
一种混合蜂群算法的自适应细菌觅食优化算法
2014-09-29 10:31杜鹏桢唐振民
计算机工程 2014年7期
杜鹏桢,唐振民,孙 研
(南京理工大学计算机科学与工程学院,南京 210094)
1 概述
细菌觅食优化算法(Bacterial Foraging Optimization Algorithm,BFOA)[1]是一种模拟细菌觅食行为的群优化算法,现已成功运用于经济调度[2]、图像处理[3]等领域。BFOA在适当的趋化步长下,通过在解空间进行连续的翻转和泳动可以达到较高的精度,是一种精细型搜索算法。但由于其采用直接复制的繁殖方式,降低了种群的多样性,容易陷入局部最优[4]。为克服基本BFOA存在的缺点,各学者提出了很多改进方法,例如:文献[5]将量子行为引入细菌的繁殖操作,增强了BFOA的种群多样性,提高了算法的全局搜索能力;文献[6-7]采用自适应的趋化步长,提高了算法的整体性能;文献[8-9]分别将BFOA与其他算法相结合,改进了繁殖方式,大幅提高了算法的求解质量。
将多种算法进行混合,发挥每个算法的优点,是当前智能优化领域的研究热点之一。文献[10]研究表明,人工蜂群(Artificial Bee Colony,ABC)算法的寻优能力与粒子群优化(Particle Swarm Optimization,PSO)算法相当,强于差分进化,且在同等条件下,ABC的收敛速度可达PSO的2倍[11],是一种比较有竞争力的新型智能优化算法。为克服BFOA的缺点,本文将ABC的部分机理引入到BFOA中,提出一种混合ABC算法的自适应BFOA算法(Bee Bacterial Foraging Optimization Algorithm,BBFO)。相比基本BFOA,BBFO有以下5点改进:(1)提出雇佣蜂式趋化,对解空间进行全局寻优;(2)改进BFOA原有趋化方式并自适应调节趋化步长,完成对解空间的局部调优;(3)依据对种群的多样性评价,完成2种趋化方式的自动切换;(4)提出基于t分布扰动的复制方式,提高种群多样性;(5)提出基于对立学习的侦察蜂式迁移,避免算法早熟。
2 细菌觅食优化算法
为方便描述,本文定义S个细菌组成的种群为:

其中,θi(j,k,l)表示第j次趋化,第k次复制,第l次驱散后的第i个个体。
BFOA把细菌觅食行为分解为趋化、聚群、复制和驱散4种算子。
(1)趋化算子模拟细菌向营养丰富区域移动的过程,包含翻转和泳动2种基本动作。个体i在n维解空间 ℝn按式(1)进行翻转,C(i)为趋化步长。如果翻转后细菌适应值得到改善,则沿当前方向φ(i)继续泳动预定步数Ns或泳动到适应值不再改善为止。

(2)聚群算子模拟个体间的相互影响,将个体间的吸引和排斥依式(3)计算为适应值Jcc,并与个体原适应值进行叠加,见式(4)。其中,dattract,wattract,h r epellant 和wrepellant为预先设定的吸引因子、吸引权重、排斥因子和排斥权重。

(3)复制算子模拟种群的繁殖。种群经过Nc次趋化后,依据适应值的累加和进行排序,较优的 Sr=S /2个细菌进行分裂繁殖,直接复制自身替换较差的Sr个细菌。
(4)驱散算子模拟种群的迁移。经过Nre次复制后,个体以概率Ped随机生成新个体,并替换原个体。
BFOA算法步骤如下:
步骤1初始化参数S,Nc,Ns,Nre,Ned,Ped,C(i),da,wa,hr和wr,在解空间 ℝn内随机初始化种群。
步骤2循环执行趋化算子Nc次,每次翻转操作后执行聚群算子。
步骤3执行复制算子,复制算子执行次数加1。若复制算子已执行Nre次,执行次数清0,转去执行步骤4,否则转去执行步骤2。
步骤4执行驱散算子,驱散算子执行次数加1。若驱散算子已执行Ned次,算法结束,否则转去执行步骤2。
3 BBFO算法
3.1 趋化
BBFO算法采用2种趋化方式:雇佣蜂式趋化(Employed Bees Style Chemotaxis,EC)和改进的自适应步长趋化(Improved Chemotaxis,IC)。
3.1.1 雇佣蜂式趋化
为提高BBFO的全局搜索能力,EC借鉴ABC雇佣蜂更新蜜源的方式[10],步长根据下式确定:

