
泉州湾污损生物生态研究
2014-10-26 01:17林和山王建军郑成兴林俊辉黄雅琴何雪宝李荣冠郑凤武江锦祥
海洋学报 2014年4期
林和山,王建军*,郑成兴,林俊辉,黄雅琴,何雪宝,李荣冠,郑凤武,江锦祥
(1.国家海洋局 第三海洋研究所 海洋生物与生态实验室,福建 厦门361005)
1 引言
海洋污损生物对海洋经济开发的负面影响早已引起人们的广泛关注,其危害包括增加船舶阻力、堵塞管道、加速金属腐蚀、破坏仪器设备的灵敏度和寿命等方面。从20世纪40年代开始,科学家们便对船舶[1]、浮标[2-4]、油气钻井平台[5-7]、养殖网箱[8-9]和其他人工设施[10-11]的污损生物进行研究和分析,涉及分类鉴定、附着机理、防除技术、污损生态效应及外来物种等方面的内容[12-30]。
泉州湾位于福建省东南部沿海,是台湾海峡西岸中部的一个重要港湾,属于开敞式海湾,口门中部有大、小坠岛横亘,湾内受晋江和洛阳江水的影响较大,但湾口主要受外海水系控制,年平均水温20.4℃,7月最高,为28.3℃,1、2月最低,为11.9℃,平均盐度26.466。泉州湾不仅是鱼、虾、蟹、贝和藻类的主要养殖基地,也是福建省非常繁忙的港口之一。
黄宗国等[2]虽于19世纪60年代在泉州湾初步进行了污损生物生态调查,但50 a来未见进一步探讨。本文通过周年挂板试验,借助多元统计分析方法,对该海区污损生物群落较长尺度的时空变化特征进行了系统研究,以期为海洋生态研究积累数据资料,并为船舶、码头、网箱、管道等海洋设施的污损生物防除提供科学依据。
2 材料与方法
2001年12月—2002年11月在泉州湾海域进行污损生物周年挂板试验,挂板位置如图1所示。采用15 c m×15 c m×2 c m的水泥试板分2组挂放,月板、季板、半年板和年板的收放周期分别按逐月、每季度、每半年和周年进行。每组试板分表、底两层挂放,试板面与海面垂直,表层试板上沿与水面齐平,底层试板置于水深3.5 m处。全年共回收试板76块,回收率100%。标本处理及室内分析和资料整理均按《海洋调查规范》[31]要求进行。
采用 Margalef物种丰富度指数(d)、Shannon-Wiener多样性指数(H′)、Pielou均匀度(J)以及Simpson物种优势度(D)进行群落多样性分析,公式如下:

式中,N为样品中所有种类的总个体数目,S为样品中的种类总数,Pi为第i种的个体数与样品中的总个体数的比值(ni/N)。

图1 泉州湾海域污损生物挂板试验点
丰度数据经四次方根转化及标准化后,在Bray-Curtis相似性测量的基础上,组合使用等级聚类(Cluster)(组平均连接法)来分析群落结构的相似性,并采用丰度/生物量比较法分析生物群落的变化,物种丰度为物种在各站位的平均分布密度,如某站没有出现该种,则不计入,另外,因藻类和苔藓虫难以计数其个体数,故只计算其生物量而不计算其密度,采用PRI MER 5.0软件进行上述指数的计算。
3 结果
3.1 种类组成及主要优势种
