
浙江省乐清湾围塘养殖泥蚶繁殖生物学研究
2014-10-08 07:33于铭茹YUMingru
价值工程 2014年14期
于铭茹 YU Ming-ru
(国家海洋标准计量中心,天津 300112)
(National Center of Ocean Standards and Metrology,Tianjin 300112,China)
0 引言
泥蚶(Tegillarca granosa)隶属软体动物门(Mollusca)、瓣鳃纲(Lamellibranchia)、翼形亚纲(Pteriomorphia)、蚶目(Arcoida)、蚶科(Arcidae)、蚶属(Tegillarca),俗称血蚶、宁蚶、花蚶、银蚶、蚶子等,属热带及温带生物,是一种栖息于沿海滩涂的广温性广盐性双壳类软体动物。泥蚶为滩涂中营埋栖生活的种类,喜栖于中、低潮区交界处,在生态上和经济上都具有重要作用。
浙江乐清湾(介于东经 120°2′-120°10′和北纬 27°58′-28°24′之间)位于浙江省瓯江口的北侧,是一个葫芦形的内湾,湾长40km,平均宽度10km,全湾有水面270km2,标准水面上的滩涂约200km2,是我国最大的泥蚶苗种生产和养殖基地,泥蚶苗种产量占全国市场份额的80%以上,有“中国泥蚶之乡”之称,也是我国较早研究人工育苗和中间培育的场所,每年为南方各省开展泥蚶养殖提供一定数量苗种。由于近年来海上活动的日益增加,海洋环境急剧恶化,海洋资源严重受损,泥蚶赖以生存的生态环境遭破坏,天然资源日趋衰竭,产量每况愈下。发展泥蚶增养殖业,需要得到高质量的精子和卵子以保证幼虫的健康,为了得到高质量的精子和卵子,有必要了解泥蚶自然群体的状况,故对该地区泥蚶繁殖生物学的研究十分必要。到目前为止,关于泥蚶的繁殖生物学研究,国内林志强等[1]对人工育苗作过研究,郑家声等[2]研究了山东省乳山湾泥蚶的性腺发育和生殖周期;而对乐清湾泥蚶繁殖生物学研究至今未见详细报道。本研究对浙江省乐清湾围塘泥蚶的繁殖周期及环境因子进行了周年调查,旨在为保护和利用浙江乐清湾泥蚶的种质资源,提高乐清湾泥蚶在养殖及开展良种选育等工作提供基础资料和理论依据,为我国泥蚶的人工育苗技术深入研究提供科学依据。
1 材料与方法
1.1 样品采集 自2010年3月至2010年8月于浙江乐清湾沿海围塘采集泥蚶,每月下旬一次,每次随机采集100-120个体,将样品鲜活运回实验室,并现场测定取样海区海水的盐度和温度,同时采集采样地海水样品,采用分光度法[3]测定叶绿素a含量。
