定程度上制约着杉木的进一步发展。杉木地力衰退原因有诸多方面。与杉木纯林,尤其是较大密度经营方式有密切相关。较大密度杉木纯林,树种单一,结构简单,林下光照不足,温度较低,造成土壤动物和微生物种类和数量的减少,凋落物分解速率低,营养物质无法良性循环[5]。杉木纯林地力衰退的机理已有较多的研究,且研究结果表明:林分养分归还量肥土作用大小主要决定于林分的枯枝落叶量和分解率[6]。有关杉木及其混交林凋落物已有一些研究[7-12],但有关杉木苦竹混交林凋落物的分解作用尚未见报道。
苦竹(Pleioblastusamarus)是南方林区常见的竹类植物,资源多、分布广。长期以来多作为杂灌小竹处理或利用而不开发,处于野生、半野生状态。20世纪80年代有关苦竹开发技术研究得到重视,总结经营技术措施,并揭示了苦竹的生物学、生态学特性,为苦竹培育发展提供了生产实践经验,苦竹已成为福建省优良的笋竹两用经济竹种[13-14]。为此,在福建省沙县开展杉木苦竹混交林构建 与凋落物分解试验研究。试图通过林分结构调整,改善林分生态环境,加快凋落物的分解,促进林分营养物质的循环,达到改良土壤肥力的作用,实现林分的可持续经营。同时,通过与杉木混交,开拓苦竹的发展空间,不断提高经营效益。
1 试验地概况
试验地位于福建省沙县(26°6′—26°46′N、117°32′—118°6′E)西北部富口镇柳坑村12林班35小班,区域性气候系大陆性兼海洋性季风气候区。年平均气温19.2 ℃左右,年均降水量1780 mm,空气相对湿度82%,霜期79~84 d,霜日18~24 d。试验林位于海拔380~550 m的高丘林地,为1997年春营造的杉木人工林,造林密度3000株·hm-2,林地属较肥沃立地类型。2002年进行1次间伐,保存密度2025~2130株·hm-2。
2 试验方法
2.1 试验设计
2005年选择同一坡面,立地条件和经营生长状况相似的10年生杉木林进行试验。试验设4个处理,SCD:杉木林不再间伐,保留杉木纯林状态,现存密度2075~2110株·hm-2;SCX:杉木林进行间伐,保留杉木密度900株·hm-2,不留养苦竹;SJK:杉木林进行间伐,保留杉木密度900株·hm-2,留养苦竹,满园后,保留苦竹6000株·hm-2;SJD:杉木林进行间伐,保留杉木密度1200株·hm-2,留养苦竹,满园后,保留苦竹6000株·hm-2。试验采用随机区组试验设计,3次重复,每小区面积20 m×30 m。在每小区中按“X”形建立5个1 m×1 m的凋落物收集框,在相邻处建立相应5个1 m×1 m的小样方。在SCX、SJK和SJD处理中杉木苦竹混交林构建技术主要有:①非均匀式配置留笋技术。通过定向留笋疏笋,在杉木林冠下多疏笋、少留竹,在林间隙地多留笋少疏笋。②团状抚育。在杉木林冠下劈草清杂,林间隙地浅锄除草,适当垦复。③切鞭诱导。沿保留杉木林冠外缘进行开沟(深20~40 cm)切鞭诱导竹鞭向水平方向延伸或回窜。
2.2 测定方法
2.2.1 林分生长状况测定 从2005年开始每年测定主要生长因子。每木测定杉木胸径,每隔3株测高等生长因子。每年进行苦竹产笋量、立竹密度、高径生长等调查。测定产笋量(现场实测鲜重和产材量(每年实测平均胸径10株的重量,计算平均值,以此推算收获产材量和现存竹材量)。通过公式计算杉木立木材积。V杉=0.00005806D1.955335H0.894033。
2.2.2 凋落物数量测定 2010—2012年连续3年逐月收集网内凋落物,装入塑料袋内全部带回室内。分别杉木、苦竹,将枝、叶、花果、其他分别称重。在80 ℃下烘干至恒重,然后称重,据此含水率把凋落物各组分核算为干重。