其中,Cec(i)表示个体i的趋化步长;rand(-1,1)表示–1与1之间的随机数;表示第k(k≠i)只细菌第j维的值,k和j均为随机生成。在EC中,无翻转操作,仅执行单维度泳动,个体i沿步长Cec(i)泳动,若适应值无改善,则退回原来位置。
3.1.2 改进的自适应步长趋化
基本BFOA对步长较为敏感,取值大则全局搜索能力强,但精度下降;取值小则局部调优能力强,但收敛速度下降,算法时间复杂度上升。在BBFO中,IC采用自适应步长,见式(6),泳动依然采用式(1)。

其中,Cic(i,j)表示第i个个体第j次趋化后的步长;δ∈(0,0.95)为步长缩减因子;Cinit(i)表示第i个个体的初始步长;domxh为解空间定义域的右边界;domxl为解空间定义域的左边界;D为自然数。
3.2 种群多样性的评价及趋化方式的切换
算法初始时,种群分散在一个较大空间,多样性较强,此时适宜进行全局搜索,加快算法收敛。随着迭代次数的增加,种群不可避免地聚集在相对狭小的空间,此时更适宜进行局部调优,以提高求解精度。鉴于此,本文利用式(8)[12]评价种群多样性,同时引入阈值 Pc,当 diversity(P) >Pc时,采用雇佣蜂式趋化进行全局搜索并提高收敛速度,否则,采用改进的自适应步长趋化,提高求解精度。当 diversity(P)较小时,种群多样性较低,此时算法可能已经陷入局部最优,具体跳出局部最优的方法将在下文进行阐述。

3.3 基于t分布扰动的复制方式
复制是BFOA的主要进化操作[13],为避免BFOA中直接复制所带来的多样性降低问题,BBFO采用基于t分布扰动的复制方式,具体见式(9)。

其中,θi为被替换个体;θk为被复制个体;t(υ)表示自由度为υ的t分布。
t分布又称学生分布,当υ趋向无穷大时,等同于标准高斯分布,当υ=1时,等同于柯西分布。本文将种群多样性作为t分布的自由度,见式(10),当种群多样性较低时,t分布近似于柯西分布,具有较大的扰动,有利于跳出局部最优,增强全局搜索能力;当种群多样性较高时,t分布近似于标准高斯分布,扰动较小,更利于提高求解精度。
3.4 基于对立学习的侦察蜂式迁移
在ABC算法中,若蜜源一定次数内无法被改善,则抛弃该蜜源,由相应的侦察蜂重新对解空间进行随机搜索。BBFO借鉴侦察蜂方式,设立参数trails,若某个解连续trails次趋化无法被更新,则重新生成该解。由于BBFO是精细型搜索算法,为了避免在原位置附近进行无用的搜索,同时提高跳出局部最优的能力,本文采用对立学习的方法生成新解,迁移方式如下:

其中,θi为迁移个体;θio为θi的近似对立数;domxh为解空间定义域的右边界;domxl为解空间定义域的左边界。
3.5 算法流程
BBFO中不需要参数Ns,整个流程采用3层循环的框架,对大小为S的种群,需要 Ned×Nre×Nc次评价,其时间复杂度为O(Ned×Nre×Nc),具体的流程如下伪码所示:


4 仿真分析
为了验证BBFO的性能,本文选取10个Benchmark函数进行测试,所有函数均为30维,理论最小值均为0,详见表1。其中,f1~f7为单峰函数,用于测试算法的求解精度和收敛速度,f8~f10为多峰函数,用于测试算法的搜索能力和跳出局部最优的能力。测试所引入的对比算法有:基本BFOA[1],ABC[10],BSA[7],QBFO[5]和EDABFO[6],相关参数参照相应文献。由于BBFO将ABC引入BFOA,因此本文将其与ABC和BFOA进行纵向比较,同时,从混合型算法的角度出发,本文将BBFO与BSA、EDABFO和QBFO进行横向比较。BBFO参数设置如下:种群大小S=100,切换阈值 Pc=0.08,缩减因子 δ=0.08,定义域划分数D=50,迁移次数 Ned=10,复制次数Nre=8,趋化次数Nc=40,总迭代次数为Ned·Nre·Nc= 320 0。仿真平台CPU 2.9 GHz,内存2 GB,VS2010编译套件。