1.2 肥满度 每月取30个个体解剖,测量壳高,壳长,壳宽,壳重,软体部重,将软体部和壳分别在105℃烘箱恒温烘干至恒重,称壳干重,软体干重,计算肥满度。
1.3 组织学 每个月取90-100个泥蚶个体,对样品进行解剖、观察,并将切取的一部分性腺组织用波恩氏液固定,常规石蜡包埋组织切片(切片厚度6μm),苏木精-伊红(H-E)染色。在显微镜下观察每个月份泥蚶的性腺发育状况。每个月份取5-10个雌性个体,每个个体观察、测量50-100个卵母细细胞的直径、求平均值以确定泥蚶性腺的成熟度。
1.3.1 试剂 无水乙醇(A.R,安徽安特生物化学有限公司)、二甲苯(A.R,无锡市佳妮化工有限公司)、苦味酸(A.R,中国医药集团上海化学试剂公司)、甲醛(A.R,衢州巨化试剂有限公司、冰醋酸(A.R,国药集团化学试剂有限公司)、石蜡(熔点56-58℃,国药集团化学试剂有限公司)、明胶、中性树胶(国药集团化学试剂有限公司)。
1.3.2 实验工具和仪器 手术剪,镊子,解剖刀,称量纸,酒精灯,量筒,样品瓶,擦镜纸,载玻片,盖玻片:用100%酒精浸泡24h,酒精灯灼烧。
包埋机(全自动生物包埋机YD-6D)、切片机(TD-1508A轮转式切片机)、烤片机(YD-B烤片机)、脱水机(生物组织脱水机YD-12G)、显微镜(荧光显微镜Nikon Eclipse80i,生物显微镜Nikon YS100)、染色机(电脑自动染色机YAB0700)、电子天平、离心机、分光光度计等。
1.3.3 实验方法与步骤
①固定、保存。固定剂选用波恩(Bouin)液,固定24h,后换多次70%乙醇,褪色,直至洗去黄色。
②脱水。样品退去黄色后依次放入80%酒精1h,95%酒精 30min,95%酒精 30min,100%酒精 30min,100%酒精30min。
③透明。样品依次放入1/2纯酒精1/2二甲苯1h,二甲苯Ⅰ20min,二甲苯Ⅱ20min。
④浸蜡。样品依次放入1/2二甲苯1/2石蜡1h,纯蜡Ⅰ1h,纯蜡Ⅱ1h。
⑤包埋。包埋过程中要注意尽量避免产生气泡而影响切片效果。
⑥切片、贴片、展片。采用轮转式切片机,切片厚度为6 μm,后将切好的蜡片用1%明胶贴于干净的载玻片上,加几滴蒸馏水,酒精灯烘烤展片。
