
End-Triassic nonmarine biotic events
2015-03-21 05:49SpencerLucasLawrenceTanner
Journal of Palaeogeography 2015年4期
Spencer G. Lucas *, Lawrence H. Tanner
a New Mexico Museum of Natural History, 1801 Mountain Road NW, Albuquerque, New Mexico 87104-1375 USA
b Department of Biological Sciences, Le Moyne College, 1419 Salt Springs Road, Syracuse,NY 13214 USA
1 Introduction
The biodiversity crisis at the end of the Triassic (Triassic-Jurassic boundary: TJB) has long been identified as one of the “big five” mass extinctions of the Phanerozoic(Sepkoski, 1982). However, during the last decade, we have questioned this identification of a severe and sudden biotic decline at the TJB (Hallam, 2002; Lucas and Tanner, 2004,2008; also see Tanneret al., 2004). Instead, our thesis is that the Late Triassic was a prolonged (approximately 30-million-year-long) interval of elevated extinction rates and low origination rates that manifested themselves in a series of discrete extinctions during Carnian, Norian and Rhaetian time. Most of these extinctions took place in the marine realm, particularly affecting radiolarians, conodonts, bivalves, ammonoids and reef-building organisms (Lucas and Tanner, 2008). On land, the case for a Late Triassic mass extinction is much more tenuous and has largely focused on tetrapod vertebrates (amphibians and reptiles), though some workers advocate a sudden end-Triassic extinction of land plants as well. Here, we focus on the Late Triassic record of land plants and terrestrial tetrapods to demonstrate that they do not document a sudden or a severe end-Triassic extinction. From a palaeogeographical point of view, the nonmarine fossil record identifies some local extinctions across the TJB, but no global mass extinction.
2 Late Triassic-Early Jurassic timescale
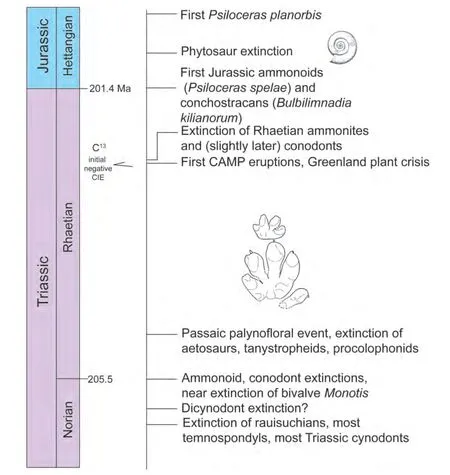
We use the standard global chronostratigraphic timescale for the Late Triassic-Early Jurassic compiled by Ogg(2012a, 2012b), but modified with regard to some recent data and analyses of its radioisotopic calibration (Lucaset al., 2012; Ogget al., 2014; Wotzlawet al., 2014) (Fig. 1).The base of one of the three Late Triassic stages (Carnian) has an agreed on GSSP (global stratotype section and point), and the base of the Norian stage will be defined by a conodont datum that is close to the traditional ammonoidbased boundary (Baliniet al., 2010; Krystynet al., 2007;Lucas, 2010a, 2013b; Orchard, 2010, 2013, 2014).
The favored definition of the Rhaetian base has as its primary signal the LO (lowest occurrence) of the conodontMisikella posthernsteini. At present, the only fully published GSSP candidate for the Rhaetian base is the Steinbergkogel section near Hallstatt, Austria (Krystynet al.,2007). However, Giordanoet al.(2010) concluded that the LO ofMisikella posthernsteiniis actually younger at Steinbergkogel than it is in the section they studied in the Lagronegro basin in southern Italy, so the Italian section may become the GSSP location. The GSSP defined for the TJB (base of Jurassic System = base of Hettangian Stage)is correlated by the lowest occurrence of the ammonoidPsiloceras spelae(Hillebrandtet al., 2013). The best numerical calibration places this boundary close to 201.4 Ma(Schoeneet al., 2010; Fig. 1).