表1 测试函数
4.1 寻优性能比较
针对表1函数,ABC、BFOA和BBFO各运算30次,每次进行3200次迭代,所得统计结果见表2。BFOA是一种精细型搜索算法,全局搜索能力较弱,在测试中效果不理想,相应的,其局部寻优能力强的优点并未得到体现。ABC具有较强的全局搜索能力,也获得了较为理想的效果,但与BBFO相比,还有较大的局部调优空间。最优值可以衡量算法的精度,平均值衡量算法的性能,标准差衡量算法的稳定性。BBFO在各方面均优于ABC和BFOA,兼具两者优点。最优值和平均值的比对表明,BBFO在具有较高的全局寻优能力的同时,保持了较高的局部调优能力。标准差的对比结果表明BBFO具有较强的稳定性。

表2 寻优性能纵向对比
为进一步验证BBFO的有效性,将其与BSA、EDABFO和QBFO进行了横向比对。其中,BBFO进行1000次迭代,由于BBFO的2种趋化依据多样性进行自动切换,因此切换阈值更改为Pc=0.15,结果见表3。可以看出,BBFO的平均值和标准差均优于对比算法,表明BBFO具有较高的全局搜索能力,同时具有较强的鲁棒性。

表3 寻优性能横向对比
4.2 收敛速度比较
收敛速度是衡量算法的重要指标之一,为对BBFO的收敛速度进行验证,本文选取Rosenbrock函数和Schwefel 1.2函数进行测试。Rosenbrock函数在最优值附近存在陡峭区域,易陷入局部最优,相关算法在测试中效果并不理想,Schwefel 1.2函数最优值存在于一个狭小区域,比较考验算法的局部调优性能。结合4.1节寻优性能的比对实验,可对算法进行较为全面的测试。由图1可见:(1)BFOA收敛速度曲线整体比较平缓;(2)迭代早期BBFO收敛速度与ABC相当;(3)在某一迭代区间,BBFO开始高速收敛。BFOA全局搜索能力较弱,在2个测试中均未搜索到全局最优区域,仅在局部最优区域进行调优,因而整体效果较差。BBFO存在2种趋化方式,迭代早期种群多样性较高,采用雇佣蜂式趋化,寻优性能与ABC相当。但从某个迭代区间开始,BBFO依据种群多样性自动切换为改进的自适应步长趋化,在图中表现为曲线的急速下降。从整体效果看,BBFO明显优于ABC和BFOA,充分证明了本文算法的有效性。
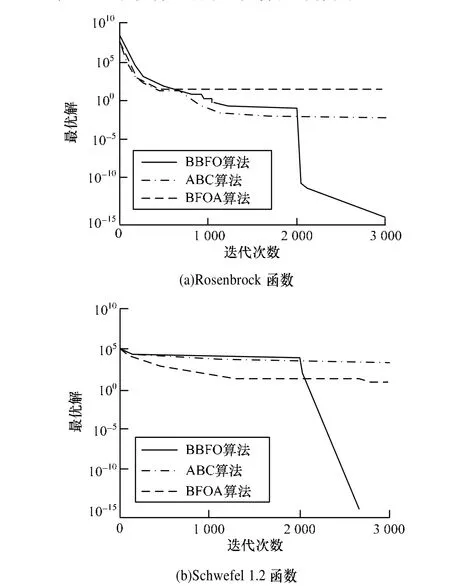
图1 收敛速度比较
4.3 多样性分析
在一般情况下,较高的种群多样性,表示算法有更大的可能跳出局部最优,可以有效避免早熟。但当已经搜索到全局最优区域时,过高的种群多样性反而表示算法收敛速度较低,局部调优能力不足。BBFO中2种趋化方式的切换和t分布扰动都基于种群多样性。为验证BBFO中种群多样性的重要性,对BFOA、ABC和BBFO进行仿真比对,结果见图2,从中可以看出,ABC一直保持较高的种群多样性,具有较强的全局搜索能力,但结合图1可见,在迭代后期,ABC的收敛速度明显下降,表明ABC局部调优能力不足。基本BFOA以固定概率进行随机迁移,很大程度上提高了种群的多样性,但其繁殖方式导致多样性迅速下降,所以多样性曲线以固定迭代代数波动。