2.2.3 凋落物及其分解速率测定 2010年开始采用样方收获法分别收集小样方内凋落物,对凋落物层分别L、F、H层称重后带回室内,取部分样品测定生物量。留取一定量的落叶凋落物,等量装入分解网袋,网袋规格为50 cm×50 cm,网眼2 mm,为便于计算,杉木以小网袋(规格20 cm×20 cm,网眼1 mm)隔开,套在大网袋中,从2010年6月1日起自然均匀地布设在林中,布设时使网袋落叶尽可能接近自然状态,并分树种测定各种分解样品初始每袋干重。每月分别随机抽取各类分解样袋5个,经80 ℃烘干至恒重后,根据含水率把凋落物换算为干重,计算落叶凋落物残留量和失重量。利用Olson(1963)[15]提出的凋落物分解公式,对分解过程中残留量Wt和初始量Wo用方程Wt=Woe-kt计算出凋落物分解系数,再计算凋落物的周转期。计算分解速率。
选择SJK处理测定年凋落物动态以SCD、SCX为对照,测定SJK等3种处理的凋落叶的分解速率;以SCD为对照,测定SJK等2种处理的凋落叶的周转期。
3 结果与分析
3.1 林分生长状况分析
森林是各种生物在复杂环境下生存的复合体,各生物为了生存对阳光、空气、水和各种养分而互相竞争[16]。同一物种个体间的竞争和不同物种个体间的竞争未必造成对一切竞争的小生境有相同的或相似的要求。杉木纯林间伐后留养苦竹,能否形成相对稳定,具有现实生产力的林分,主要表现在林分生长方面。从表1、表2可知:不同处理7 a后林分生长状况存在明显差异。SJK处理与SCD、SCX、SJD处理相比:杉木平均胸径分别增加20.7%、5.2%、7.2%,平均树高分别增加7.5%、2.1%、1.4%;SJK处理苦竹立竹数6015株·hm-2、平均胸径4.2 cm、平均高6.1 m,2010—2012年平均产笋量857.5 kg·hm-2、平均产材量5155.40 kg·hm-2。杉木冠层位于主林层,比苦竹分布层高2~4 m左右,形成层次明显的复层林分。既有利于杉木培育大径材,又可以收获苦竹竹产品。SJD处理与SJK处理相比,杉木保留株数增加33.3%。平均胸径降低6.7%,平均树高降低1.4%;苦竹保留株数基本相同,平均胸径降低11.9%,平均高降低9.8%。表明SJK处理,上层杉木保留900株·hm-2的低密度状态有利于杉木的生长和林中苦竹的生长发育。
SCX处理与SJK处理相比,杉木保留株数相等,林中没有留养苦竹,杉木平均胸径降低4.9%,平均树高降低2.1%。表明SJK处理,留养苦竹不仅没有影响上层杉木的生长,而且促进了杉木生长,说明杉木间伐,尤其是在较大强度间伐后,林地中腾出的生态位需要及时填补,减少其他恶性杂草侵入,防止水土流失。
SCD处理,保留杉木纯林状态,现存密度2075~2110株·hm-2,郁闭度达到0.87~0.95,与SJK处理相比,平均胸径降低17.2%,平均树高降低6.9%;与SJD处理相比,平均胸径降低11.2%,平均树高降低5.6%;与SCX处理相比,平均胸径降低12.9%,平均树高降低5.0%。SCD处理在4种处理中杉木胸径、树高均最小,难以培育杉木大径材。崔国发[17]认为;杉木林下光照不足、温度较低,形成了相对封闭的小气候,造成土壤动物和微生物种类和数量的减少,从而影响了凋落物的分解速率,导致土壤肥力难以恢复。同时,杉木较大密度所形成的单层结构林分,林下植被稀少,杉木个体间竞争加剧,影响高径生长。

表1 不同处理杉木间伐前后林木生长状况

表2 不同处理苦竹生长状况及产量
*:表中数据为2010—2012年平均值。