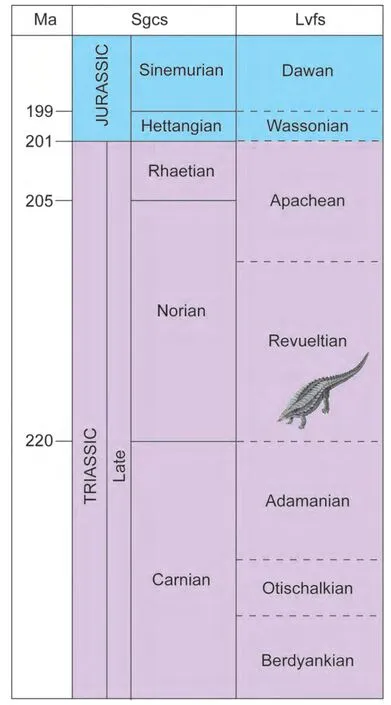
Figure 1 Correlation of the Late Triassic-earliest Jurassic standard global chronostratigraphic scale (sgcs) to tetrapod biochronology based on land-vertebrate faunachrons (lvfs). See text for sources.
Correlation of nonmarine and marine biochronology in the Late Triassic-Early Jurassic remains imprecise. Here,we primarily rely on the correlations based on tetrapod biostratigraphy and biochronology (land-vertebrate faunachrons) advocated by Lucas (1998, 2008, 2010b), Lucaset al.(2007a, 2012) and Lucas and Tanner (2007b) (Fig. 1).Many Late Triassic megafossil plant localities can be directly related to this tetrapod-based biochronology (e.g.,Lucas, 2013a). Palynostratigraphic correlations in much of this interval are discussed in a comprehensive way by Cirilli(2010) and Kürschner and Herngreen (2010). Conchostracan biostratigraphy for the Late Triassic-Early Jurassic also can be tied directly to the tetrapod biostratigraphy and biochronology (Kozur and Weems, 2005, 2007, 2010; Lucaset al., 2011, 2012; Weems and Lucas, 2015).
An extremely important point is that correlation of Late Triassic nonmarine biochronology to the standard global chronostratigraphic scale is achieved primarily by correlation to the Germanic basin section, where marine intercalations and other data provide a reasonably precise cross-correlation to the marine Upper Triassic sections in Austria and Italy (Kozur and Bachman, 2005, 2008; Kozur and Weems, 2010; Lucas, 2010a, 2010b). For the Lower Jurassic, nonmarine-marine correlation tiepoints exist (e.g.,Lucas, 2008), but are constrained in much less detail than is the Upper Triassic record.
Magnetostratigraphy also provides some service to correlation of nonmarine and marine strata across the TJB,but not the precision always needed. The Rhaetian-early Hettangian interval is a time of dominantly normal polarity, and each section studied reveals from two to four short intervals of reversed polarity in the dominantly normal Rhaetian-Hettangian multichron (e.g., Hounslow and Muttoni, 2010; Hounslowet al., 2004; Lucas and Tanner,2007b; Lucaset al., 2011; Marzoliet al., 2004). Correlation of these short reversals to each other is not straightforward and forces a reliance on biostratigraphic or chemostratigraphic inferences to provide precise correlations within the Rhaetian-Hettangian multichron (for example, see Lucas and Tanner, 2007b, Fig. 7).
Incorrect placement of the TJB boundary in the Newark Supergroup long confounded nonmarine-marine correlations and gave the impression of a significant extinction of plants and tetrapods at the TJB (see discussion below).However, well supported and now well accepted correlation of the TJB in the Newark section (Cirilliet al., 2009; Kozur and Weems, 2005, 2007, 2010; Lucas and Tanner, 2007b;Olsenet al., 2011, 2015; Whitesideet al., 2010) moves the boundary upward stratigraphically to a level well removed from any evidence of a sudden biotic turnover.
Correlation of nonmarine strata to the now defined TJB in marine strata is achieved by some workers using either palynostratigraphy or carbon isotope stratigraphy. However, palynostratigraphy does not unambiguously identify the boundary in many sections (Bonis and Kürschner, 2012;Boniset al., 2009; Pieńkowskiet al., 2011). And, many nonmarine carbon isotope records rely on too few data points to convincingly identify the carbon-isotope excursions recognized in marine sections across the TJB or produce records with such high noise levels as to defy correlation (e.g., McEwainet al., 1999). Recent examples of such problematic carbon-isotope-based correlations of the TJB in nonmarine strata include Gómezet al.(2007) in Spain,Whitesideet al.(2010) in the USA, Pieńkowskiet al.(2011,2014) in Poland and Hesselboet al.’s(2002) carbon-isotope curve in East Greenland, discussed below.