本次调查共记录污损生物11门112种,其中,多毛类最多(39种),其次是甲壳类(26种)和软体动物(23种),三者占总种数的78.6%;还有藻类6种、刺胞动物7种、扁形动物2种、纽形动物2种、苔藓动物5种、棘皮动物1种和海鞘类1种(见附表1)。长鳃麦秆虫(Caprellaequilibra)和网纹藤壶(Amphibalanusreticulates)在泉州湾污损生物群落中居绝对优势,其中前者的出现频率、月平均附着密度和生物量分别为90.8%、114 063个/m2和260 g/m2(见图2),后者为85.5%、38 932个/m2和1 727 g/m2(见图3)。另外,裂片石莼(Ulvaf asciata)、中胚花筒螅(Tubularia mesembryanthemum)、薮枝螅(Obeliasp.)、纵条矶海葵(Halipl anellal uciae)、外伪角涡虫(Pseudocerosexoplatus)、柄涡虫(Stylochussp.)、僧帽囊牡蛎(Saccostreacucull ata)、翡翠贻贝(Per naviridis)、巴西地钩虾(Podocerusbrasiliensis)、强壮板钩虾(Stenothoe valida)、镰形叶钩虾(Jassaf alcata)和角突麦秆虫(Caprell ascaur a)等种类的出现频率也较高(22.4%~90.8%),且附着数量较大。
3.2 附着量及其时空变化
泉州湾全年各月均有生物附着,但附着种类和数量的季节性差异明显,附着盛期为4—11月,12—2月为附着淡季。长鳃麦秆虫在全年各月均有附着,且附着密度极大,2—7月为其附着高峰期,5月的平均附着密度可达481 775个/m2;除了2月份,网纹藤壶在其余月份均有附着,附着数量很大,5—10月为其附着盛期,两次附着高峰出现在8月和10月,8月的平均附着生物量可达9 845 g/m2;裂片石莼、中胚花筒螅、镰形叶钩虾和角突麦秆虫等种类适宜较凉爽的月份,而薮枝螅、外伪角涡虫、翡翠贻贝和巴西地钩虾等则较喜好高温的月份(见图4)。
泉州湾污损生物群落附着量的季节差异明显。月板的平均附着密度和生物量分别为264 917个/m2和2 624 g/m2,因长鳃麦秆虫(表层板)和角突麦秆虫(底层板)的大量附着,4月的附着密度达到最高(807 781个/m2),而8月的附着生物量最大(10 945 g/m2),覆盖面积达100%,这与网纹藤壶的密集附着有关;季板的平均附着密度和生物量分别为197 872个/m2和5 309 g/m2,春季附着密度最高(511 769个/m2),而夏季的附着生物量最大(11 295 g/m2),覆盖面积比例也达100%,主要贡献种类分别是长鳃麦秆虫和网纹藤壶;半年板的平均附着生物量为6 210 g/m2,下半年的种类数更是高达59种,附着强度也大于上半年;周年板的平均附着生物量达到8 247 g/m2,覆盖面积比例达100%,主要优势种为翡翠贻贝和网纹藤壶。详见图5、6和表1。
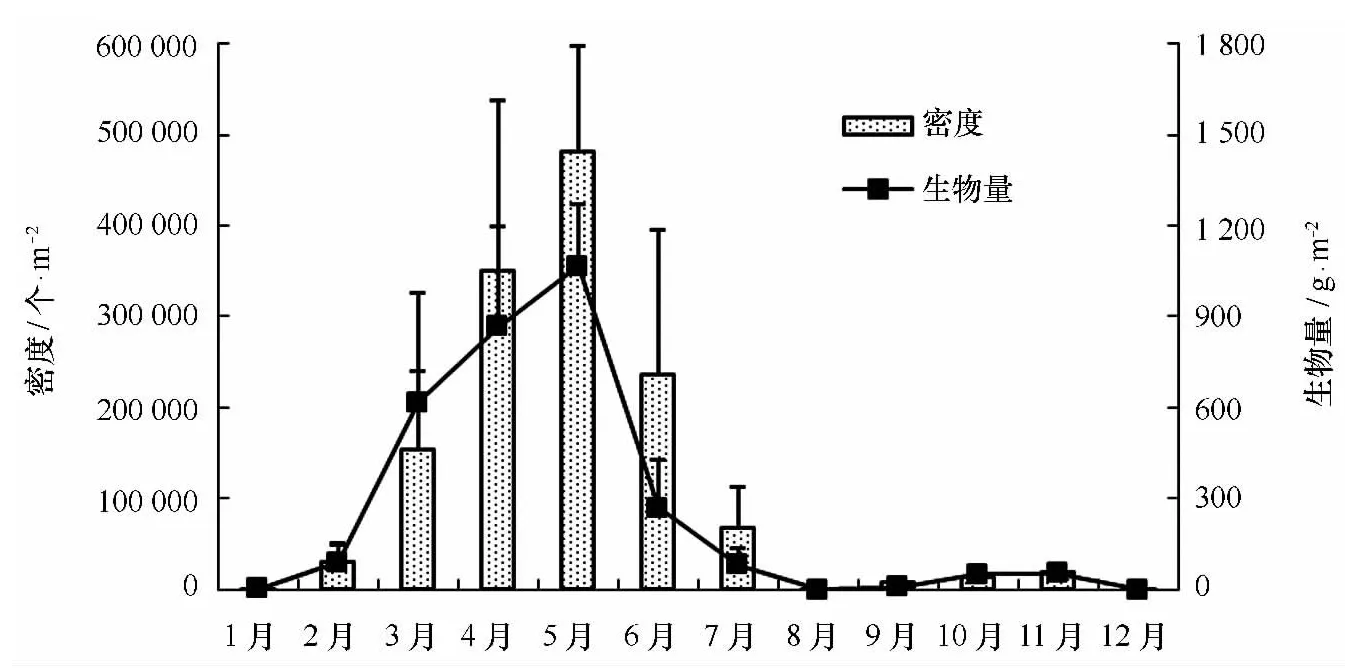
图2 泉州湾长鳃麦秆虫密度和生物量的逐月变化情况

图3 泉州湾网纹藤壶密度和生物量附着的逐月变化情况

