
冀北辽河源阔叶混交林与油松林土壤呼吸及其影响因子1)
2015-03-10 05:34于舒韩海荣康峰峰程小琴立天宇赵伟红
东北林业大学学报 2015年4期
于舒 韩海荣 康峰峰 程小琴 立天宇 赵伟红
(北京林业大学,北京,100083)
冀北辽河源阔叶混交林与油松林土壤呼吸及其影响因子1)
于舒 韩海荣 康峰峰 程小琴 立天宇 赵伟红
(北京林业大学,北京,100083)
对生长季内阔叶混交林和油松林的土壤呼吸速率进行研究,探讨冀北辽河源地区土壤呼吸与地下5 cm土壤湿度、地下5、10、15 cm土壤温度(T5、T10、T15)、近地面大气温度、土壤微生物量碳、土壤理化性质的关系。结果表明:阔叶混交林和油松林的土壤呼吸速率的季节变化呈明显的单峰曲线,平均值分别为4.28、3.69 μmol·m-2·s-1,与土壤温度的季节变化曲线大致相同,而滞后于大气温度。阔叶混交林、油松林的各温度与土壤呼吸均呈显著的正相关关系(P<0.01);而土壤湿度和各土层的阔叶混交林与土壤呼吸的相关性不显著,但群落间土壤有机碳、w(C)/w(N)、全氮、全磷、土壤微生物量碳均达到显著差异。综合分析,地下5 cm土壤温度为该地区森林土壤呼吸速率季节变化的主要影响因子;而土壤微生物量碳、凋落物种类及土壤理化性质的综合影响可能是引起两种群落的土壤呼吸速率差异的原因。
阔叶混交林;油松林;土壤呼吸;土壤温度;土壤微生物量碳;土壤理化性质
We studied the soil respiration of mixed broad-leaved forest (MBF) andPinustabulaeformisforest (YF) within the growing season, and discussed the relationships between soil respiration rate and soil moisture at 5 cm depth (W), soil temperature at 5 cm, 10 cm, 15 cm depth (T5,T10,T15), near-surface air temperature (Ta), soil microbial biomass carbon (MBC), soil physical and chemical properties in Liaoheyuan Nature Reserve of Northern Hebei. The seasonal variations of YF and MBF’s soil respiration rate represented a single peak curve with the means of 4.28 and 3.69 μmol·m-2·s-1, respectively. It was the same with the seasonal variation of soil temperature curve, but laged behind atmospheric temperature.T5,T10,T15,Taand corresponding soil respiration of MBF, YF showed a significant positive correlation (P<0.01), and the relationship between W, each layer of MBC and soil respiration were not significant. But significance between communities of soil organic carbon,w(C)/w(N), total nitrogen, total phosphorus, soil microbial biomass carbon reached significant levels. By comprehensive analysis,T5was the main factor leading forest soil respiration seasonal variations in this area, and the combined effects of MBC, litter type and soil physical and chemical properties in all probability caused the differences between the soil respirations of two communities.