Nevertheless, such considerations do not deter some workers from precise placement of the TJB in nonmarine sections where few if any data exist to support precise placement. Thus, for example, Shaet al.(2011, 2015) recently placed the TJB in nonmarine strata of the Junggur Basin in northwestern China using palynomorphs and nonmarine bivalves that cannot be unambiguously correlated with precision to any other TJB section.
More problematic are nonmarine sections where the TJB can only be approximated, and that approximation separates very different nonmarine fossil assemblages that differ in age by many millions of years. A good example of this is provided by the study of the tetrapod record in the Gondwana basins of India by Bandyopadhyay and Sengupta(2006). This record has an obviously Late Triassic tetrapod assemblage (with phytosaurs, aetosaurs,etc.) of likely Norian age in the lower Dharmaram Formation, overlain hundreds of meters higher by the clearly Jurassic (likely Sinemurian), dinosaur-dominated tetrapod assemblage of the Kota Formation. All that intervenes are two undescribed tetrapod taxa — a sphenodontian and a plateosaurid dinosaur — from the upper Dharmaram Formation. Lucas and Huber (2003) suggested these fossils are likely Late Triassic in age, but Bandyopadhyay and Sengupta (2006) assigned them a Hettangian age. They then claimed the Gondwana basin record shows a substantial turnover in tetrapods at the TJB, even though the two assemblages in India that are comparable across that boundary are likely Norian and Sinemurian in age, and thus separated by at least 6 million years, invalidating their conclusion.
Another example comes from the work of Pieńkowskiet al.(2011, 2014) in Poland, where late Norian tetrapod body fossils are compared to Hettangian(?) footprint assemblages and show a change from diverse, dicynodont- and archosaur-dominated faunas to dinosaur-dominated footprint faunas. Dinosaur-dominated footprint faunas, however, are also known from the Late Triassic (e.g., Lucas, 2007), so the comparison of the Polish Norian bone and Hettangian(?)footprint records does not necessarily reflect biotic turnover across the TJB.
3 TJB terrestrial record
It is fair to say that only seven places on Earth have a record of non-marine TJB fossil plants and tetrapods of sufficient extent, diversity and stratigraphic density to be relevant to evaluating a TJB extinction: (1) Chinle-Glen Canyon Group succession of the American Southwest; (2)Newark Supergroup strata of the eastern seaboard of North America from Nova Scotia to the Carolinas, and including rift basin deposits of Morocco in North Africa; (3) Jameson Land in eastern Greenland; (4) European records inside and outside of the Germanic basin, particularly in Germany, the United Kingdom and Italy; (6) northwestern Argentina; and(7) the Karoo basin of South Africa (Fig. 2).
Today, and (based on actualism) during the TJB interval, much terrestrial diversity is accounted for by monerans and arthropods. Yet, the monerans have virtually no fossil record, and the record of arthropods across the TJB is essentially nonexistent for many groups. Arthropod groups that have relatively good TJB fossil records, such as ostracods and conchostracans, show no significant extinction at the TJB (e.g., Kozur and Weems, 2010). Insects also have a fossil record that shows no evidence of a diversity crash across the TJB (Grimaldi and Engel, 2005; Labandeira and Sepkoski, 1993). Indeed, Grimaldi and Engel (2005, p. 73)concluded, “there seems to have been little differentiation between insect faunas of the Late Triassic and Early Jurassic.”
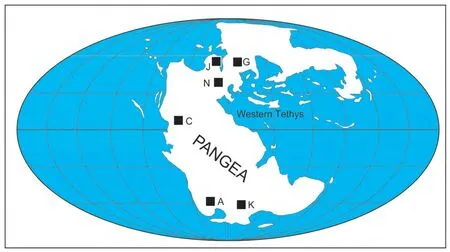
Figure 2 Principal localities that are referred to in the text where the nonmarine Triassic-Jurassic boundary interval has a dense fossil record: A = Argentina, C= Chinle basin, American Southwest, G = Germanic basin, J = Jameson Land, East Greenland, K=Karoo basin,South Africa and N=Newark Supergroup basins.