BBFO在迭代早期拥有较强的全局搜索能力,相应多样性保持在一个较高的水平上,在迭代后期,随着趋化方式的改变,算法侧重于局部调优,种群多样性迅速下降。特别的,图2(a)中BBFO的多样性曲线在迭代后期出现了剧烈的波动,原因是基于t分布扰动的复制方式在起作用,随着多样性的降低,t分布近似于柯西分布,可以产生较大的扰动。整体来看,BBFO依据多样性进行趋化方式自动切换的理念是合理可行的,效果非常明显。
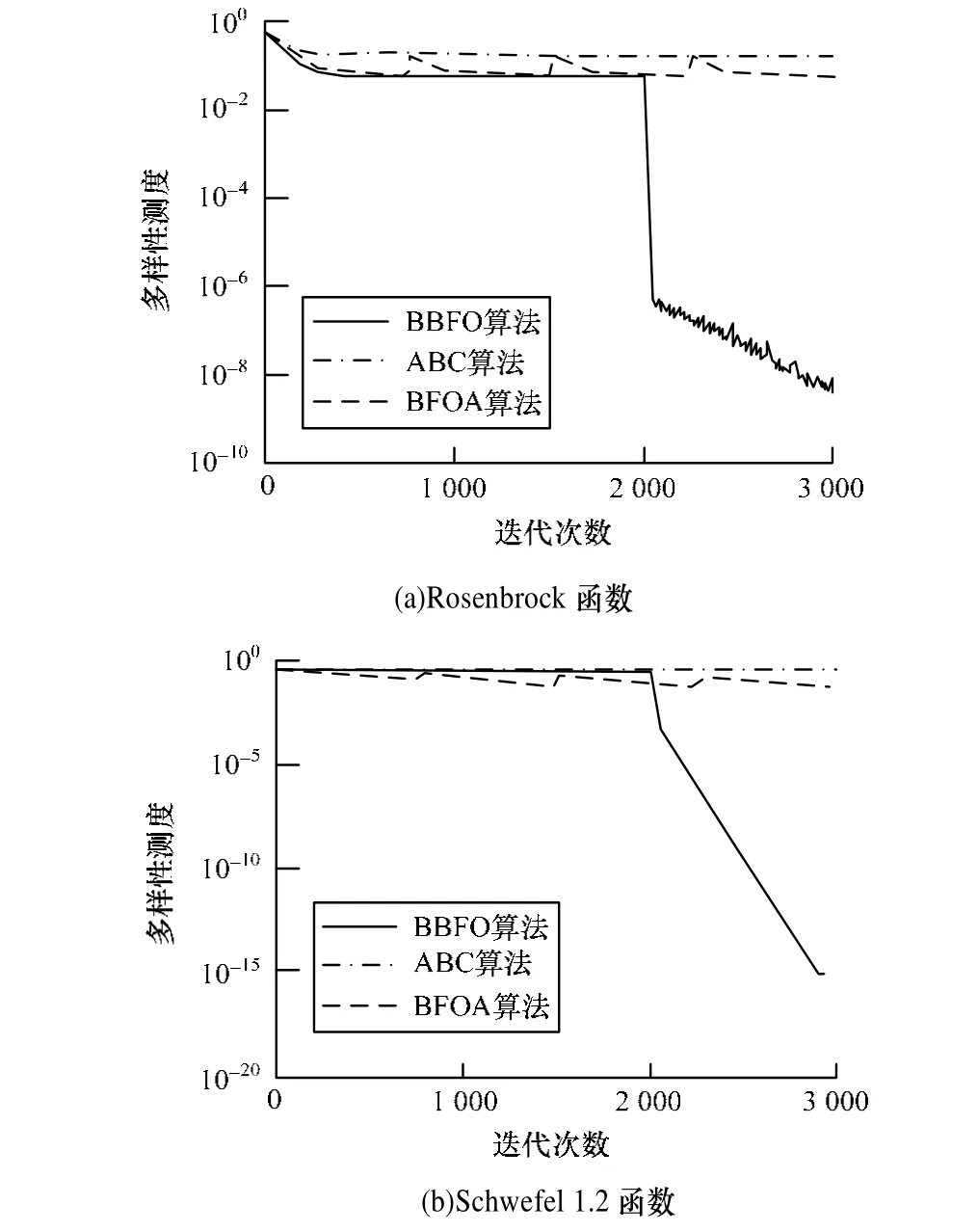
图2 种群多样性比较
5 结束语
在对细菌觅食优化算法进行研究的基础上,本文提出一种混合蜂群算法的自适应细菌觅食优化算法(BBFO)。通过设计新的雇佣蜂式趋化方式提高了全局搜索能力,同时改进BFOA原有趋化中的固定步长为自适应步长,充分发挥了BFOA局部调优能力强的优点。在此基础上,引入种群多样性评价,并依据评价结果完成2种趋化方式的自动切换。提出基于t分布扰动的复制方式,有效提高了算法的收敛速度和求解精度。此外,还将种群多样性作为t分布的自由度,使扰动强度可以自适应调节。为避免算法早熟,提出基于对立学习的侦察蜂式迁移,在跳出局部最优的同时提高了种群的多样性。实验结果表明,本文算法在寻优能力、求解精度和收敛速度方面均优于对比算法,并具有较强的稳定性。下一步工作是利用该算法解决更高维复杂的工程问题,并应用于相关收敛性理论的研究。
[1]Passino K M.Biomimicry of Bacterial Foraging for Distributed Optimization and Control[J].IEEE Control Systems,2002,22(3):52-67.
[2]Pandit N,Tripathi A,Tapaswi S,et al.An Improved Bacterial Foraging Algorithm for Combined Static/Dynamic Environmental Economic Dispatch[J].Applied Soft Computing,2012,36(12):3500-3513.
[3]Sathya P D,Kayalvizhi R.Modified Bacterial Foraging Algorithm Based Multilevel Thresholding for Image Segmentation[J].Engineering Applications of Artificial Intelligence,2011,24(4):595-615.
[4]Chatzis S P,Koukas S.Numerical Optimization Using Synergetic Swarms of Foraging Bacterial Populations[J].Expert Systems with Applications,2011,38(12):15332-15343.