⑦染色。将烘干的切片依次放入二甲苯10min,1/2纯酒精1/2二甲苯4min,100%乙醇4min,95%乙醇4min,80%乙醇4min,70%乙醇4min,50%乙醇 4min,蒸馏水 4min,苏木精8min,染色后用自来水6min,酸酒精1min分色,50%乙醇 4min,95%乙醇 4min,曙红 30s,90%乙醇 15s,100%乙醇Ⅰ2min,乙醇Ⅱ2min,1/2纯酒精1/2二甲苯4min,二甲苯10min,二甲苯15min,取出切片后,立即用中性树胶封片。
⑧烘干。封片后的切片自然风干或烘干。
⑨挑选比较清晰的切片进行显微镜观察、拍照。
1.4 数据分析 数据统计分析采用SPSS 17.0软件处理。对条件指数和卵母细胞直径的测量数据用单因素方差分析(ANOVA)进行显著性检验,月间的差异采用Turkey检验(P<0.05),图片用 Adobe Photoshop CS 8.0.1 软件处理。
2 结果与分析
2.1 环境因子的周年变化 图1显示了采样围塘海水水温和盐度的周年变化。其中水温呈明显的季节性变化,在6月份的时候达到最高值(29.8℃),随后逐渐降低,至12月达到最低(4.5℃),之后温度回升。盐度为10.0-30.0,年平均值为22.7,7月和8月波动明显,与研究期间2010年3-7月浙江乐清湾正值雨季,雨水较多有关。
图2显示了叶绿素a含量的周年变动模式。表现为春、夏季高,秋、冬季低的特点。3月份叶绿素a含量逐渐升高至6月达到最高值,含量为22.60μg/L。8月迎来次高峰,叶绿素a含量为22.43μg/L。随后叶绿素a的含量逐渐下降,并于12月降至全年最低值,为2.43μg/L。此后,叶绿素a含量开始上升。
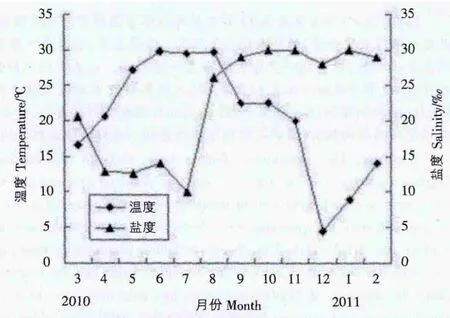
图1 取样围塘水温和盐度的周年变化
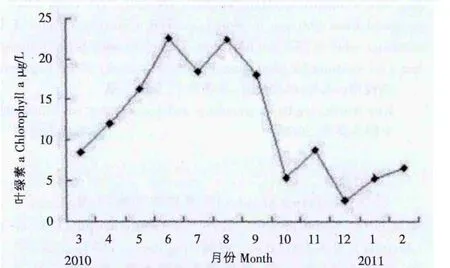
图2 取样围塘叶绿素a含量的周年变化
2.2 泥蚶生物学性状和肥满度的周年变化 表1为泥蚶的生物学性状,月平均壳高的变化范围为20.12-23.33 mm,月平均壳长为26.04-30.11mm,月平均壳宽为17.55-21.64mm,月平均软体部湿重在1.12-2.65g之间变化,在12月,软体部的湿重和干重都达到了最低值,分别为1.12g和 0.14g。