在整个生态圈中,土壤作为一个巨大的碳库,其碳储量约为1 500 Pg,其中大约73%的碳由森林生态系统维持[1-2]。土壤呼吸作为土壤碳库的主要输出途径和大气CO2的重要来源[3],往往作为土壤生物活性和土壤肥力的指标受到重视[3-6]。研究表明,引起土壤呼吸季节差异的原因主要是土壤温度[5,7-8],土壤温度通过影响土壤理化性质、植被根系和土壤微生物等的活动而影响土壤呼吸。凋落物作为土壤有机质输入的主要来源,是土壤呼吸的物质基础。在同一气候区,凋落物的分解速率主要受凋落物基本性质和凋落物分解者的影响,一般来说,阔叶树与针叶树相比,阔叶林凋落物良好的性质和快的分解速率被认为可增加阔叶林下土壤的养分可利用性[9],进而影响土壤微生物的活动,提高土壤微生物量碳含量[10]。土壤微生物量碳作为活的有机质[11],参与土壤有机质分解、养分循环、污染物的降解和土壤结构形成等诸多土壤生态过程[12-13],对土壤呼吸起到一定的指示作用。
目前,国内关于针叶林与阔叶林土壤理化性质、土壤微生物量碳与土壤呼吸关系的研究较少,而关于冀北地区油松林与阔叶混交林的研究尚未见报道。因此,本研究以冀北辽河源油松(Pinustabulaeformis)天然次生林和山杨(Populusdavidiana)、白桦(Betulaplatyphylla)、辽东栎(Quercusliaotungensis)混交林为研究对象,初步了解辽河源地区土壤呼吸的季节动态特征,分析土壤呼吸与环境因子的关系,并试图揭示该气候区内2种代表性植被群落土壤呼吸的变化规律、差异及其原因。
1 试验地概况
本研究于2013年5—9月在河北省辽河源大窝铺林场进行(118°22′~118°37′E,41°01′~41°21′N)。该地区处于暖温带向寒温带过渡地带,属于半湿润半干旱大陆性季风型山地气候,年均气温7.3 ℃。年平均降水量550 mm。土壤主要是棕壤土和褐土,土层深厚,凋落物层厚1.2~6.0 cm。
油松天然次生林海拔高度为958 m,坡度为25°,林龄为30 a左右,树种组成为油松纯林,林下出现大量油松幼苗,伴生灌木有毛榛子(Corylusmandshurica)、银背风毛菊(Saussureanivea)、胡枝子(Lespedezabicolor)等,草本有细叶苔草(Carexstenophylloides)、黄花蒿(Artemisiaannua)等。平均树高12.24 m,郁闭度0.45。
阔叶混交天然次生林海拔高度为1 067 m,坡度为19.5°,林龄为35 a左右,组成树种以白桦、山杨、辽东栎为主,混有糠椴(Tiliamandshurica)和一些槭树科树种,灌木层有蒙椴(Tiliamongolica)、土庄绣线菊(Spiraeapubescens)等。平均树高11.26 m,郁闭度0.65。
2 研究方法
2.1 样地设置
2013年4月,在油松天然次生林、阔叶混交天然次生林内各设置3块30 m×20 m的固定样地,每个样地均匀选取3个1 m×1 m的小样方,每个样方内设置一个直径为20 cm,高10 cm的PVC土壤环,PVC土壤环露出地面3.5 cm左右。
2.2 土壤呼吸速率及土壤温湿度、大气温度的测定
在5—9月,每月的中下旬选择2 d晴好天气,采用Li-8100土壤碳通量测定仪测定两种群落土壤呼吸速率,每种林型测定1 d,测定时间在08:00—18:00,每个环重复测定3次。5、10、15 cm土壤温度(T5、T10、T15),5 cm土壤湿度由测量系统自带的温度探针同步测得;近地面大气温度(Ta)由测量系统同步测定。
2.3 土壤微生物碳量的测定
在5—9月,每月土壤呼吸测定后,在土壤呼吸观测点附近用钻土芯法取土壤样品。每个样地内各钻取0~5、10~15、15~20 cm深度土壤样品一份,两种群落共计18份。样品立即装入保温箱内带回实验室置于4 ℃下贮存,采用氯仿熏蒸浸提法测定[14]0~5、10~15、15~20 cm深度土壤微生物量碳(MBC5、MBC10、MBC15)。