Plant diversity across the TJB is based on a much better fossil record, both in the form of megafossil plants and their microfossil “proxies,” the palynomorphs (sporomorphs). The record of terrestrial fossil vertebrates across the TJB is comparable in quality to the plant record, and it shows no evident extinction of freshwater fishes (McCune and Schaeffer, 1986; Milneret al., 2006). Supposed TJB tetrapod extinctions focus on three principal groups: temnospondyl amphibians, some archosaur taxa and some synapsid taxa. These records have been evaluated using both body fossils (bones and teeth) and trace fossils (footprints).
4 Land plants
4.1 Overview
An extensive literature does not identify a major extinction of land plants at the TJB (e.g., Ash, 1986; Brugman,1983; Burgoyneet al., 2005; Cascales-Miñana and Cleal,2012; Cleal, 1993a, 1993b; Edwards, 1993; Fisher and Dunay, 1981; Galliet al., 2005; Hallam, 2002; Kelber, 1998;Knoll, 1984; Kuerschneret al., 2007; Lucas and Tanner,2004, 2007b, 2008; McElwain and Punyasena, 2007; Niklaset al., 1983; Orbell, 1973; Pedersen and Lund, 1980; Ruckwiedet al., 2008; Schuurman, 1979; Tanneret al., 2004;Traverse, 1988). Most reviews identify only the extinction of peltasperms, a clade of seed ferns, at the TJB. Global compilations at the species and family levels show no substantial plant extinction at the TJB (Cleal, 1993a, 1993b;Edwards, 1993; Knoll, 1984; Niklaset al., 1983). A recent analysis by Cascales-Miñana and Cleal (2012: 76-77) concluded that “the changes observed in the plant fossil record in the Late Triassic Series are mainly reflecting an ecological reorganization of the terrestrial habitats weeding-out some of the families of less adaptable plants that had filled the newly available niches during Triassic times, rather than a global ecological crisis on the scale of that seen at the end of Permian times.”
Some data do reveal local turnover of paleofloras across the TJB. A good example is a study of Swedish benettitaleans that documented restriction of two species to the Rhaetian followed by five species restricted to the Hettangian. However, this is not considered by its authors, Pott and McLoughlin (2009: 117), as anything more than a “moderate taxonomic turnover,” and it is a palaeogeographically local event.