3.2 凋落物数量及组成
凋落物是森林土壤有机质的主要来源,它是森林生态系统中营养元素生物循环过程的重要环节,对地力维持和生态系统养分平衡起重大作用。从表3可知:2012年,SCD凋落量为3321.61 kg·hm-2;而SCX凋落量为1768.46 kg·hm-2,只有SCD凋落量的53.24%。显然这是由于保留株数较少的缘故。SJK凋落量为4042.02 kg·hm-2,其中杉木凋落量1659.87 kg·hm-2,占41.07%;苦竹凋落量2240.7 kg·hm-2,占55.44%;其他141.45 kg·hm-2,占3.5%;苦竹凋落量占较大比例。SJK凋落量与SCD、SCX相比,分别增加21.69%、128.56%,可见苦竹凋落量大于杉木凋落量。SJD凋落量为4055.48 kg·hm-2,其中杉木凋落量1715.24 kg·hm-2,占42.29%;苦竹凋落量2201.37 kg·hm-2,占54.28%;与SJK相比,杉木凋落量增加3.34%,苦竹凋落量降低1.76%。与SJD杉木株数增加,SJK苦竹个体增长有关。表明混交比例不同,不仅影响生长状况,而且凋落量也不同。

表3 不同处理凋落物数量及组成(2012年) kg·hm-2
不同处理凋落物组成存在差异。从表3可知:杉木纯林与杉木苦竹混交林凋落物组成存在明显不同。杉木纯林(SCD)枝叶占大多数,在总凋落物数量中杉木叶占75.21%,杉木枝占19.92%,杉木枝叶凋落物数量占总凋落物数量的95.13%。杉木苦竹混交林(SJK)凋落物数量中杉木叶占31.67%,杉木枝占8.32%,杉木枝叶凋落物数量占总凋落物数量的39.99%,杉木枝叶凋落物数量占总凋落物数量的比例明显下降;苦竹叶占34.48%,枝占15.74%,笋(秆)箨占5.21%,枝叶(含箨)占总凋落物数量的55.43%,这与苦竹生物学特性密切相关,苦竹与其它竹类植物一样,开花周期长,每年都有大量新竹形成,周期性换叶,所以枝叶(含箨)占绝大多数;其它植物凋落物数量占3.50%,其它植物凋落物数量所占比例与SCD相比有所下降,但其它凋落物数量比SCD净增11.24 kg·hm-2,增加8.6%。其它凋落物数量的提高,从一个侧面表明杉木苦竹混交林林分结构的改变,有利于生物多样性的形成,对提高林分的稳定性和抗逆性有积极作用。
3.3 年凋落物量动态变化
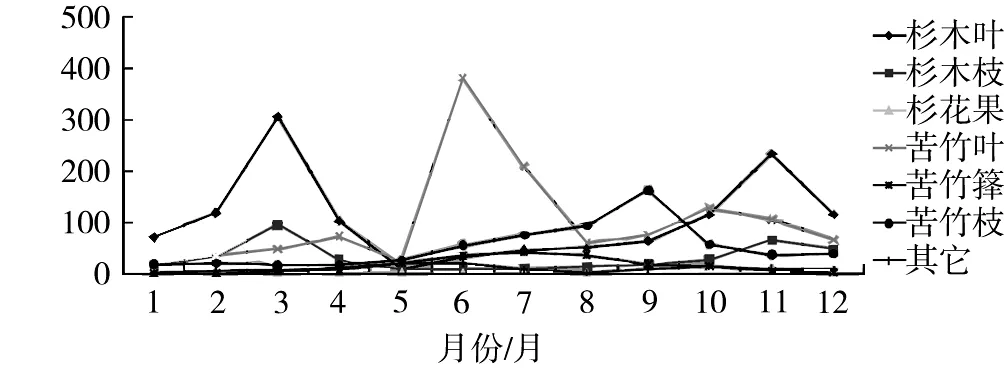
图1 杉木苦竹混交林凋落物月动态
不同树种不同月份凋落量存在差异(图1)。SJK处理平均年凋落物量3977.7 kg·hm-2,其中苦竹2107.7 kg·hm-2,占53.0%;杉木1731.6 kg·hm-2,占43.5%;其余为其他生物凋落物。从图1可以看出,凋落物组分、数量在各月的分布是不均匀的,无论是杉木还是苦竹,凋落量的月动态波动较大。2012年杉木凋落物呈明显双峰型,第1个高峰期在3月,凋落量占全年的23.6%;第2个高峰期在11月,凋落量占全年的17.8%。6—10月凋落量均较小,凋落量最小月份出现在5月,月最大凋落量是最小凋落量的15.5倍。这与温远光等[7]、马祥庆等[8]研究较吻合,但第2个高峰值出现时间比马祥庆研究结论提早30 d。这可能与经营密度较小,且下层保留大量苦竹有关。曹永康[9]研究6~9年生杉木幼林结果表明:杉木凋落量峰型,不同年份呈不同动态变化,有3峰型、双峰型和单峰型,与每年气候尤其是降雨量有关,表现出非生理性落叶因素。笔者认为:杉木凋落量双峰型是主要峰型,且出现在季节变化期,系生理性落叶。杉木落叶与落枝基本同步,针叶经常连着小枝一起掉落。这种凋落方式,使杉木针叶难以与土壤密切接触,杉木枝叶含有防腐物质,较难分解[18],不利于针叶的进一步分解。