2.4 土壤理化性质的测定
在7月采集土样并测定。土壤密度采用环刀法测定;土壤有机质质量分数测定采用重铬酸钾氧化—外加热法;土壤全N质量分数测定采用浓硫酸—高氯酸消煮—凯氏定氮法;全P质量分数测定用浓硫—高氯酸消煮—钼锑抗比色法;全K质量分数测定采用氢氧化钠熔融—火焰光度计法。
2.5 数据处理
采用Excel2013进行数据整理。采用Sigmaplot12.5绘图。采用SPSS18.0软件检验两种群落土壤呼吸及其环境因子的显著性差异,分析土壤呼吸与环境因子的相关性。由于土壤温度与土壤呼吸有一定的指数关系,因此采用指数模型模拟二者的相关性[15]:
Rs=aebTa,
(1)
Rs=aebT。
(2)
式中:Rs表示土壤呼吸速率,Ta表示近地面大气温度,T表示土壤温度,a、b为待定参数。
3 结果与分析
3.1 阔叶混交天然次生林与油松天然次生林的土壤呼吸
两种群落土壤呼吸速率的季节变化呈明显的单峰曲线,表现为5、9月份最低,6—8月最高,与土壤温度、土壤湿度的季节变化大体一致,而滞后于空气温度。从6月开始,气温开始升高,降水开始增多,阔叶混交天然次生林的土壤温度、土壤湿度达到最大值,根系呼吸、土壤微生物活动旺盛,凋落物分解作用增强,土壤呼吸速率也达到最大值(6.22 μmol·m-2·s-1),到8月,油松林的土壤温度、土壤湿度也达到最大值,相应的凋落物呼吸(凋落物对土壤呼吸的贡献)和土壤呼吸速率均达到最大值(5.86 μmol·m-2·s-1)(表1—表3)。
表1 两种群落土壤及凋落物呼吸速率的季节变化

μmol·m-2·s-1
注:表中数值为平均值±标准差。
两种群落的土壤呼吸无显著差异,阔叶混交天然次生林(4.28 μmol·m-2·s-1)>油松天然次生林(3.69 μmol·m-2·s-1),与土壤微生物碳量(P<0.05)、全氮(P<0.05)、全磷(P<0.05)、全钾的大小顺序一致,而与有机碳(P<0.05)、W(C)/W(N)(P<0.05)的大小顺序相反,水热因子均无显著差异。在6—8月,阔叶混交天然次生林良好的水热条件、较高的土壤养分、土壤有机质和较低的W(C)/W(N)可能更利于微生物的分解消耗有机碳[16],导致其有机碳较小,而土壤微生物碳量较大。随着深度的增加,土壤温度、土壤湿度和土壤微生物碳量均明显下降(表3—表5)。说明不同群落土壤呼吸速率除与温湿度因子有关,还与土壤理化性质和土壤微生物碳量等环境因子有关。

表2 阔叶混交天然次生林水热因子的季节变化
注:表中数值为平均值±标准差。
3.2 土壤呼吸与环境因子的关系
3.2.1 温湿度
阔叶混交天然次生林和油松天然次生林的近地面大气温度、土壤温度与土壤呼吸速率均呈极显著的指数相关(P<0.01)(表6、表7),土壤呼吸与土壤温度的相关性高于近地面大气温度,其中与T5的相关性最高,T5可分别解释72.4%和69.2%的土壤呼吸速率变化;而近地面大气温度则可以分别解释43.1%和31.0%的土壤呼吸速率变化。而两种群落土壤呼吸随土壤湿度增加而增大,但二者之间的相关关系不显著(P>0.05)(表8)。

表3 油松天然次生林水热因子的季节变化
注:表中数值为平均值±标准差。

表4 两种群落土壤微生物量碳的季节变化
注:表中数值为平均值±标准差。

表5 两种群落土壤理化性质及土壤微生物碳量月均值
注:MBC5、MBC10、MBC15分别表示0~5、10~15、15~20 cm土壤微生物量碳;不同字母表示两种林型之间差异显著(P<0.05)。
从两种群落的土壤呼吸速率比较看,土壤温度、土壤湿度对土壤呼吸速率的影响较复杂。在7月,油松天然次生林的土壤温湿度大于阔叶混交天然次生林,但土壤呼吸却小于油松天然次生林。该结果表明,土壤温度、土壤湿度并不是导致两种群落土壤呼吸速率差异的主要原因,除此之外,土壤呼吸还与其他环境因素有关。
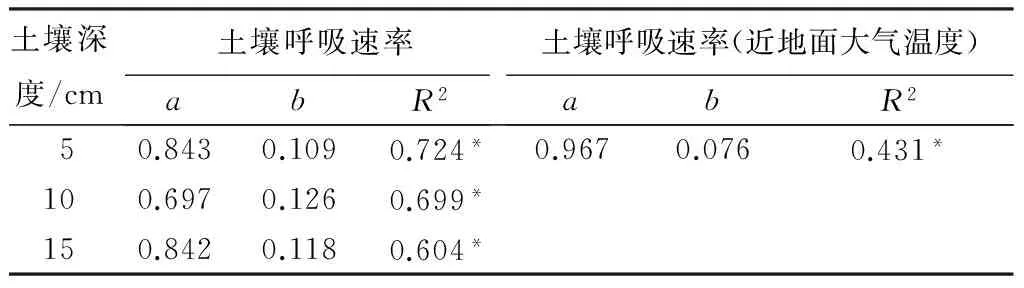