Contrary to prevailing thought, McElwain and collaborators (Baconet al., 2013; Belcheret al., 2010; Manderet al., 2010, 2012; McElwain and Punyasena, 2007; McElwainet al., 1999, 2007, 2009) claim significant changes in the megaflora at the TJB in East Greenland (Fig. 3). According to these workers, these encompass a sudden change from high diversity Late Triassic plant communities to lower diversity and less taxonomically even Early Jurassic communities. This is the only location where a case has been made for a major turnover in the megaflora at the TJB,but we question whether the data support recognition of a megafloral crisis in East Greenland of more than local palaeogeographic significance.
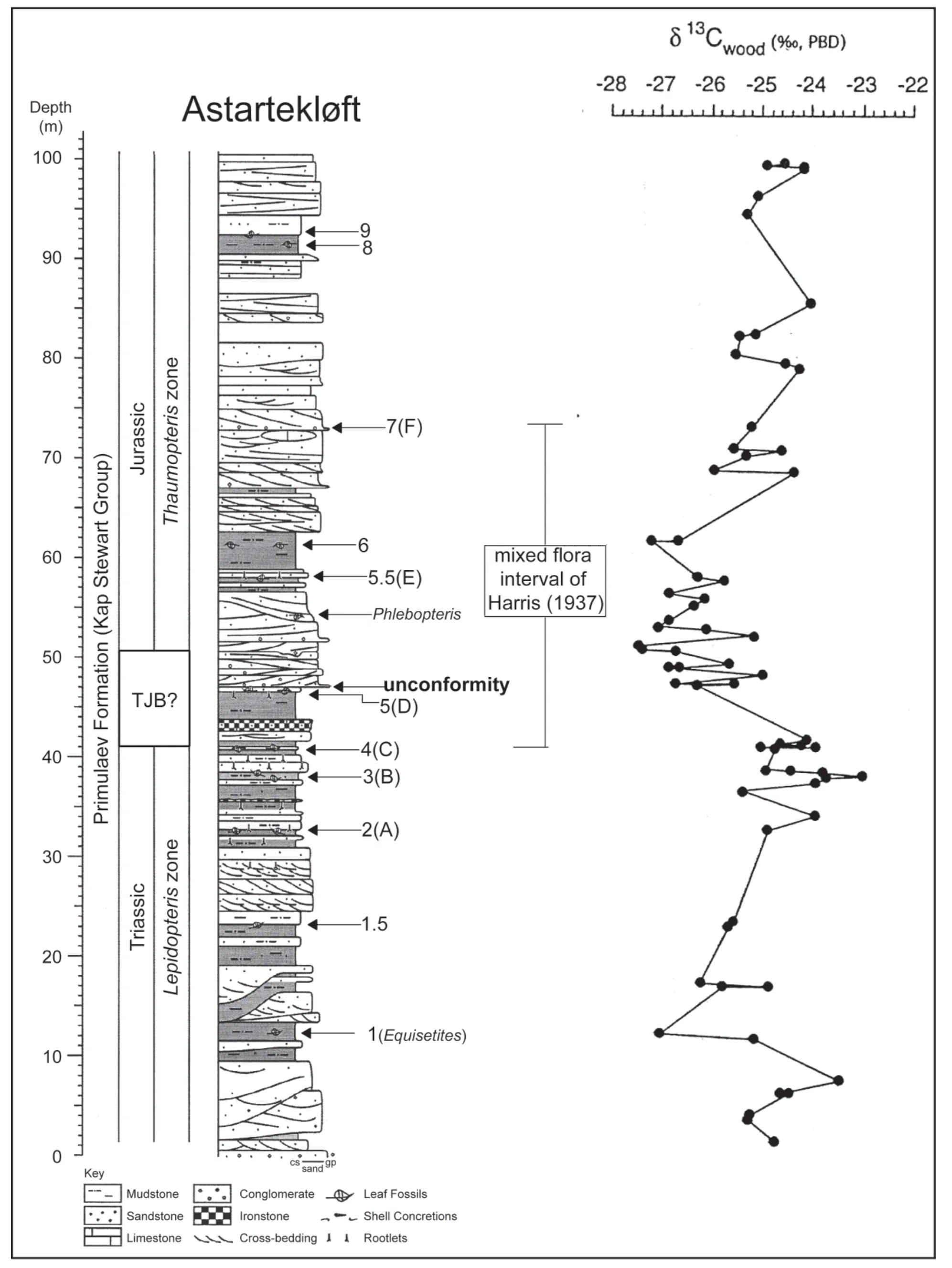
Figure 3 Measured secion of Triassic-Jurassic boundary interval of Kap Stewart Group in East Greenland showing distribution of megafossil plants and published carbon-isotope curve. The lithologic column is modified from McElwain et al. (2007) and shows megafossil plant beds using their scheme (1, 1.5, 2, etc.) followed by the scheme used by Harris (1937) in parentheses (Equisetites, A, B, etc.). The carbon isotope curve is from Hesselbo et al. (2002).
4.2 Plant extinctions in East Greenland
The Astartekløft section of the Primulaev Formation of the Kap Stewart Group in Jameson Land, East Greenland(Fig. 3) apparently encompasses the TJB and records the transition from theLepidopterisfloral zone to theThau‑matopterisfloral zone, with few species shared by both zones (Harris, 1937). The former is characterized by the presence of palynomorphs, includingRhaetipollis, while the latter containsHeliosporites(Pedersen and Lund,1980), and although extinction of some species across the transition between the two zones is evident, many species occur in both zones. Thus, Harris (1937: 76) emphasized that “real mixing” of the two floral zones occurs over a 10 m interval (Fig. 3), and “this is real mixing….the two sets of plants will be seen on the same bedding plane and they continue to be found together from top to bottom, of one of these beds….” Therefore, the data and analyses of Harris (1937) and Pedersen and Lund (1980) did not identify an abrupt change in the land plants across the TJB in Jameson Land. Furthermore, what species turnover looks abrupt in this section was attributed by Harris (1937) and Pedersen and Lund (1980) to species range truncations at a depositional hiatus, an unconformity also recognized in the Astartekløft section by Dam and Surlyk (1993) and Hesselboet al.(2002) (Fig. 3).
