
水稻单片段代换聚合系的产量比较试验
2015-02-18 05:45
华南农业大学学报 2015年4期
水稻单片段代换聚合系的产量比较试验
朱海涛,刘桂富,曾瑞珍,刘自强,傅雪琳,代资举,王少奎,张桂权
(广东省植物分子育种重点实验室/华南农业大学农学院,广东广州510642)
摘要:【目的】基于水稻单片段代换系聚合而成的品系,是分子设计育种过程中的中间材料,通过评价其优劣,为育种和生产实践提供有益的参考.【方法】以25个聚合系为材料,在2013年早、晚两季采用随机区组试验测验它们与华粳籼74主要农艺性状的差异显著性,并探讨所考察性状的表型和相关的遗传基础.【结果和结论】5个聚合系P05、P08、P14、P15和P21的产量显著地高于华粳籼74,适于早、晚季种植;另2个高产聚合系P07和P12则仅适于晚季种植.所有考察性状的遗传率变动在0. 885 9~0. 993 7之间,其中,普通遗传率为0. 554 7~0. 968 8,互作遗传率为0. 024 9~0. 331 2.着粒密度降低产量,贡献率为15. 5%;而千粒质量增加产量,贡献率达36. 6%.试验结果为这些性状的遗传改良和育种上的间接选择提供了依据.
关键词:单片段代换系;聚合系;农艺性状;产量;水稻;遗传率
朱海涛,刘桂富,曾瑞珍,等.水稻单片段代换聚合系的产量比较试验[J].华南农业大学学报,2015,36(4):43-49.
优先出版网址: http: / /www.cnki.net/kcms/detail/44.1110.s.20150610.1541.003.html
水稻Oryza sativa L.是世界上最重要的粮食作物之一.一方面,随着人口的不断增长,预计到2030年粮食产量需要再增加40%才能满足人们对粮食的需求[1];另一方面,随着生活水平的不断提高,人们对大米品质的要求愈来愈高,因而水稻高产、优质已成为重要的育种目标.矮化育种、杂种优势利用和超级稻育种,使水稻单产持续上升到一个较高的平台[2-4],但近10年来已停滞不前、难有进一步的突破,且稻米品质较差,无法满足人们的需求[5].Peleman等[6]提出了设计育种的新思路,包括定位相关性状的QTL、评价这些位点的等位性变异和开展设计育种,有望突破水稻高产、优质等方面的瓶颈.
广东省植物分子育种重点实验室(本实验室)培育了1 600多份水稻单片段代换系[7-8],用这些材料鉴定了许多重要农艺性状的QTLs及其等位基因[9-13],并广泛开展了分子设计育种[14-15].用带有产量、品质和抗性等优良基因的单片段代换系聚合,获得了一批高产、优质、多抗的多片段聚合品系或品种,该项工作仍在紧张地进行中,新的聚合系不断呈现.然而,由于设计育种的理论和方法尚不够完善,对单个优良基因被聚合后的综合表现尚不甚了解,因此许多聚合系并不尽如人意,有必要经过评比试验挑选出有潜力的聚合系以参加后续的品比试验,供生产上推广应用.
本研究拟对本实验室新近培育的25个聚合系进行品比试验,对照采用单片段代换系的受体亲本华粳籼74(本实验室培育的高产优质多抗籼稻品种).一方面,通过对一些重要农艺性状的遗传和相关分析,探索这些性状的遗传基础,以便为它们的遗传改良和相关选择提供依据;另一方面,通过对比,挑选出综合性状优良的聚合系,为后续的品比试验提供参试材料和依据.
1 材料与方法
1.1材料
选用25份新近培育的聚合系为试验材料,以华粳籼74为对照,进行品比试验.这些聚合系材料来源于受体亲本华粳籼74不同单片段代换系的聚合,有关聚合基因及其对应表现型见表1.
1.2方法
1.2.1田间试验试验在华南农业大学教学试验农场进行,于2013年早、晚两季分别品比.