图4 泉州湾污损生物优势种和主要种的附着季节
3.3 群落的季节更替
依据Bray-curtis相似性系数聚类分析和多维尺度排序,泉州湾的污损生物可分成3个群落类型(见图7、图8):群落A,中胚花筒螅-镰形叶钩虾-长鳃麦杆虫-角突麦杆虫群落,该群落具有附着密度大、优势度高等特点,较适宜凉爽的月份,以水螅和端足类为优势附着类群;群落B,中胚花筒螅-网纹藤壶群落,适宜水温较低的冬季,该群落种类较少,网纹藤壶占绝对主导;群落C,裂片石莼-翡翠贻贝-网纹藤壶-巴西地沟虾-长鳃麦秆虫群落,该群落附着周期较长,种类丰富,附着密度和生物量均较高,较适宜炎热的夏季,以无柄蔓足类和端足类为主。

图5 泉州湾表层污损生物的密度和生物量

图6 泉州湾底层污损生物密度和生物量

表1 泉州湾污损生物的数量组成
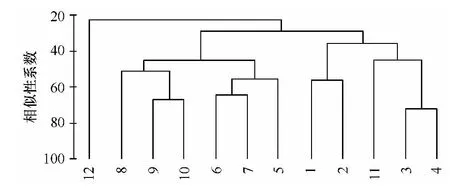
图7 泉州湾污损生物群落Bray-Curtis相似性聚类
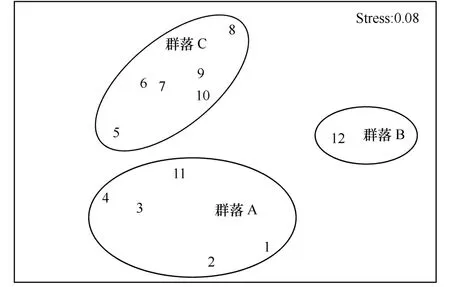
图8 泉州湾污损生物群落多维尺度排序
3.4 群落多样性
泉州湾季板上污损生物的丰度指数d值平均为2.11,秋季最高,周年板的d值为3.62,下半年大于上半年;多样性指数H′值和均匀度指数J值四季平均分别为1.09和0.24,秋冬二季大于其他两季,下半年大于上半年,周年板的H′值和J值分别为2.54和0.47;优势度指数D值四季平均为0.64,春夏二季大于秋冬二季,上半年大于下半年,周年板的D值为0.23(表2)。丰度指数d、多样性指数H′和均匀度指数J均基本符合秋冬高、春夏低、下半年大于上半年的规律,而优势度指数D则相反。

表2 泉州湾污损生物群落物种指数
3.5 群落结构的稳定性
随着试板浸海时间的延长,半年板和周年板上的污损生物群落多样性明显增加。用丰度生物量比较法(ABC曲线)对泉州湾季板、半年板和周年板的污损生物群落结构进行分析(见图9,Ⅰ~Ⅶ),结果表明4个季度和上、下半年的污损生物群落结构均不稳定:春季、秋季、冬季、上半年和下半年的丰度生物量复合k-优势度曲线出现翻转、交叉或重叠(Ⅰ、Ⅲ、Ⅳ、Ⅴ和Ⅵ),这与个体小质量轻的长鳃麦秆虫和网纹藤壶等小型甲壳类的暴发性增殖有关;夏季的生物量累积百分优势度达96%(Ⅱ),这与网纹藤壶的大量附着有关,其生物量可达10 770 g/m2,占总生物量的95.4%;周年板的生物量优势度曲线始终位于丰度上方(Ⅶ),表明周年板上污损生物群落结构相对稳定。
4 讨论
泉州湾地处近岸港湾内,盐度比河口港高,同时受到外海水系和沿岸水系的制约,其污损生物种类组成以近岸暖水种为主,为典型的亚热带内湾群落。该海域群落组成与东山湾、洋山港、大亚湾、三沙湾、海坛海峡和厦门港[27,30,32-33,36]相近,网纹藤壶是它们共有的压倒性优势种,但本次网纹藤壶的附着量远远高于其他港湾,这可能是因为本次的挂板位置位于泉州湾口门,水流较为通畅,有利于网纹藤壶的附着;而有别于舟山朱家尖海域、闽江口、沙埕港和汕头港[26-27,34-35]的污损生物群落,这些港湾的平均盐度较低,且月平均盐度变化较大,以河口低盐种泥藤壶(Balanusuliginosus)为主要代表种;也与台湾海峡东岸的八斗子港和大鹏湾的污损生物群落有显著不同[36],这主要是因为这两个港湾均处于高盐和潮差较小的海域(不规则半日潮,潮差仅为0.4 m)。