In contrast, in this section McElwain and collaborators claim: (1) a sudden, 85% drop in species diversity at the TJB (based on megafossil plants but not reflected in the palynomorph record); (2) a decrease in the relative abundance of common plant species at the TJB; (3) a change in leaf physiognomy to larger and rounder leaves; (4) an increase in charcoal indicative of wildfire at the TJB; and(5) an increase in chemical damage to sporomorphs at the TJB. They see all but possibly the last of these as indicative of a “super greenhouse” caused by CAMP (Central Atlantic Magmatic Province) eruptions.
We question, though, the validity of the placement of the TJB in the Astartekløft section. McElwain and collaborators place it at about the 47 m level of this section, basing that placement on the carbon isotope record published by Hesselboet al.(2002) (Fig. 3). This record shows an apparent, but modest (~2 ‰), negative excursion beginning at about the 45 m level. We note that this “excursion” coincides with an unconformity of unknown duration. Hence,the relationship of the isotopic trend above this hiatus to the missing, underlying strata is undetermined. Moreover,the general isotopic trend for this section bears little similarity to the published isotope stratigraphy for sections of putative identical age.
Nevertheless, Hesselboet al.(2002) assumed that the base of the excursion represents the “initial isotope excursion” and that the remainder records the “main isotope excursion” seen in marine sections that cross the TJB (e.g.,Ruhlet al., 2010). Therefore, the intervening section with a more positive isotopic trend (spanning 4 m at St. Audrie’s Bay in England) is conveniently missing, so we regard Hesselboet al.’s(2002) correlation as tenuous at best. Furthermore, if the initial negative excursion at the ~ 45 m level in the Astartekløft section is the well documented (in marine sections) negative excursion in carbon then it is not the TJB, but instead a point in the Rhaetian (e.g., Hillebrandtet al., 2013; Lucaset al., 2007b; Manderet al., 2013; Ruhlet al., 2010). Thus, all the events McElwain and collaborators associate with the TJB actually preceded it.
The chemostratigraphic correlation proposed by Hesselboet al.(2002) relies on the presence of an unconformity at 47 m in the Astartekløft section. Questions should be raised about the nature of this unconformity and the temporal magnitude of the hiatus it represents, as well as the associated substantial change in lithofacies (Fig. 3).Indeed, we note that most of the megafossil plant assemblages below that level are from mudrock-dominated intervals, whereas almost all of those above that level are from sandstone (Fig. 3). According to McElwainet al.(2007, p.551), the mudrock likely represents floodplain deposits of mainly autochthonous plants, whereas the sandstones represent channel and splay deposits that also include allochthonous plants. However, in evident contradiction to parts of their text and measured stratigraphic section, McElwainet al.(2007, table 1) identify all the plant localities up through their bed 5 as coming from “sheet splay” deposits,the plants from bed 6 as from a coal swamp (though no coal is present in the section at Astartekløft) and their plant beds 7 and 8 as coming from abandoned channels. In the absence of more data (e.g., detailed lithologic descriptions of the plant-bearing strata) and an actual sedimentological analysis of the plant-bearing beds, we are not able to resolve these evident contradictions in the study of McElwainet al.(2007).
Instead, it seems evident that the section at Astartekløft embodies a major lithofacies change at an unconformity,from a mudstone- and sheet-sandstone-dominated depositional system to a thick, channel-sandstone-dominated system (Fig. 3). On face value, it preserves two taphofloras in different lithofacies, and this raises the possibility that at least some of the apparent floral change inferred by McElwain and collaborators is due to the change in the depositional system (lithofacies). Assuming the TJB is present in this section near the 47 m level, the megafossil plant assemblages across the TJB are clearly not “isotaphonomic” (as claimed by McElwainet al., 2007), so they are not strictly comparable to each other in terms of composition,diversity and relative abundance.
According to McElwain and collaborators, a substantial perturbation of plant ecology and diversity is preserved at the TJB in East Greenland. We suggest that the data they present are indicative of a palaeogeographically local change in the paleoflora largely driven by lithofacies changes resulting in changing taphonomic filters. No catastrophic land plant extinction is documented and, at most,the floral turnover in East Greenland is nothing more than a palaeogeographically local event, as no similar, coeval event is documented elsewhere (Hallam and Wignall, 1997;Lucas and Tanner, 2008; Tanneret al., 2004). Furthermore,whatever happened to the megaflora in East Greenland, it happened very late in the Triassic, not at the TJB.