苦竹凋落物的月动态变化与杉木不同。苦竹凋落量峰型呈大小双峰型,主峰值出现在6月,凋落量占全年的22.5%,凋落物主要为竹叶。这与苦竹生物学特性相关。苦竹在4月中下旬出笋,5月底新竹高生长基本停止,开始展叶,6月出现大量换叶,但竹枝尚未脱落,之后7—8月枝逐渐凋落,在9月出现小峰值,凋落量占全年的12.5%,这一阶段落枝是主要凋落物。苦竹与杉木凋落方式明显不同在于竹叶与竹枝是分开凋落,且变态叶(箨)叶面积大,当年生,较幼嫩,易被分解。
3.4 分解速率
有机物经分解者分解成无机物,回归无机自然界是物质循环的另一个重要环节。杉木苦竹混交林(SJK)凋落物现存量18.66 t·hm-2(表4),具明显的F层(半分解)、H层(腐殖质)、L层(枯枝落叶)。杉木纯林凋落物SCD现存量9.21 t·hm-2,SCX现存量 5.67 t·hm-2,SJK分别是SCD、SCX的2倍、3.3倍。SCD、SCX、SJK半分解和腐殖质与枯枝落叶的比值分别为2.30、0.27、0.41,可见混交林中枯枝落叶分解速率比杉木纯林快。一方面说明苦竹凋落物分解快于杉木凋落物,另一方面意味着杉木苦竹凋落物混合后,可能加快了杉木凋落物的分解。苦竹凋落量大、分解快,应用苦竹与杉木混交可防止林地地力衰退,维持林业的可持续经营。
凋落物的分解速率应用衰减指数模型,Wt=Woe-kt计算。

表4 不同处理林分凋落物现存量及分配
*:比值为F、H之和与L的比值。

表5 不同处理凋落叶年分解速率及周转期
根据3 a的测定结果,不同林分,不同树种分解速率不同。从表5可知,不同处理、不同树种年分解速率存在差异,混交林中,苦竹落叶年分解速率比混交林中杉木落叶年分解速率提高1.70倍,比杉木纯林提高2.18倍。这与不同树种分解基质的类型、凋落的季节、凋落规律有关。而混交林中杉木与杉木纯林分解速率的差别,主要与环境因子有关。混交林中温度、湿度以及微生物、腐食小动物等分解者的数量等因素优于纯林[19-22]。试验结果表明,落叶95%分解时,SJK中杉木所需要时间为3.86 a(1409 d),苦竹所需时间为2.27 a(827 d),苦竹落叶凋落物的周转时间比杉木能够提早580 d。表明苦竹落叶的分解速率较快,有利于营养元素的生物循环。苦竹 叶片较大,每年有大量叶片凋落,易分解。而混交林形成荫蔽湿润的小环境也有利于枯枝落叶分解。而杉木纯林中杉木落叶凋落物周转时间比SJK中杉木所需要时间多394 d,表明杉木苦竹凋落混合物能够促进杉木的分解,而通过构建杉木苦竹混交复层林能够在一定程度上提高林分的自我培肥能力。有关杉木凋落叶周转期前人试验结果[23-26]认为杉木凋落叶周转期3.89~5.73 a不等,均大于本试验SJK处理中杉木周转期(3.86 a),从一个侧面说明杉木苦竹混交林有利于杉木凋落叶的分解。
4 小结
杉木苦竹混交复层林具有较高的生长效应。SJK处理(杉木林进行间伐,保留杉木密度900株·hm-2,留养苦竹,满园后,保留苦竹6000株·hm-2),7a后现存杉木900株·hm-2,平均胸径16.3 cm,平均树高14.4 m,与SCD、SCX、SJD处理相比,杉木平均胸径分别增加20.7%、5.2%、7.2%,平均树高分别增加7.5%、2.1%、1.4%;苦竹6015株·hm-2,平均胸径4.2 cm,平均高6.1 m,2010—2012年平均产笋量857.5 kg·hm-2,平均产材量5155.4 kg·hm-2。杉木冠层位于主林层,比苦竹分布层高2~4 m左右,形成层次明显的复层林分。既有利于杉木培育大径材,又可以收获苦竹竹产品。