[5]章国勇,伍永刚,谭宇翔.一种具有量子行为的细菌觅食优化算法[J].电子与信息学报,2013,35(3):614-621.
[6]王雪松,程玉虎,郝名林.基于细菌觅食行为的分布估计算法在预测控制中的应用[J].电子学报,2012,38(2):333-339.
[7]Tang W J,Wu Q H,Saunders J R.A Bacterial Swarming Algorithm for Global Optimization[C]//Proceedings of CEC’07.Singapore:IEEE Press,2007:1207-1212.
[8]刘小龙,赵奎领.基于免疫算法的细菌觅食优化算法[J].计算机应用,2012,32(3):634-637.
[9]刘小龙,李荣钧,杨 萍.基于高斯分布估计的细菌觅食优化算法[J].控制与决策,2011,26(8):1233-1238.
[10]Akay B,Karaboga D.A Modified Artificial Bee Colony Algorithm for Real-parameter Optimization[J].Information Sciences,2012,192(6):120-142.
[11]El-Abd M.Performance Assessment of Foraging Algorithms Vs.Evolutionary Algorithms[J].Information Sciences,2012,182(1):243-263.
[12]Sun J,Fang W,Palade V,et al.Quantum-behaved Particle Swarm Optimization with Gaussian Distributed Local Attractor Point[J].Applied Mathematics and Computation,2011,218(7):3763-3775.
[13]Abraham A,Biswas A,Dasgupta S,et al.Analysis of Reproduction Operator in Bacterial Foraging Optimization Algorithm[C]//Proceedings of CEC’08.Hong Kong,China:[s.n.],2008:1476-1483.
猜你喜欢
数学物理学报(2022年4期)2022-08-22
数学年刊A辑(中文版)(2022年1期)2022-08-20
数学物理学报(2022年2期)2022-04-26
成都信息工程大学学报(2021年5期)2021-12-30
数学物理学报(2021年5期)2021-11-19
数学年刊A辑(中文版)(2020年2期)2020-07-25
金桥(2018年4期)2018-09-26
哈尔滨师范大学自然科学学报(2015年2期)2015-04-19
中国卫生(2014年5期)2014-11-10
电测与仪表(2014年2期)2014-04-04