图9 泉州湾污损生物群落丰度生物量复合k-优势度曲线
泉州湾污损生物的附着特点是种类丰富、附着强度大。本次调查共记录污损生物112种,明显高于舟山朱家尖海域(85种)、洋山港(19种)、沙埕港(70种)、三沙湾(50种)、海坛海峡(71种)、闽江口(43种)、泉州湾(57种)、厦门港(91种)、东山湾(102种)和汕头港(72种),以及台湾海峡东岸的八斗子港和大鹏湾(共60种),但低于大亚湾(198种)。泉州湾污损生物的种类组成与东山湾[30]较为相似,但泉州湾的附着强度(月板和季板的平均附着生物量分别为2 624 g/m2和5 309 g/m2)明显高于东山湾(月板和季板的平均附着生物量分别为979 g/m2和4 142 g/m2),泉州湾夏季板的附着生物量可达11 295 g/m2,为四季最高,附着强度在中国港湾污损生物中属于2级[27]。
另外,本次调查与1962—1963年黄宗国等在泉州湾的研究[27]不同的是,后者主要以泥藤壶和双节螅(Bimeriasp.)等河口低盐种为主,污损生物种类数和附着强度也明显低于本次调查,这主要是因为1962—1963年的调查位置在泉州湾内(见图1),受江水(洛阳江和晋江)影响其盐度和水质变化较大。实验结果表明泉州湾污损生物群落组成与水温、盐度以及水流畅通程度有一定关系。
5 结论
泉州湾污损生物附着的特点是种类丰富、附着强度大。本次调查共记录污损生物112种,以近岸暖水种为主,是典型的亚热带内湾群落类型,其中网纹藤壶和长鳃麦秆虫是该海域最具代表性的优势种;该海区污损生物全年均可附着,附着量的季节差异明显,附着盛期在4—11月,月板、季板和周年板的平均附着生物量分别为2 624、5 309和8 247 g/m2,夏季板的附着生物量可达11 295 g/m2,为四季最高,附着强度在中国港湾污损生物中属于2级。
该海区的污损生物可分成3个群落类型,即群落A(中胚花筒螅-镰形叶钩虾-长鳃麦杆虫-角突麦杆虫群落)、群落B(中胚花筒螅-网纹藤壶群落)和群落C(裂片石莼-翡翠贻贝-网纹藤壶-巴西地沟虾-长鳃麦秆虫群落),随着试板浸海时间的延长,试板上污损生物群落的多样性明显增加,且结构也趋于稳定。实验结果表明泉州湾污损生物群落组成与水温、盐度以及水流畅通程度有一定关系。

附表1 泉州湾污损生物种名录
[1]Cristiane Maria Rocha Farrapeira,Arthur Vinicius de Oliveira Marrocos de Melo,Débora Ferreira Barbosa,et al.Ship hull fouling in the port of Recife,Pernambuco[J].Brazilian Journal of Oceanography,2007,55(3):207—221.