早季于2月22日播种、3月24日移栽;晚季于7 月19日播种、8月6日移栽.采用3次重复的田间随机区组设计;每个小区长6. 33 m、宽2. 20 m,小区面积14. 96 m2;行株距为20. 00 cm×16. 67 cm,每个小区种植418穴,每穴插1株苗.
1.2.2性状考察田间考察包括如下性状:分糵数,移栽后5周调查每穴分蘖总数(含主茎) ;抽穗期,统计单株从播种至始穗的天数;株高,成熟期测量单株从地面到主穗顶(不连芒)的长度;穗数,有5粒以上实粒的穗数.每小区均调查对角20个单株的目标性状.为考察穗部性状,在每小区的对角处根据穗数分别选择2个单株,共4个单株,从穗颈节之下剪取全部稻穗,晾干后用于室内考种.分别考察以下性状:穗长、实粒数、空粒数、结实率、着粒密度、千粒质量.每小区内的植株全部收割、晒干风净后称产.
1.2.3统计方法按随机区组设计试验资料进行相关的统计分析,方差分析的线性模型为:
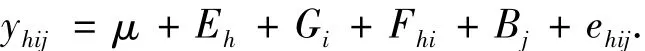
式中,yhij、μ、Eh、Gi、Fhi、ehij分别为表型值、平均值、环境效应、基因型效应、基因型与环境互作效应、区组效应和试验误差,下标h、i、j分别为对应的序号.用最小显著差数法(Least significant difference,LSD 