杉木苦竹混交复层林增加了凋落物量,促进了凋落物分解。研究结果表明:SJK处理凋落量为4042.02 kg·hm-2,其中来自杉木凋落量1659.87 kg·hm-2,占41.09%;苦竹凋落量2240.70 kg·hm-2,占55.44%;其它141.45 kg·hm-2,占3.5%。凋落物量从大到小依序为:SJD处理>SJK处理> SCD处理>SCX处理.SJK处理中无论杉木、苦竹凋落物月动态波动较大。杉木凋落节律呈明显双峰型,第1次高峰出现在3月,第2次高峰出现在11月,苦竹凋落节律为大小峰型,主峰值出现在6月,次峰值在9月,枝叶分开凋落。凋落节律和落叶方式与杉木不同。SJK、SCX、SCD处理凋落物现存量中F、H、L比值分别为2.30、0.27、0.41。混交林中凋落物的分解速率比杉木纯林快。采用衰减指数模型测算,SJK处理中苦竹落叶年分解速率比SJK处理中杉木提高1.70倍,比杉木纯林(SCD)提高2.18倍。按估算SJK处理中苦竹凋落物周转期2.27 a,杉木3.86 a,杉木纯林4.94 a。SJK处理中苦竹周转期比杉木可提早580 d,杉木纯林中杉木落叶凋落物周转时间比SJK中杉木所需时间多198 d。
杉木苦竹混交复层林,不仅苦竹凋落物量大,分解快,而且形成的林分结构,凋落物混合状态,有利于促进杉木凋落物的分解,对维持地力,实现可持续发展有重要积极的作用。在通常状态下,营养元素的归还途径主要通过枝叶枯落,根系更新遗留,昆虫有机体的归还及降雨淋洗。由于条件限制,本研究仅对枝叶枯落归还定期进行测定分析。混交林和纯林由于林木组成,凋落物数量及性质等的差异,直接影响营养元素的归还,进而影响土壤养分贮量及有效性。有关营养物质循环有待于进一步研究。
[1]方奇.杉木连栽对土壤肥力及其杉木生长的影响[J].林业科学,1987,23(4):389-397.
[2]俞新妥.杉木连栽林地土壤生物特性及土壤肥力的研究[J].福建林学院学报,1989,9(3):263-271.
[3]杨承栋.杉木人工林地力衰退的原因机制及其防治措施[J].世界林业研究,1997,10(4):34-39.
[4]俞新妥.杉木栽培学[M].福州:福建科学技术出版社,1997.
[5]崔国发.人工林地力衰退机理及其防止对策[J].世界林业研究,1996(5):61-69.
[6]俞新妥.混交林营造原理及技术[M].北京:中国林业出版社,1989.
[7]温远光,韦胜章,秦武明.杉木人工林凋落物动态及其与气候因素的相关分析[J].生态学报,1990,10(4):367-372.
[8]马祥庆,刘爱琴,何智英,等.杉木幼林生态系统凋落物及其分解作用研究[J].植物生态学报,1997,21(6):564-570.
[9]曹永康.6—9年杉木幼林凋落物及其养分特征[J].亚热带资源与环境学报,2010,5(1):42-47 .
[10]吴承祯,洪伟.武夷山杉木林凋落物动态初探[J].热带亚热带植物学报,2002,10(3):201-206.
[11]周东雄.杉木乳源木莲混交林凋落物研究[J].生态学杂志,2005,24(6):595-598.
[12]杨玉胜,郭剑芬,陈银秀,等.福建柏和杉木人工林凋落物分解及养分动态的比较[J].林业科学,2004,40(3):19-24.
[13]连标勇,周东雄.福建沙县苦竹经营技术调查研究[C]//中国林学会经济林分会.中国经济林现状与对策.北京:中国林业出版,1996:279-283.