[2]Woods Hole Oceanographic Institution.Marine fouling and its prevention[M].US Naval Institute,1952:1—388.
[3]Yan T,Yan W X,Dong Y,et al.Marine fouling on floating installations west of Dongsha Islands,the northern South China Sea[J].Inter national Biodeterioration &Biodegradation,2009,63:1079—1087.
[4]郑成兴,黄宗国,李传燕,等.汕头港码头、浮标污损生物[J].海洋学报,1996,18(1):115—124.
[5]Yan T,Yan W X,Dong Y,et al.Marine fouling of offshore installations in the northern Beibu Gulf of China[J].International Biodeterioration &Biodegradation,2006,58:99—105.
[6]Whomersley P,Picken G B.Long-ter m dynamics of fouling communities found on offshore installations in the Nort h Sea[J].Journal of the Marine Biological Association of the UK,2003,83(5):897—901.
[7]Sammarco P W,Atchison A D,Boland G S.Expansion of coral communities within the northern Gulf of Mexico via offshore oil and gas platfor ms[J].Marine Ecology Progress Series,2004,280:129—143.
[8]Greene J K,Grizzle R E.Successional develop ment of fouling communities on open ocean aquaculture fish cages in the western Gulf of Maine,USA[J].Aquaculture,2007,262:289—301.
[9]王建军,黄宗国,李传燕,等.厦门港网箱养殖场污损生物的研究[J].海洋学报,1996,18(5):93—102.
[10]Susanne Q,Hans K,Torleif M.Develop ment of fouling communities on vertical structures in the Baltic Sea[J].Estuarine,Coastal and Shelf Science,2006,67:618—628.
[11]Ahmed M E,Aisha A B.Marine fouling in Suez Canal,Egypt[J].Aquatic Research,2004,30(A):189—206.
[12]Satheesh S,God win W S.Seasonal variability in the recruit ment of macrofouling community in Kudankulam waters,east coast of India[J].Estuarine,Coastal and Shelf Science,2008,79:518—524.
[13]Wang J J,Huang Z G,Lin S,et al.An ecological study of fouling organisms in Beihai Harbour,Beibu Bay,China[M]∥ Morton B.The Marine Biology of the South China Sea,vol.1.Hong Kong:Hong Kong University Press,1993:167—180.
[14]Yan T,Yan W X,Dong Y,et al.Fouling in offshore areas southeast of t he Pearl River delta,nort hern South china Sea[J].Acta Oceanologica Sinica,2003,22(2):201—211.
[15]Yan T,Yan W X.Fouling of offshore structures in China—a review[J].Biofouling,2003,19(suppl.):133—138.
[16]Yan T,Yan W X,Dong Y,et al.Offshore fouling:Investigation methods[J].Acta Oceanologica Sinica,2004,23(4):733—739.
[17]Pequegnat W E,Pequegnat L H.Larval transport,settlement and population structure of offshore biofouling assemblages in the northeastern Gulf of Mexico[C]∥ Acker R F,Brown B F,DePal ma J R,et al.Proceedings of 3rd International Congress on Marine Corrosion and Fouling.Evanston:Northwestern University Press,1973:731—743.