法)测验各聚合系与对照间的差异显著性;普通遗传率估计为:,互作遗传率估计为:,其中遗传方差,互作遗传方差,表型方差,式中的MSG、MSG×E、MSe分别为基因型、基因型与环境互作、误差均方,k,r分别为环境数和区组数;成对性状间的表型相关系数估算为:,遗传相关系数估算为:其中,式中,CMS和COV分别为遗GP传协方差和表型协方差,CMSG、CMSG×E、CMSe分别为基因型、基因型与环境互作、误差协均方.所有分析用统计软件SAS进行.
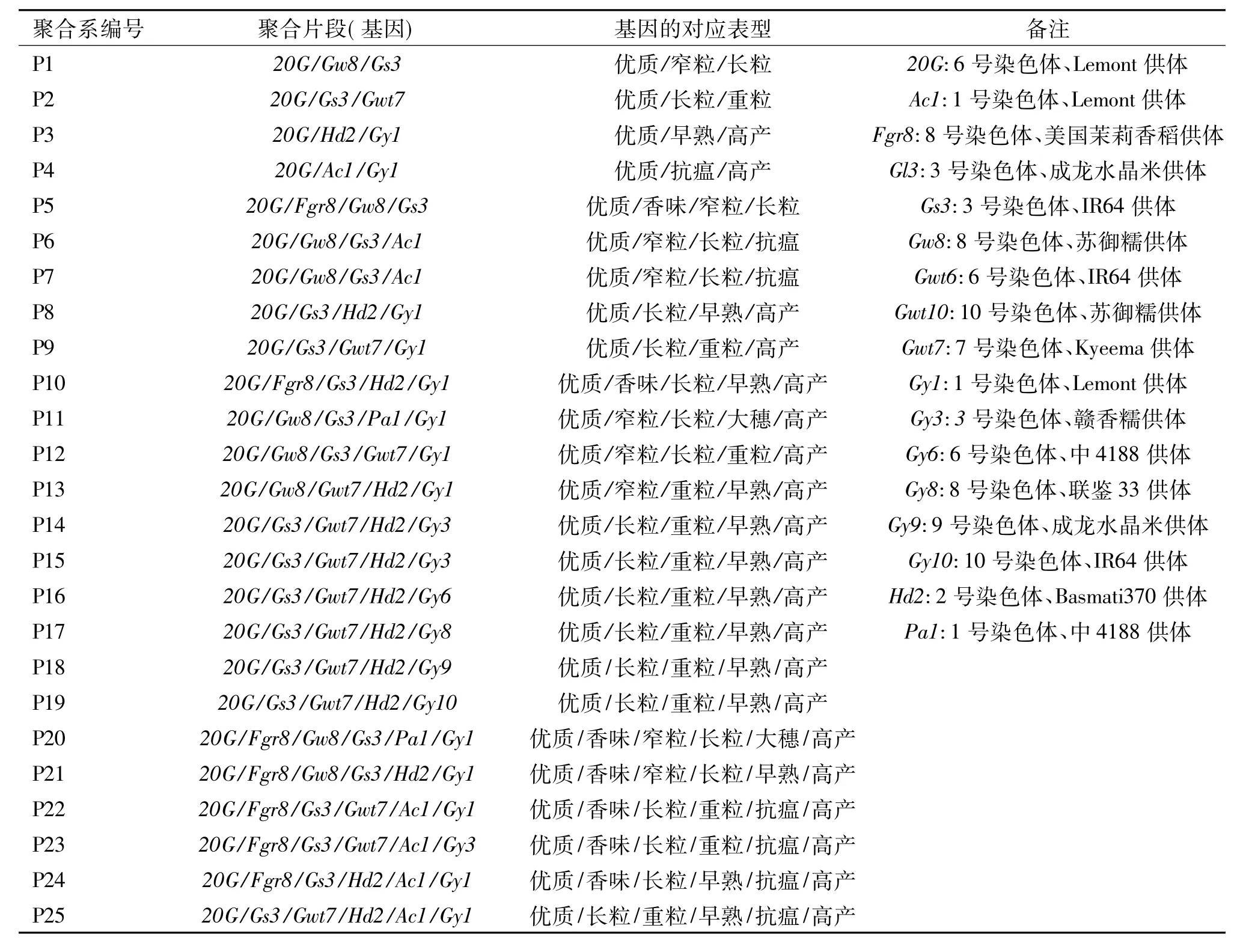
表1 聚合系聚合的优良基因及其对应表型Tab.1 The target genes and their corresponding phenotypes of the pyramiding lines
2 结果与分析
2.1性状的遗传分析
将参试的25个聚合系和华粳籼74共26个材料,按2个环境3次重复的随机区组设计试验资料作方差分析,结果列于表2.表2的结果表明,在整个试验群体中,分蘖数、实粒数、结实率和千粒质量4个性状早、晚季间差异不显著,其余性状则表现出显著的差异.不同的聚合系材料间成穗率和结实率没有显著的差异,但有些聚合系在早、晚季会有不同的结实率;其余性状则表现出显著的差异,且有些聚合系的抽穗期、株高、穗长、着粒密度、千粒质量和产量在早、晚季表现不同.总的来看,4个性状(抽穗期、株高、着粒密度和产量)的变异主要来源于环境、基因型及其互作,穗数主要来源于环境和基因型,千粒质量主要来源于基因型及其与环境的互作,其余性状(分蘖数、成穗率、实粒数和结实率)的表现则主要取决于某一因素(或环境或基因型或基因型与环境互作).