表6 阔叶混交天然次生林土壤呼吸与温度的关系
注:a、b为待定参数;*表示差异极显著(P<0.01)。

表7 油松天然次生林土壤呼吸与温度的关系
注:a、b为待定参数;*表示差异极显著(P<0.01)。

表8 土壤呼吸与环境因子的相关关系
注:W表示土壤湿度;MBC5、MBC10、MBC15分别表示0~5、10~15、15~20 cm土壤微生物量碳。
3.2.2 土壤微生物碳量
随着凋落物的积累以及降水等淋溶作用使凋落物释放的养分和微生物活性逐渐增加,两种群落的土壤微生物碳量从5月开始上升,在8月达到最大值,从9月份开始下降;土壤呼吸不仅包括微生物呼吸,还包括根系呼吸、凋落物呼吸等,其季节变化受土壤温度的影响较明显。因此,土壤微生物碳量与土壤呼吸作用相关但并不完全同步。本研究表明,阔叶混交天然次生林的土壤微生物碳量与土壤呼吸呈负相关,且随着土壤深度的增加,相关性降低;油松天然次生林的土壤微生物碳量与土壤呼吸呈正相关,但与10~15 cm层的土壤微生物碳量相关性最好,但两种群落二者之间的回归关系均未达到显著水平。
4 结论与讨论
两种群落的土壤呼吸速率与土壤温度、近地面大气温度均呈现明显的季节变化(P<0.01),二者之间呈极显著的相关关系(P<0.01),土壤温度对土壤呼吸的解释能力明显高于近地面大气温度,分别可以解释土壤呼吸季节变化的60.4%~72.4%和60.9%~69.2%。说明土壤温度是影响两种群落土壤呼吸季节变化的重要因子,其中T5与土壤呼吸的相关性最好。这与周存宇等[17]、史广松等[18]、王旭等[19]关于森林群落土壤呼吸的研究相同,而与陈全胜等[20]、彭家中等[21]、付刚等[22]关于草原群落或高寒地区土壤呼吸的研究不同。由于T5较高、土壤微生物碳量较大,反映此层的土壤微生物活性和数量较高,加上较好的水热条件有利于有机质的分解和养分释放循环,导致土壤呼吸速率与此层的土壤温度相关性最好。
土壤湿度是影响两种群落土壤呼吸速率的另一个重要因子。很多野外试验测量的结果表明土壤湿度与土壤呼吸速率呈不显著的正相关关系,只有在最高和最低的情况下才会抑制土壤CO2通量[23]。本文研究结果与之一致,两种群落土壤呼吸速率与土壤湿度的关系均不显著,但当土壤温度不成为限制因子的情况下,土壤湿度并没有抑制土壤呼吸,反而使土壤呼吸速率增加。可见,不同群落的土壤呼吸对于土壤湿度范围的反应有所差别。
两种群落的土壤呼吸比较而言,阔叶混交天然次生林明显大于油松天然次生林,但油松天然次生林的土壤温度、土壤湿度略大于阔叶混交天然次生林。因此,土壤温度、土壤湿度并不是导致两种群落土壤呼吸速率差异的主要原因。
从整个测定期间来看,阔叶混交天然次生林的平均土壤呼吸速率明显大于油松天然次生林,但两种群落的微生境因子(近地面大气温度、土壤温度、土壤湿度)并无明显差异(P>0.05)。在水热条件相对稳定的情况下,土壤有机质质量分数是决定土壤呼吸的重要因子[24-25],杨金艳等[26]研究发现土壤呼吸年通量与A0和A层土壤有机质呈显著的正相关关系。在生长旺盛期,阔叶混交天然次生林较高的土壤微生物碳量以及合适的土壤温度、土壤湿度,使土壤微生物开始加速分解土壤有机碳,导致土壤总有机碳含量降低;同时阔叶林的凋落物质量较高,容易分解淋溶转化成可溶性有机碳,为微生物的异养呼吸提供大量有效的碳源[27],加上较低的土壤W(C)/W(N)、较高的土壤养分从而使两种林分的土壤呼吸速率产生差异。由此可见,凋落物质量、土壤理化性质及土壤微生物碳量互相影响形成正反馈机制,最终导致土壤呼吸速率的变化。从季节动态来看,阔叶混交天然次生林的土壤呼吸速率与土壤微生物碳量呈负相关,且随着土层的加深相关性降低,因为阔叶混交天然次生林的O层较薄,而A层较厚,导致0~5 cm的土壤大部分由A层组成,A层的生物与微生物比较活跃,同时植物根系尤其是生理活动最旺盛的植物细根集中分布在这个层次[28]。因此,这一层的土壤微生物碳量与土壤呼吸的相关性较高。同理,油松天然次生林的O层较厚,导致10~15 cm深度土壤土壤微生物碳量与土壤呼吸的相关性较好。但两种群落二者之间的相关性均未达到显著水平,说明土壤微生物碳量可以指示土壤呼吸的大小,但并不能指示土壤呼吸的季节变化。另外,由于不同研究地区、不同森林类型的土壤有机质种类和组成的复杂性,引起土壤养分循环、土壤微生物数量和活性、微生物可利用的底物质量的变异[29],导致土壤呼吸与土壤微生物碳量的反馈关系仍有很多的不确定性。