图3显示了泥蚶肥满度的周年变化,波动不大,3月份起,肥满度平均值开始增大,增殖期开始,5月份达到最大值7.74×10-2,此时正值卵巢内的卵母细胞的大生长期,随着成熟期和排放的来临,肥满度迅速下降,8月份达到最低值3.25×10-2,9月份为3.42×10-2,以后各月逐渐平缓稳定,12月为3.30×10-2,此后又开始上升。
2.3 泥蚶繁殖周期 泥蚶配子发育和卵径存在周年变化,泥蚶的配子发育具有明显的季节性(图5),卵径月间差异显著(P<0.05)。
参考Ivell,结合泥蚶性腺的组织学和外部形态特征,将泥蚶的性腺发育分为5个时期:增殖期,生长期,成熟期,排放期和休止期(图4)。
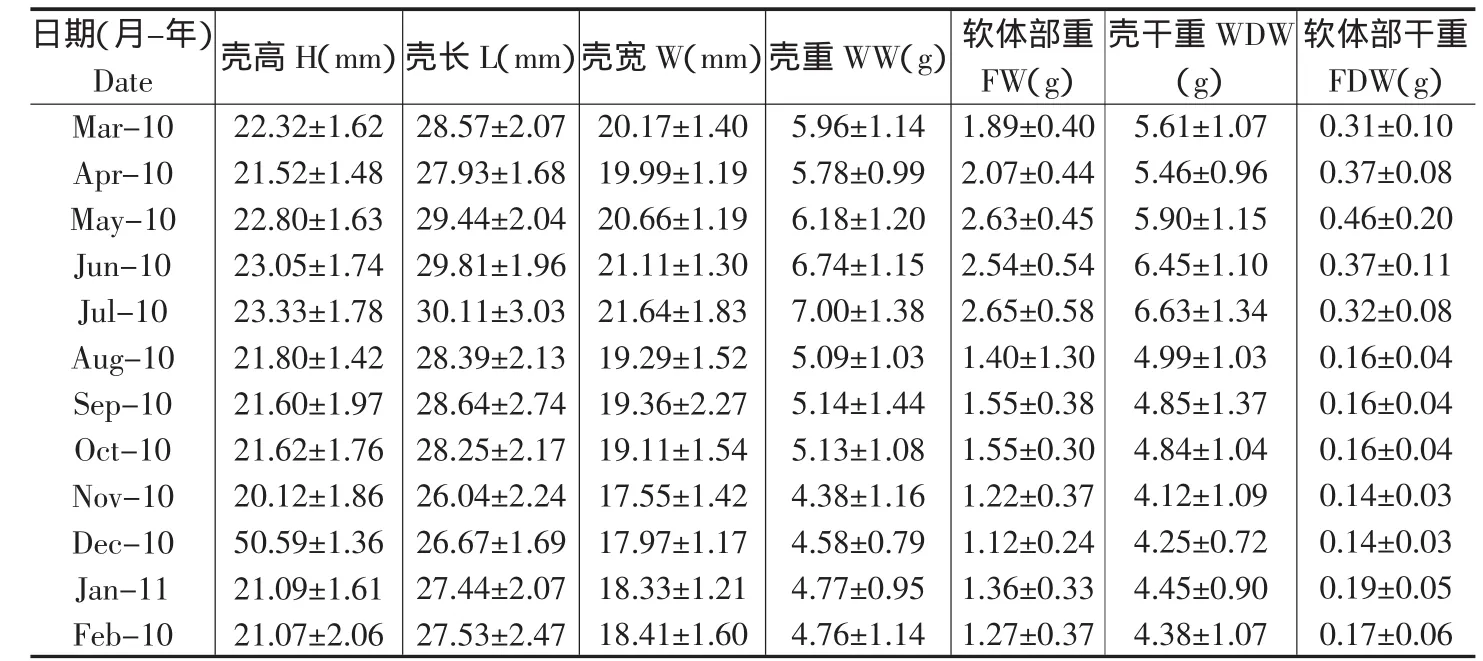
表1 泥蚶的生物学性状。平均值±标准差,n=30
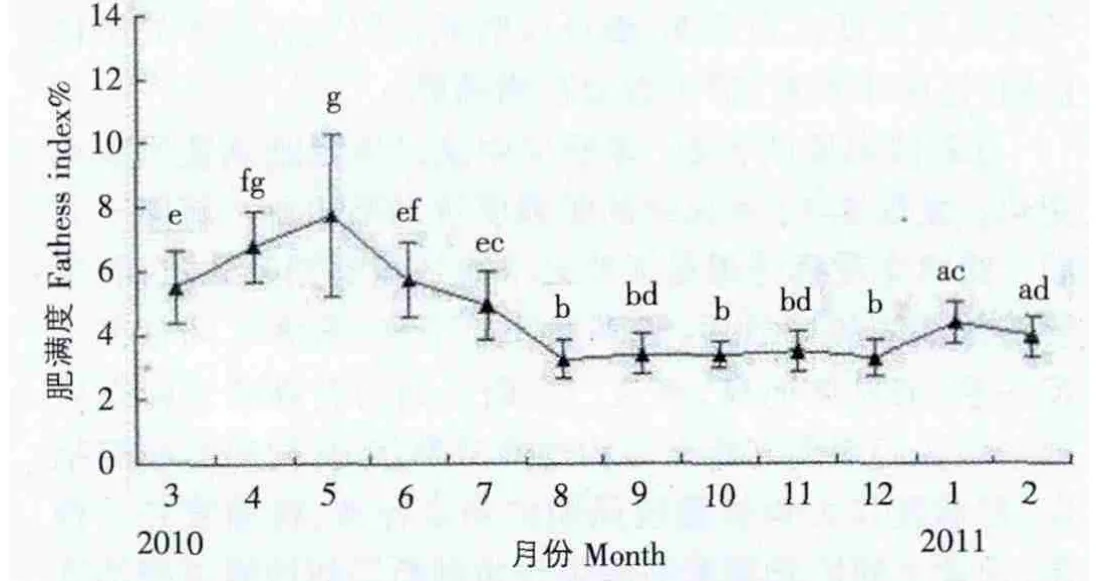
图3 泥蚶肥满度的周年变化
第Ⅰ期,即增殖期,本期发育时间为3月初至4月底,随着海区水温的回升,此期性别刚刚分化,月平均水温为16.6-20.6℃。组织切片观察,雌性滤泡开始出现,滤泡腔大,滤泡壁由单层上皮构成,配子发育刚刚开始。随着滤泡壁开始增厚,卵巢内出现单层不连续的卵原细胞,卵原细胞处在活跃增殖时期,胞体呈卵圆或三角形(图4Ⅰ-O),3月份平均直径约8.87μm,为全年最低值,至4月份,平均卵径值可增至16.99μm(图5),3月57.84%的个体处于Ⅰ期,4月74.47%的个体处于Ⅰ期(图5)。雄性滤泡开始出现,体积小壁薄不规则,滤泡间结缔组织丰富。滤泡壁上形成多层的精原细胞,数量逐渐增多,精原细胞呈圆形或三角形(图4Ⅰ-T)。
第Ⅱ期,生长期,滤泡数量增加,体积增大,分布范围广。卵原细胞继续分裂增殖,雌性滤泡腔内空隙逐渐减少,卵原细胞逐渐分化,其一端已明显地突向滤泡腔,呈长形或倒梨形有少量成熟卵子出现(图4Ⅱ-O),本期发育时间主要为5月份,有60.78%的个体处于生长期,并有少量个体(8.82%)性腺发育成熟(图5),此期平均卵径为34.08 μm,水温为27.2℃。雄性滤泡数量增多,体积增大,壁加厚,颜色加深,可观察到精原细胞到精子各个生长阶段,少量精子形成,至本期中,精原细胞约占滤泡截面积的50%以上(图4Ⅱ-T)。