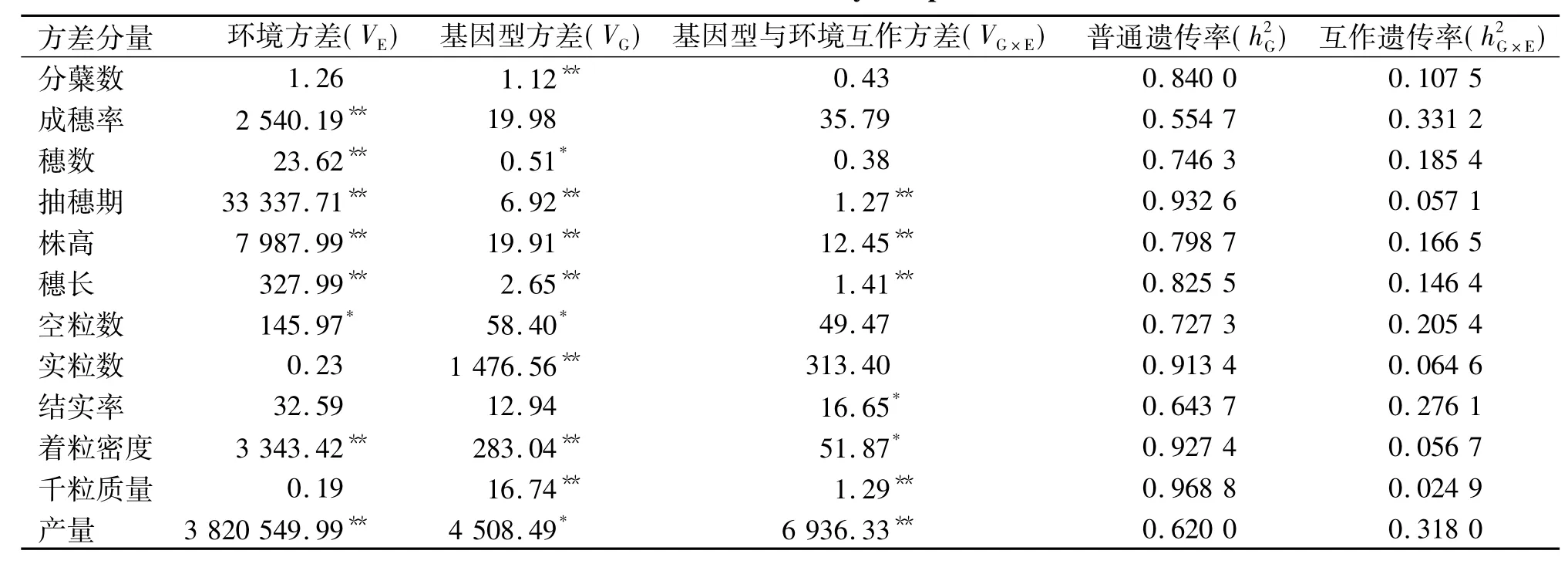
表2 单个性状的方差和遗传率分量1)Tab.2 The estimations of variance and heritability components for each of observed traits
为了说明各遗传因素(基因型及其与环境互作)对性状影响的重要性,普通遗传率和互作遗传率分别被估算(表2).前者是稳定的,适用于所有基因型和环境;后者则随基因型和环境的不同而异,仅适用于特定的基因型和环境.遗传率的高低常作为性状选择的一个参考指标,用以判断性状的变异传递给后代能力的强弱.遗传率高,表明性状的变异主要来源于遗传因素,对这种性状选择有效;遗传率低,则表明性状的变异主要由非遗传因素所引起,对这种性状的选择效果差.本研究所考察性状的广义遗传率变动在0. 885 9~0. 993 7之间,表明性状的变异绝大多数源于不同的基因型,试验误差较小.其中普通遗传率在0. 554 7~0. 968 8之间,是不受环境影响的分量,适用于早、晚季;互作遗传率在0. 024 9~0. 331 2之间,是受到环境影响的分量,仅适用于特定的环境(早季或晚季).
2.2性状间的相关分析
相关系数(r)是指双变量的协方差除以它们各自标准差的乘积,用来度量2个变量间的相关性和密切程度.利用78对(26×3)数据可估得各考察性状间的表型和遗传型相关关系(表3).前者描述的是表型值间的相关,后者则是基因型值间的相关.
结果表明,考察性状间的66个表型相关系数(右上角数据)变动在-0. 938~0. 929之间,其中46 个(占69. 7%)达到显著或极显著水平;分别有37个(占56. 0%)呈正相关,29个呈负相关.进一步将相关系数平方得到决定系数(r2),用以度量相关的密切程度,其值变动在0~0. 880之间,表明某一性状的变异对另一性状的贡献不超过88. 0%.然而,由于表型值受到许多非遗传因素的影响,因此表型相关并不能真实反映2个性状间的关联是缘于基因型还是其他非遗传因素.遗传相关系数反映了2个性状基因型值间的相关.结果表明,在估得的66个遗传相关系数(左下角数据)中,其值在-0. 840~0. 937之间变动,有27个达5%或1%显著水平,36个为正相关,遗传决定系数在0~0. 878之间变动.