[18]Maruzzo D,Conlan S,Aldred N,et al.Video observation of surface exploration in cyprids ofBalanusamphitrite:the movements of antennular sensory setae[J].Biofouling:The Journal of Bioadhesion and Biofil m Research,2011,27(2):225—239.
[19]Pradhan N N,Gohad N V,Orihuela B,et al.Develop ment of an automated algorith m for tracking and quantifying barnacle cyprid settlement be-havior[J].Journal of Experimental Marine Biology and Ecology,2011,410:21—28.
[20]Wright S,Caldwell G S,Clare A S.Sper m activation in acorn barnacles by elevation of seawater p H[J].Invertebrate Reproduction and Development,2012,56(1):79—85.
[21]Nogata Y,Tokikuni N,Yoshimura E,et al.Salinity limitations on larval settlement of four barnacle species[J].Sessile organisms,2011,28:47—54.
[22]Tasso M,Conlan S L,Clare A S,et al.Active enzy me nanocoatings affect settlement of Balanus amphitrite barnacle cyprids[J].Advanced Functional Materials,2012,22(1):39—47.
[23]Zhang Y F,Wang G C,Xu Y,et al.The effect of butenolide on behavioural and morphological changes of marine fouling speciesBalanusamphitriteandBugulaneritina[J].Biofouling:The Journal of Bioadhesion and Biofil m Research,2011,27(5):467—475.
[24]Yamaguchi T,Prabowo R E,Ohshiro Y,et al.The introduction to Japan of t he Titan barnacle,Megabalanus coccopoma(Dar win,1854)(Cirripedia:Balanomorpha)and t he role of shipping in its translocation[J].Biofouling:The Journal of Bioadhesion and Biofil m Research,2009,25(4):325—333.
[25]李静,严涛,曹文浩,等.近海污损生物生态研究进展[J].海洋通报,2010,29(1):113—119.
[26]郑成兴,黄宗国,李传燕,等.闽江口污损生物生态[J].海洋学报,1996,18(3):139—146.
[27]黄宗国,蔡如星.海洋污损生物及其防涂(上册)[M].北京:海洋出版社,1984:1—158.
[28]王建军,黄宗国,郑成兴,等.厦门和东山外来物种沙筛贝的种群动态和结构[J].台湾海峡,1999,18(4):372—377.
[29]严涛,黎祖福,胡煜峰,等.中国沿海无柄蔓足类研究进展[J].生态学报,2012,32(16):5230—5241.
[30]林和山,王建军,郑成兴,等.东山湾污损生物生态研究[J].海洋学报,2012,34(6):160—169.
[31]GB/T 12763.6-2007.海洋调查规范[S].北京:中国标准出版社,2007.
[32]国家海洋局第三海洋研究所.大亚湾海洋生态文集Ⅱ[G].北京:海洋出版社,1990:478—488.
[33]王宝强,薛俊增,庄骅,等.洋山港海域大型污损生物生态特点[J].海洋学报,2012,34(3):155—162.
[34]李传燕,林盛,黄宗国,等.沙埕港污损生物的生态研究[J].台湾海峡,1994,13(2):111—117.
[35]王晓波,龙绍桥,何东海,等.舟山朱家尖海域污损生物的分布特征及其与环境因子的关系[J].海洋学研究,2012,30(1):41—50.
[36]黄宗国,陈丽淑.台湾省两个港湾污损生物初步研究[J].海洋学报,2002,24(6):92—98.
[37]蔡立哲,李复雪.厦门港大型污损生物的垂直分布[J].热带海洋,1986,5(4):10—18.
猜你喜欢
计算机仿真(2022年9期)2022-10-25
食品研究与开发(2022年16期)2022-08-24
军事文摘(2021年22期)2022-01-18
大科技·百科新说(2021年5期)2021-07-27
数学小灵通·3-4年级(2021年6期)2021-07-16
今日农业(2020年14期)2020-08-14
资源节约与环保(2019年11期)2019-01-21
环境保护与循环经济(2017年5期)2018-01-22
延河(下半月)(2015年4期)2015-09-24
延河·绿色文学(2015年4期)2015-06-01