[14]周东雄.马尾松苦竹复合林分生产力研究[J].林业科技开发,2002,16(2):27-29.
[15]Olosn.JS.Energystoragenadthebalnaceofproducersnaddeeomposersinecologicalsystem[J].Ecoloyg,1963(44):322-331.
[16]S.H斯波尔,R.V巴恩斯.森林生态学[M].赵克绳,周祉,译.北京:中国林业出版社,1982.
[17]崔国发.人工林地力衰退机理及其防止对策[J].世界林业研究,1996(5):61-69.
[18]盛炜彤,薛秀康.福建柏、杉木及其混交林生长与生态效应研究[J].林业科学,1992,28(5)397-404.
[19]胡肄慧,陈灵芝,孔繁志.两种中国特有树种的枯枝分解速率[J].植物生态学与地植物学报,1986,10(1):35-43.
[20]杨玉盛.南平溪后杉木林取代杂木林后土壤肥力变化的研究[J].植物生态学学报,1994,18(3):236-242.
[21]Mesquita RDCG,Workman SW,Neely CL.slow litter decomposition in a Cecropia-dominated secondary forest of central amazonia[J].soil biochem.,1998,30(2):167-175.
[22]Staaf H.Influrnce of chemical composition,addition of raspberry leave,and nitrogen supply on decomposition rate and dynamics of nitrogen and phosphorus in beech leaf litter[J].oikos,1980,35(1);55-62.
[23]樊后保,刘文飞,谢友森,等.杉木人工林凋落物分解动态[J].南昌工程学院学报,2007,26(4):1-4.
[24]马祥庆,刘爱琴,何智英.杉木幼林生态系统凋落物及其分解作用研究[J].植物生态学报,1997,2(6):564-570.
[25]何宗明,陈光水,刘剑斌.杉木林凋落物产量、分解率与储量的关系[J].应用与环境生物学报,2003,9(4):352-356.
[26]薛立,邝立钢.杉木凋落物分解速率的研究[J].四川林业科技,1990,11(1):1-4.
Tree Growth and Litter Decomposition in a Mixed Plantation of Chinese Fir andPleioblastusamarus
LIAN Hua-ping
(SanmingBambooDevelopmentStation,Sanming365000,Fujian,China)
Tree growth,litter production and decomposition of a multi-storey mixed stand of Chinese fir (Cunninghamialanceolata) andPleioblastusamarus,developed from thinning a 10-year-old pure Chinese fir stand and retaining understorey speciesP.amarus,were studied in this article.Treatment SJK (thinning a pure Chinese fir stand to density 900 stems·hm-2and retaining understorey speciesP.amarusto density 6000 stems·hm-2) could develop obvious multi-storey crown,it was in favor of cultivating Chinese fir big-diameter timber,provided extra bamboo products and improved tree growth.Multi-storey mixed stand of Chinese fir andP.amarushad good stand structure,higher litter production and decomposition rate ofP.amarus,and litter mixture had a positive effect on decomposition rate of Chinese fir litter.Turnover time ofP.amarusand Chinese fir leaf litter in treatment SJK were 2.27 a and 3.86 a,respectively,and the former was 580 d shorter than the latter.Turnover time of Chinese fir leaf litter in pure plantation (unthinned,current density 2075~2110 stems·hm-2) was 4.94 a,394 d longer than that in the mixed stand with treatment SJK.
Chinese fir;Pleioblastusamarus; multi-storey mixed stand;growth;litter;decomposition
10.13428/j.cnki.fjlk.2014.03.002
2013-11-12;
2013-12-27
福建省林业厅林业科研基金资助项目(闽林科[2012]2号)
连华萍(1964—),女,福建福安人,三明市竹业开发站高级工程师,从事森林资源培育研究。E-mail:smlhp1964@163.com。
S725.2
A
1002-7351(2014)03-0006-06
猜你喜欢
东北财经大学学报(2022年1期)2022-02-23
广西植物(2021年9期)2021-10-16
商场现代化(2021年10期)2021-07-29
滇池(2019年11期)2019-11-22
湖北林业科技(2019年1期)2019-06-20
中国乡镇企业会计(2019年3期)2019-04-12
森林工程(2018年4期)2018-08-04
中国乡镇企业会计(2018年1期)2018-07-12
方圆(2018年23期)2018-01-07
广西林业科学(2016年4期)2016-03-16