产量是重要的育种目标,是多种因素综合影响的结果,与许多性状包括株形和穗部性状等关系密切.本研究表明,产量与分蘖数、实粒数、千粒质量的表型相关不显著,但与其他性状间的相关则达显著或极显著水平,其中与穗长、空粒数呈负相关,其余为正相关;表型决定系数在0~0. 863之间变动.然而,与产量遗传相关显著的则仅有着粒密度和千粒质量2个性状,前者为负相关,决定系数为0. 155,贡献率为15. 5%;后者为正相关,决定系数0. 366,贡献率为36. 6%.

表3性状间表型(右上)和遗传型(左下)相关系数1)Tab.3 The estimations of phenotypic (upper right data) and genotypic (lower left data) correlation coefficients of the evaluated traits
2.3聚合系的评价
将聚合系各性状平均数分别与对照华粳籼74比较,并用LSD法作显著性检验,结果列于表4.表4表明,3个聚合系P05、P18和P22比对照显著提高了分蘖数,仅P05能提高有效穗数;没有检测到有聚合系能显著地改良成穗率、实粒数和株高;共有12个聚合系比对照显著地提早0. 8~3. 7 d抽穗,聚合系P18使主穗增长0. 8 cm,4个聚合系显著地降低单株空粒数6. 4~9. 4粒,聚合系P03提高结实率4. 5%,聚合系P13主穗着粒密度每10 cm增加了6. 4粒,18个聚合系增加千粒质量达1. 0~5. 0 g,5个聚合系P05、P08、P14、P15和P21比对照的产量显著地提高了388. 5~454. 5 kg·hm-2.
2. 4聚合系效应早、晚季的差异
为评价早、晚两季各聚合系的表现,它们各季与对照的差值及其显著性列于表5.表5的结果表明,早、晚季之间一些聚合系对某些性状表现出较大的差异,而对另一些性状则比较稳定.如P05除千粒质量外,其他性状的效应早、晚季间差异明显.早、晚季产量效应间的差异较大,早季没发现有显著改变产量的聚合系,晚季显著增产的聚合系有P05、P07、P08、P12、P14和P21.

表4 聚合系各性状与对照华粳籼74的差值及显著性1)Tab.4 The difference values and significances of the observed traits between the pyramiding lines evaluted and the contrast Huijingxian 74

表5 2013年早、晚两季聚合系各性状与对照华粳籼74的差值及其显著性1)Tab.5 The difference values and significances of the observed traits between pyramiding lines evaluated and the contrast Huijingxian 74 in the early and late seasons in 2013,respectively
3 讨论与结论
自设计育种概念提出以来,分子设计育种实践已成为潮流[16-18].利用分子标记技术可鉴定和分离优良基因(QTL)并将它们聚合到某一优良品种中.本实验室一直致力于分子设计育种实践,用华粳籼74的单片段代换系鉴定和分离了许多重要农艺性状的优良基因(QTL),并按照育种目标将不同的优良基因组装到华粳籼74中,期望华粳籼74有进一步的改良[14-15].本研究的25个试验材料即是在华粳籼74中聚合了不同优良基因的聚合系,但它们大多数的表现却与预期的不一致,许多重要农艺性状,如成穗率、株高、穗长、实粒数和着粒密度等的表现反而与人们的意愿相违背,这也表明了分子聚合育种的难度.5个聚合系P05、P08、P14、P15和P21比华粳籼74的产量有显著的提高,其原因可能是产量构成因素中的千粒质量增加,它们可作为后续品比试验的候选品系.2个高产品系P07 和P12仅适用于晚季.