第Ⅲ期,成熟期,配子发育结束,产卵还未发生或刚刚开始。本期发育6月和7月为成熟期,性腺发育成熟的个体分别占42.50%和58.06%(图5),有部分个体开始产卵,平均水温为29.5-29.8℃。本期滤泡为全年最丰满,仅少量结缔组织存在,滤泡间的间隙基本消失。游离的成熟卵约占80%以上,卵子互相挤压,成不规则形状,有椭圆形、梨形、多边形等(图4Ⅲ-O),卵径在6月份为42.20μm,7月卵径达最大值,为 48.47μm(图 5)。本期精巢达到全年最丰满阶段,滤泡腔内充满了成熟的精子,精子为鞭毛型,呈辐射状排列,精子头部朝向滤泡壁,尾部朝向滤泡腔(图4Ⅲ-T)。
第Ⅳ期,排放期,自7月下旬开始,滤泡内生殖细胞逐渐减少,开始出现大小不等的空腔,滤泡腔体积逐渐减小,雌雄排放期和成熟期均有重叠现象存在。雌性滤泡壁上仅剩少量卵母细胞,有些滤泡壁破裂,滤泡腔内残存少量成熟卵子(图4Ⅳ-O),此期发育时间为7月下旬至 8月下旬,8月为排放鼎盛期,8月卵径降低至18.57μm,部分个体产卵期会延续到11月份;雄性滤泡开始出现大小不等辐射状的腔,滤泡萎缩,精子数量明显减少,滤泡腔内仍有少量精母细胞和精子存在(图4Ⅳ-T)。
第Ⅴ期,即休止期,本期发育时间为9月至翌年2月,个别为排放期和增殖期,此期间,仅4.91%个体可辨雌雄,显微镜观察卵径减小现象明显,故未进行测量计数。切片观察滤泡空虚,变成一个大空腔,形状不规则,滤泡壁薄,随后滤泡逐渐变小直至消失,结缔组织占据了整个性腺(图4Ⅴ-ND),性腺发育完成一个周期。图4所示为泥蚶性腺发育分期。
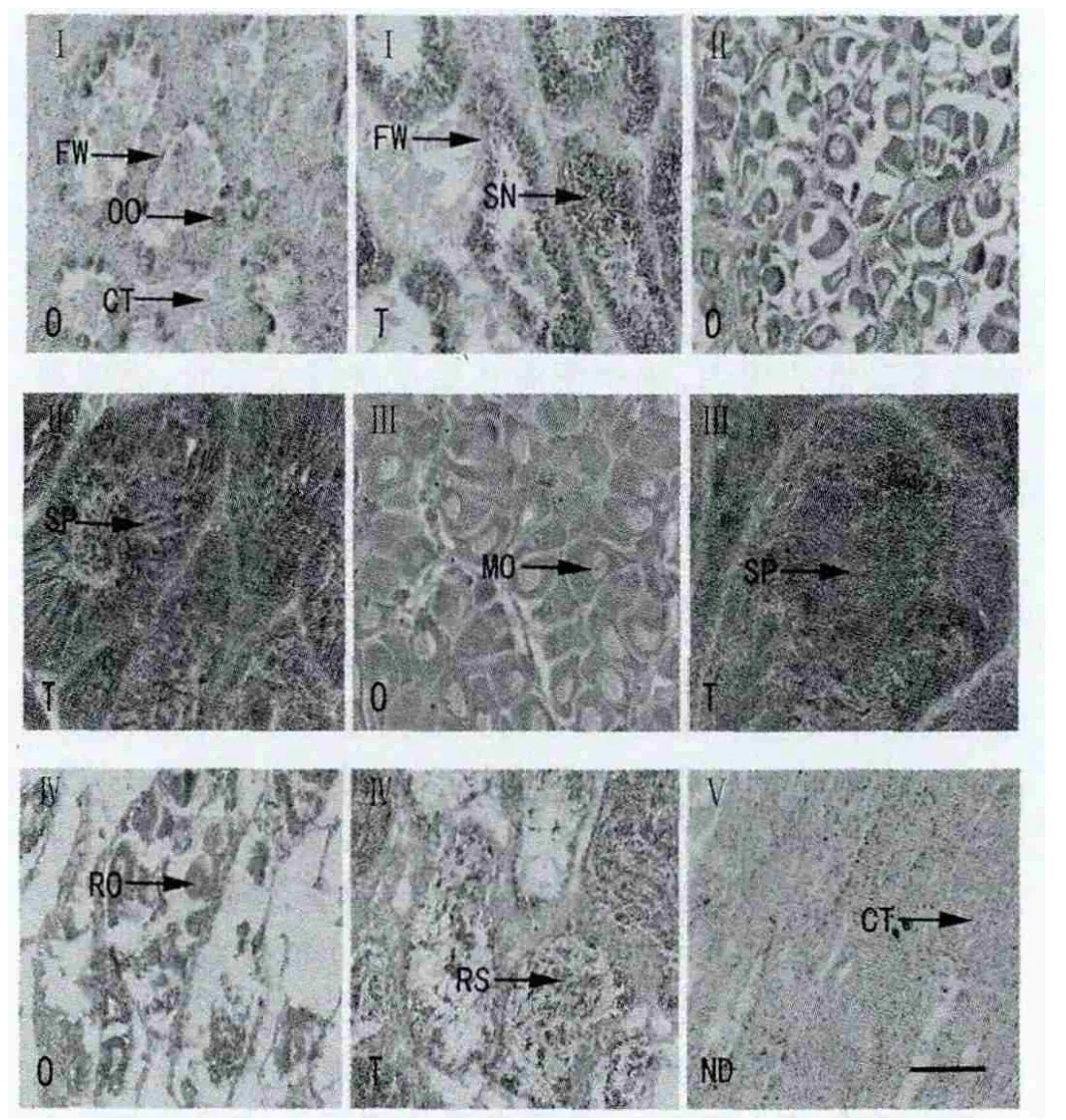
图4 泥蚶性腺发育分期
3 讨论
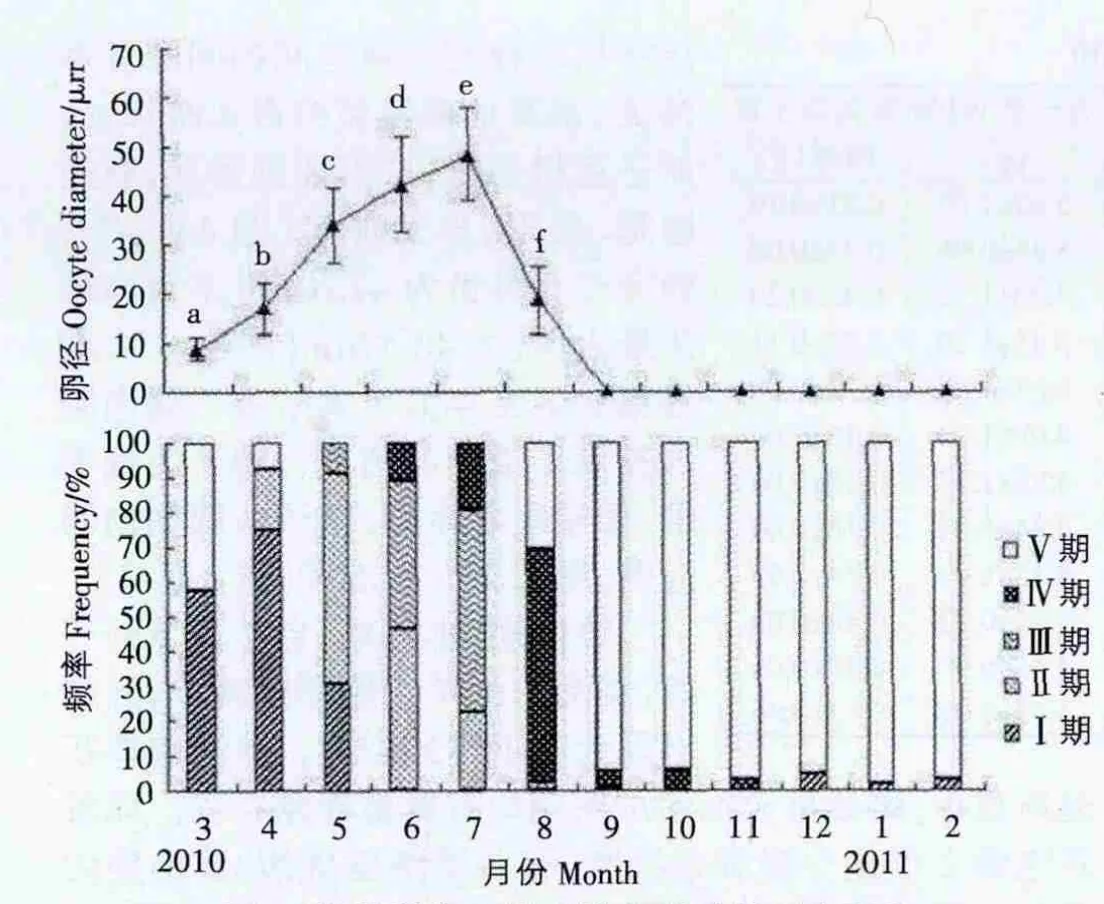
图5 泥蚶卵径和性腺发育状况的周年变化
3.1 泥蚶的性腺发育周期与环境因子的关系 海洋贝类的繁殖周期受外部因素(温度,盐度,食物丰度等)和内部因素(神经内分泌系统调节等)共同影响[5]。贝类的繁殖策略可以认为是对周围环境(如温度和盐度)和食物的可得性等的一种适应,因环境随海区差异变化较大,不同海区海洋贝类性腺开始发育和产卵时间是不同的[6,7]。
3.1.1 水温与繁殖周期关系 本实验结果显示,乐清湾围塘大部分泥蚶配子发育的开始时间为3-4月,此时水温16.6-20.6℃。本实验的组织学结果显示,5月份50%以上的个体的泥蚶处于Ⅱ期,并有部分个体性成熟,6月开始有个体产卵、排精,此时水温为29.8℃,7月后大规模产卵、排精。由此可见,泥蚶产卵所需温度要高于配子发育启动所需温度。