[1] Post W M, Emanuel W R. Soil carbon pools and world life zones[J]. Nature,1982,298:156-159.
[2] Dixon R K, Brown S, Houghton R A, et al. Carbon pools and flux of global forest ecosystem[J]. Science,1994,263:185-190.
[3] Neilson J W, Pepper I L. Soil respiration as an index of soil aeration[J]. Am J Soil Sci Soc,1990,54:428-432.
[4] Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J]. Tellus,1992,44(2):81-99.
[5] 刘绍辉,方精云.土壤呼吸的影响因素及全球尺度下温度的影响[J].生态学报,1997,17(5):19-26.
[6] Oyonarte C, Rey A, Raimundo J, et al. The use of soil respiration as an ecological indicator in arid ecosystems of the SE of Spain: Spatial variability and controlling factors[J]. Ecological Indicators,2012,14(1):40-49.
[7] 刘春霞,王玉杰,王云琦,等.重庆缙云山3种林型土壤呼吸及其影响因子[J].土壤通报,2013,44(3):587-593.
[8] Hartemink A E, Janssen B H, Buresh R J, et al. Soil nitrate and water dynamics in sesbania fallows, weed fallows and maize[J]. Soil Science Society of America Journal,1996,60(2):568-574.
[9] 王相娥,薛立,谢腾芳.凋落物分解研究综述[J].土壤通报,2009,40(6):1473-1478.
[10] 刘纯,刘延坤,金光泽.小兴安岭6种森林类型土壤微生物量的季节变化特征[J].生态学报,2014,34(2):451-459.
[11] Lavelle P. Ecological challenges for soil science[J]. Soil Science,2000,165(1):73-86.
[12] Wardle D A. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J]. Biological Reviews,1992,67(3):321-358.
[13] Piotrowska A, Dugosz J. Spatio-temporal variability of microbial biomass content and activities related to some physicochemical properties of Luvisols[J]. Geoderma,2012,173:199-208.