本试验所考察的性状,如株型性状分蘖数、穗数、株高等,穗部性状穗长、实粒数、千粒质量、产量等,均是水稻重要的农艺性状,分析其在不同品系间性状的差异对于了解水稻的特性及进一步培养理想品种具有重要意义[19].本研究结果表明,一些性状如成穗率、穗数、抽穗期、株高、穗长、空粒数、着粒密度和产量,是环境敏感性的,早、晚季会有不同的表现;而分蘖数、实粒数、结实率和千粒质量早、晚季差异不大.性状的遗传主要取决于基因型及其与环境的互作2个分量.一些性状如分蘖数、穗数、空粒数和实粒数的遗传主要由基因型决定,因而在早、晚季选择均是有效的;另一些性状如抽穗期、株高、穗长、结实率、着粒密度、千粒质量和产量,带有显著的基因型与环境互作方差,因而仅在早季或晚季选择有效.
由于性状的无限性和遗传物质载体的有限性,必然导致遗传连锁现象,察觉、利用或打破连锁是遗传育种研究的重要课题[20].性状间的表型相关系数,反映了性状表型值之间的相关性和密切程度,本研究表明,产量与成穗率、穗数、抽穗期、株高、结实率、着粒密度呈正的表型相关,贡献率在4. 0%~86. 3%;而产量与穗长、空粒数呈负相关,贡献率分别为61. 8%和4. 7%.表型相关系数包含非遗传因素的干扰,而遗传相关系数排除了非遗传因素的干扰,可作为育种间接选择的参考指标.本研究表明,产量与着粒密度和千粒质量有显著的遗传相关性,前者为负,贡献率15. 5%;后者为正,贡献率36. 6%.这2个性状可作为今后育种的参考指标.
参考文献:
[1]KHUSH G S.What it will to feed 50 billion rice consumers in 2030[J].Plant Mol Biol,2005,59(1):1-6.
[2]周开达,马玉清,刘太清,等.杂交水稻亚种间重穗型组合选育:杂交水稻超高产育种的理论与实践[J].四川农业大学学报,1995,13(4):403-407.
[3]袁隆平.杂交水稻超高产育种[J].杂交水稻,1997,12 (6):1-6.
[4]程式华,庄杰云,曹立勇,等.超级杂交稻分子育种研究[J].中国水稻科学,2004,18(5):377-383.
[5]PENG S B,TANG Q Y,ZOU Y B,et al.Current status and challenges of rice production in China[J].Plant Prod Sci,2009,12(1):3-8.
[6]PELEMAN J D,VAN DER VOORT J R.Breeding by design[J].Trends Plant Sci,2003,8(7):330-334
[7]ZHANG G Q,ZENG R Z,ZHANG Z M,et al.The construction of a library of single segment substitution lines in rice (Oryza sativa L.)[J].Rice Genet Newslett,2004,21: 85-87.
[8]XI Z Y,HE F H,ZENG R Z,et al.Development of a wide population of chromosome single segment substitution lines (SSSLs) in the genetic background of an elite cultivar in rice (Oryza sativa L.)[J].Genome,2006,49: 476-484.
[9]杨自凤,朱海涛,刘自强,等.基于单片段代换系的水稻抽穗期QTL上位性研究[J].华南农业大学学报,2014,35 (6) :24-28.
[10]何风华,席章营,曾瑞珍,等.利用单片段代换系鉴定水稻株高及其构成因素的QTL[J].中国水稻科学,2005,19(5):387-392.
[11]LIU G F,ZHU H T,ZHANG G Q,et al.Dynamic analysis of QTLs on tiller number in rice (Oryza sativa L.) with single segment substitution lines[J].Theor Appl Genet,2012,125: 143-153.
[12]WANG S K,WU K,YUAN Q B.Control of grain size,shape and quality by OsSPL16 in rice[J].Nat Genet,2012,44(8):950-955.
[13]YING Y Q,ZHU H T,YE G Y,et al.Detection of QTL on panicle number in rice (Oryza sativa L.) under different densities with single segment substitution lines[J].Euphytica,2014,195(3):355-368.