是的,他知道,刚才被拍下的那名少女,并非他的顾客,但他迟疑了一下,还是报出四张照片的价钱。他每个月都要来湖村给人拍照,这几个姑娘算得上他的熟客,而名叫夏小凡的,每次都跟着,不远不近,好像是看热闹,又好像对拍照这回事,充满好奇与渴望。也许是没钱,也许是害臊,她从未露出只言片语,请他替她拍一张。
4.3 Plant microfossils
Like the megafossil plant record, the palynological record provides no evidence for a mass extinction of land plants at the TJB (Bonis and Kürschner, 2012). Decades ago,Fisher and Dunay (1981) demonstrated that a significant proportion of theRhaetipollis germanicusassemblage that defines the Rhaetian in Europe (e.g., Orbell, 1973; Schuurman, 1979) persists in lowermost Jurassic strata, and this remains the case (e.g., Bonis and Kürschner, 2012; Boniset al., 2009; Cirilli, 2010; Kürschner and Herngreen, 2010).Brugman (1983) and Traverse (1988) thus concluded that floral turnover across the TJB was gradual, not abrupt.Kelber (1998) described the megaflora and palynoflora for Central Europe in a single unit he termed “Rhaeto-Liassic,”and concluded there was no serious disruption or decline in plant diversity across the TJB. More recently, Kuerschneret al.(2007) and Boniset al.(2009) documented the transitional nature of the change in palynomorphs across the TJB in the Northern Calcareous Alps of Austria (also see Hillebrandtet al., 2013), and the palynomorph record in East Greenland also does not document an abrupt TJB extinction(Manderet al., 2010, 2013).
Nevertheless, profound palynomorph extinction at the TJB was long identified in the Newark Supergroup record in eastern North America by Olsen and collaborators (Cornet,1977; Cornet and Olsen, 1985; Fowell and Olsen, 1993; Olsen and Sues, 1986; Olsenet al., 1990, 2002a, 2002b). Their work identified the TJB in the Newark section by a decrease in diversity of the plant microfossil assemblage, defined by the loss of palynomorphs considered typical of the Late Triassic, followed by dominance of the palynoflora by several species of the genus “Corollina” (=Classopollis), especiallyC. meyeriana(Cornet and Olsen, 1985; Fowell and Olsen,1993; Fowell and Traverse, 1995; Fowellet al., 1994; Olsenet al., 1990). They thus placed the TJB in the Newark section at the base of theClassopollis meyerianapalynofloral zone (Fig. 4). These workers attached considerable importance to a peak abundance of trilete spores at this level,which they termed a “fern spike” (Cornet and Olsen, 1985;Fowell and Olsen, 1993; Fowellet al., 1994; Whitesideet al., 2007). Although not present in all of the Newark basins(e.g., Cirilliet al., 2009), an increase in trilete spores has been noted in some European sections, where the acme zone is referred to as the Triletes Beds (Kuerschneret al.,2007; Ruhlet al., 2010; van de Schootbruggeet al., 2009).As noted by van de Schootbruggeet al.(2009), however,occurrences of a fern-dominated flora at the level of this apparent palynological turnover are limited strictly to the Northern Hemisphere.
This palynofloral change has been referred to as the “T-J palynofloral turnover” (Whitesideet al., 2007) or the “Passaic palynofloral event” (Lucas and Tanner, 2007b). But, as Kozur and Weems (2005: 33) well observed, “there are no age-diagnostic sporomorphs or other fossils to prove that this extinction event occurred at the Triassic-Jurassic boundary.” Indeed, Kuerschneret al.(2007) concluded that the Newark palynological event most likely represents an older, potentially early Rhaetian event, a conclusion shared by Kozur and Weems (2005, 2007, 2010) and by Lucas and Tanner (2007b). Thus, the palynological turnover in the Newark preceded the TJB (Fig. 4) and was a palaeogeographically regional event, not a global mass extinction.