[14] Luo Y Q, Wan S Q, Hui D F, et al. Acclimatization of soil respiration to warming in a tall grass prairie[J]. Nature,2001,413:622-625.
[15] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:228-233.
[16] Paul E A, Clark F E. Soil Microbiology and Biochemistry[M]. San Diego: Academic Press,1989.
[17] 周存宇,周国逸,王迎红,等.鼎湖山针阔叶混交林土壤呼吸的研究[J].北京林业大学学报,2005,27(4):23-27.
[18] 史广松,刘艳红,康峰峰.暖温带森林土壤呼吸随林分类型及其微生境因子的变异规律[J].江西农业大学学报,2009,31(3):408-436.
[19] 王旭,周广胜,蒋延玲,等.山杨白桦混交次生林与原始阔叶红松林土壤呼吸作用比较[J].植物生态学报,2007,31(3):348-354.
[20] 陈全胜,李凌浩,韩兴国,等.水热条件对锡林河流域典型草原退化群落土壤呼吸的影响[J].植物生态学报,2003,27(2):202-209.
[21] 彭家中,常宗强,冯起.温度和土壤水分对祁连山青海云杉林土壤呼吸的影响[J].干旱区资源与环境,2008,22(3):165-169.
[22] 付刚,周宇庭,沈振西,等.不同海拔高寒放牧草甸的生态系统呼吸与环境因子的关系[J].生态环境学报,2010,19(12):2789-2794.
[23] Xu L K, Baldocchi D D, Tang J W. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature[J]. Global Biogeochemical Cycle,2004,18(4):963-975.
[24] Fang C, Moncrieff J B. A model for soil CO2production and transport1: Model development[J]. Agricultural and Forest Meteorology,1999,95(4):225-236.
[25] Priess J A, Koning G H, Veldkam p A. Assessment of interactions between land use change and carbon and nutrient fluxes in Ecuador[J]. Agriculture Ecosystem Environment,2001,85:269-279.
[26] 杨金艳,王传宽.东北东部森林生态系统土壤碳贮量和碳通量[J].生态学报,2005,25(11):2875-2882.
[27] Huang Z Q, Wan X H, He Z M, et al. Soil microbial biomass, community composition and soil nitrogen cycling in relation to tree species in subtropical China[J]. Soil Biology and Biochemistry,2013,62:68-75.
[28] 李凌浩,林鹏,邢雪荣.武夷山甜槠林细根生物量和生长量研究[J].应用生态学报,1998,9(4):337-340.
[29] Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change[J]. Nature,2006,440:165-173.
Soil Respiration and Impact Factors ofPinustabulaeformisForest and Mixed Broad-leaved Forest in Liaoheyuan Nature Reserve of Northern Hebei
Yu Shu, Han Hairong, Kang Fengfeng, Cheng Xiaoqin, Li Tianyu, Zhao Weihong(Beijing Forestry University, Beijing 100083, P. R. China)/Journal of Northeast Forestry University,2015,43(4):52-56.
PinustabulaeformisForest; Mixed Broad-leaved Forest; Soil respiration; Soil temperature; Soil microbial biomass carbon; Soil physical and chemical properties
于舒,女,1989年7月生,北京林业大学林学院,硕士研究生。E-mail:ys2064@yeah.net。
韩海荣,北京林业大学林学院,教授。E-mail:hanhr@bjfu.edu.cn。
2014年12月5日。
S718.5
1) 国家林业局林业公益性行业科研专项(201204101)。
责任编辑:潘 华。
猜你喜欢
北京林业大学学报(社会科学版)(2022年4期)2023-01-04
建筑与预算(2022年6期)2022-07-08
天津农林科技(2020年3期)2020-08-13
电子制作(2019年15期)2019-08-27
现代农业研究(2017年11期)2018-01-12
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
现代园艺(2017年20期)2017-02-03
前卫文学(2016年3期)2016-07-01
高原山地气象研究(2016年4期)2016-02-28