[14]陈忠平.基于单片段代换系的高产优质水稻设计育种[D].广州:华南农业大学,2010.
[15]梁海福.基于单片段代换系的高产优质水稻设计育种[D].广州:华南农业大学,2011.
[16]薛勇彪,王道文,段子渊,等.分子设计育种研究进展[J].中国科学院院刊,2007,22(6):486-490.
[17]苏岩,钱前,曾大力,等.水稻分子设计育种的现状和展望[J].中国稻米,2010,16(2):5-9.
[18]WAN J M.Perspectives of molecular design breeding in crops[J].Acta Agron Sin,2006,32: 455-462.
[19]张坚勇,肖应辉,万向元,等.水稻品种外观品质性状稳定性分析[J].作物学报,2004,30(6):548-554.
[20]何秀英,陈钊明,廖耀平,等.水稻收获指数遗传及其与主要农艺性状的相关研究[J].作物学报,2006,32(6):911-916.
【责任编辑周志红】
Comparative trial on grain yield for pyramiding lines of single segment substitution lines in rice
ZHU Haitao,LIU Guifu,ZENG Ruizhen,LIU Ziqiang,FU Xuelin,DAI Ziju,WANG Shaokui,ZHANG Guiquan
(Guangdong Key Lab of Plant Molecular Breeding/College of Agriculture,South China Agricultural University,Guangzhou 510642,China)
Abstract:【Objective】Pyramiding lines derived from single segment substitution lines in rice are intermediate materials in the process of molecular design breeding,so it would provide helpful consults for breeding and production practice via evaluation of their phenotypes.【Method】Twenty-five pyramiding lines were applied as experimental materials,and planted in the field with randomized block design in early and late seasons in 2013 respectively to test the differences of important agronomic traits between each of them and Huajingxian 74.The genetic basis of phenotypes and correlations of all observed traits were explored.【Result and conclusion】The grain yields of 5 pyramiding lines P05,P08,P14,P15 and P21 were markedly higher than that of Huajingxian74,which were suitable for planting in early and late seasons.The high-yield lines P07 and P12 were merely suitable for planting in late season.The heritability ratios estimated for all observed traits varied from 0. 885 9 to 0. 993 7,of which the general heritability ratios were 0. 554 7-0. 968 8 and the interaction heritability ratios were 0. 024 9-0. 331 2.Two traits of seed density and thousand seed mass were detected to influence grain yield,with the former decreasing it by the contribution ratio of 15. 5% while the later increasing it by 36. 6%.The results provide a basisbook=44,ebook=176of genetic improvement and indirect selection in breeding practice for these investigated traits.
Key words:single segment substitution line; pyramiding line; agronomic trait; grain yield; rice; heritability ratio
基金项目:NSFC-广东联合基金重点项目(U1031002) ;广东省自然科学基金(S2013010012939和S2013010013184) ;广州市科技计划项目(2014J4100241) ;亚热带农业生物资源保护与利用国家重点实验室开放课题(SKL-CUSAb-2013-07)
作者简介:朱海涛(1978—),男,实验师,硕士,E-mail: zhuht@ scau.edu.cn;通信作者:张桂权(1957—),男,教授,博士,E-mail: gqzhang@ scau.edu.cn
收稿日期:2014-10-12优先出版时间:2015-06-10
文章编号:1001-411X(2015) 04-0043-07
文献标志码:A
中图分类号:S511; S502
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27
当代水产(2021年10期)2021-12-05
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-11-25
今日农业(2020年20期)2020-11-26
文苑(2020年6期)2020-06-22
文苑(2019年22期)2019-12-07
聚氯乙烯(2018年9期)2018-02-18
中国中药杂志(2016年20期)2016-11-19

- 华南农业大学学报的其它文章
- 5-氯甲基水杨醛缩L-酪氨酸锰(II)配合物的合成及生物活性的研究