3.1.2 盐度与繁殖周期关系 水域中的盐度也会影响泥蚶的繁殖周期,泥蚶对盐度适应能力强,成蚶在盐度为10.4-32.5之间的海水中均能正常生活,不过在河口或海水盐度较低的内湾,雨季使海水盐度降得过低时,泥蚶则向下层潜伏,出入水孔被淤闭,使其在泥层中受到保护,待盐度上升后,泥蚶再回到上层,进行正常生活。样品都是从淤泥里采集,观察泥蚶存活情况,有个别死亡现象,9月开始雨水天气减少,盐度保持在30左右,采样发现泥蚶存活率保持在较高水平,9月开始值翌年雨季前,存活率均超过95%。本实验中,盐度在3月到8月变化很大,在10.02-26.16之间波动,特别是6月中旬到8月上旬,盐度一直维持在较低水平(10-14),主要原因是这段时间降雨过久,而乐清湾又属内湾,盐度受降雨量影响明显,实验期间浙江省6月中旬至7月中旬经历2000-2010年少有的典型梅雨季节,而8月下旬盐度增至26.16,并逐渐稳定,泥蚶于8月进入产卵鼎盛期,据此判断乐清湾泥蚶产卵具有雨季之后盐度较高时开始大规模产卵的特点,该规律与Suja等[8]对分布在印度的两个不同地区的婴儿蛤的研究结果基本一致。而国内的报道鲜有提到盐度对泥蚶繁殖的影响,因此两者之间的关系有待进一步研究。
3.1.3 食物与繁殖周期关系 本实验对取样围塘的叶绿素a的含量进行了周年测定,3-6月叶绿素含量呈现快速增加的趋势,且含量总体上处于较高水平,结合组织学观察结果,乐清湾海区围塘养殖泥蚶在3月至4月配子开始发育,而春季的叶绿素a的值也很高,饵料丰富,且温度也超过发育起始最低温度,符合性腺发育规律。泥蚶的配子发育不仅需要起始最低温度(性腺发育生物学零度),也需要充足的饵料保障。6月份泥蚶配子发育进入生长期,叶绿素a的值达到最高(22.6μg/L),饵料最丰富,为泥蚶发育提供了良好的食物基础,有重要的促进作用。8月泥蚶已进入排放鼎盛期,而叶绿素a的值达到次高峰(22.43 μg/L),水中浮游植物数量很大,平均温度达次高峰29.7℃。9月到翌年2月,取样围塘海水叶绿素a含量较低,水中浮游植物数量很少,并且由于雨雪天气,夏季至秋冬季温度降低尤为迅速,致此段时期泥蚶性腺基本处于休止期,性腺不发育,极少数处在增殖期。
3.2 成熟度的判断 本研究中通过测定肥满度的周年变化,发现8-12月泥蚶的肥满度波动不明显,而同一时期,组织学观察结果是泥蚶处于排放鼎盛期及休止期,尽管排放期间和排放后,肥满度开始下降,两者之间存在一定关系,但总体而言,在这一时期泥蚶的肥满度未能很好的帮助我们准确判断泥蚶的性腺成熟状况,同时有研究指出,肥满度作为鱼类繁殖周期的判定标准,精确度并不很高,因此泥蚶的肥满度不建议作为判断泥蚶性腺成熟的直接标准。
3.3 雌雄同体问题 瓣鳃类中的雌雄同体现象较普遍[9,10]。本研究在1243个泥蚶性腺切片观察结果表明,雌雄比例为1:1.13,雄性略多于雌性,与吴洪喜等[7]观察相符,与郑家声等[2]对418颗生长在山东青岛市薛家岛镇唐岛湾的泥蚶观察结果,雌雄比l:1.16很接近,本研究中未发现泥蚶雌雄同体现象,相关报道[2]也未提及发现雌雄同体现象,因此可以断定泥蚶为雌雄异体贝类,不存在雌雄同体现象。
4 结论
本文对浙江省温州市乐清湾围塘养殖的泥蚶的繁殖生物学性状、繁殖周期及其与环境因子(温度、盐度和叶绿素a含量)的关系进行了研究。泥蚶为雌雄异体,其性腺发育受到饵料丰富度的影响成周年变化,同时泥蚶产卵时所需温度要高于配子发育启动所需温度,且具有雨季之后盐度较高时开始大规模产卵的特点。泥蚶配子发育始于3-4月,配子主要在6、7月成熟,大规模产卵主要在8月份,雌雄异体。泥蚶卵径的周年变化是判断泥蚶成熟度的良好指标。海洋贝类的繁殖周期受外部因素和内部因素共同影响,要真正了解控制泥蚶繁殖周期的生理机制还需要对影响其繁殖周期的内部因素(神经内分泌系统调节等)做大量的工作。
[1]林志强,汪德清,徐桂仙等.泥蚶人工育苗试验报告[J].浙南水产科技,1980,1:1-11.
[2]郑家声,王梅林,王志勇等.泥蚶的性腺发育和生殖周期[J].青岛海洋大学学报,1995,25(4):503-510.
[3]戴玉蓉,卢敬让.养殖水体浮游植物叶绿素a的简易测定[J].齐鲁渔业,1997,14:35-36.
[4]Ivell R.The biology and ecolog y of a brackish lagoon bivalve,Cerastoderma glaucum Bruguiere,in an English lagoon,the widewater,Sussex[J].J Moll Stud,1979,45:383-400.
[5]Berthelin C,Kellner K,Mathieu M.Storage metabolism in the Pacific oyster (Crassostrea gigas)in relation to summer mortalities and reproductive cycle(west coast of France)[J].Comp Biochem Physiol,2000,125 B:359-369.
[6]Ahn I Y,Surh J,Park Y G,et al.Growth and seasonal energetics of the Antarctic bivalve Laternula elliptica from King George Island,Antarctica[J].Mar Ecol Prog Ser,2003,257:99-110.
[7]Dridi S,Romdhane M S,Elcafsi M.Seasonal variation inweight and biochemicalcomposition ofthe Pacific oyster,Crassostrea gigasin relation to the gametogenic cycle and environmentalconditions ofthe Bizertlagoon,Tunisia [J].Aquaculture,2007,263:238-248.
[8]Suja N,Muthiah P.The reproductive biology of the baby clam,Marcia opima,from two geographically separated areas of India.Aquaculture,2007,273:700-710.
[9]楼允东.组织胚胎学[M].北京:中国农业出版社,1996:223-238.
[10]陈品健,游岚,倪法鸿.尖刀蛏的生殖周期[J].海洋科学,1994,18(4):38-42.
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21
生物学通报(2021年11期)2021-09-28
模具制造(2019年4期)2019-12-29
传媒评论(2018年1期)2018-03-22
中华戏曲(2016年2期)2016-01-22
应用海洋学学报(2015年4期)2015-11-24
食品工业科技(2014年15期)2014-03-11
微创泌尿外科杂志(2014年4期)2014-02-28
中学理科·综合版(2008年9期)2008-10-15
中国美容医学(2004年3期)2004-09-17