Bonis and Kürschner (2012) provided a comprehensive review of TJB palynological records to conclude that they indicate vegetation changes that are non-uniform (different changes in different places), not synchronous and not indicative of a mass extinction of land plants. They attributed these changes to climate change driven by CAMP volcanism that produced a warmer climate and stronger monsoonal flow across Pangea, which developed drier interiors and wetter coastal regions. As they well put it, “…instead of a major and globally consistent palynofloral extinction event, the TJB is characterized by climate-induced quantitative changes in the sporomorph assemblages that vary regionally in magnitude and composition” (Bonis and Kürschner, 2012: 256). In summary, there is no evidence of a global mass extinction of land plants at the TJB.
5 Tetrapods
5.1 Overview
The idea of a substantial nonmarine tetrapod extinction at the TJB began with Colbert (1949, 1958), and has been more recently advocated by Olsenet al.(1987, 1990, 2002a,2002b). Nevertheless, Weems (1992), Benton (1994), Lucas(1994), Tanneret al.(2004) and Lucas and Tanner (2004,2007b, 2008) rejected this conclusion.
Colbert (1958) concluded that the temnospondyl amphibians, a significant component of many late Paleozoic and Early-Middle Triassic tetrapod assemblages, underwent complete extinction at the TJB. However, these temnospondyls are only a minor component of Late Triassic tetrapod assemblages, being of low diversity and relatively small numbers in most stratigraphic units (e.g., Hunt 1993;Long and Murry, 1995). Moreover, current data demonstrate the disappearance of most of the temnospondyls — capitosaurids, metoposaurids and latiscopids — at or just before the Norian-Rhaetian boundary, and only one family extinct at the end of the Triassic (plagiosaurids) (Milner, 1993,1994; Schoch and Milner, 2000). Late Triassic temnospondyl extinctions thus largely preceded the TJB.
To Colbert (1958), the bulk of the TJB tetrapod extinction was in the disappearance of the “thecodonts.” These“thecodonts” were more recently referred to as the crurotarsans (phytosaurs, aetosaurs and rauisuchians), though that name is no longer advocated by the latest cladistic analysis (Nesbitt, 2011). Phytosaurs, aetosaurs and rauisuchians are the only members of Colbert’s “thecodonts”to have a substantial Norian or younger fossil record based on current data. Other groups of thecodonts that have been portrayed by some as going extinct at the TJB either lack Rhaetian records (for example, the Ornithosuchidae:von Baczko and Ezcurra, 2013) or have so-called Rhaetian records of problematic age, such as records from British fissure-fill deposits (e.g., Fraser, 1994).
A non-“thecodont” group that did go extinct during the Rhaetian is the Procolophonidae (Fig. 5). However, procolophonids were a group of low diversity by Norian time, and many of their so-called Rhaetian records are from British fissure fills (e.g., Fraser, 1994). The tetrapod taxa from these fissure fills are mostly endemic taxa of no biochronological significance or cosmopolitan taxa with long stratigraphic ranges. As Lucas and Hunt (1994: 340) noted, “a single age should not necessarily be assigned to the fossils from one fissure and....individual fossils from the fissures may range in age from middle Carnian to Sinemurian.” We thus regard as problematic the precise age of many of the Triassic tetrapod fossils from the British fissure fills.
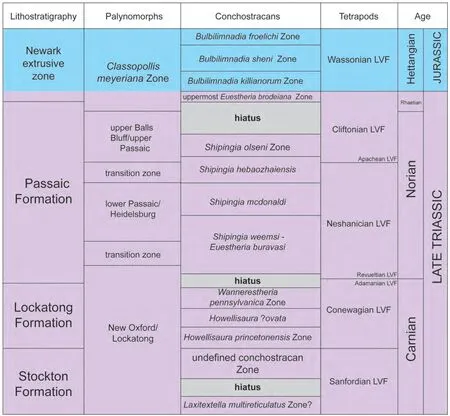
Figure 4 Biostratigraphic correlation of the Newark Supergroup section from the Late Triassic through the Triassic-Jurassic boundary.Note unconformities indicated by missing conchostracan zones. Modified from Kozur and Weems (2010), Lucas et al. (2012) and Weems and Lucas (2015).
Phytosaur fossils are known from Rhaetian-age strata in the Newark section, the Glen Canyon Group and the Germanic basin (e.g., Lucas and Tanner, 2007a; Olsenet al.,2002a, 2002b; Stocker and Butler, 2013). A Lower Jurassic phytosaur record has been documented by Maisch and Kapitzke (2010). This is an unabraded snout fragment found in the lowermost Jurassic pre-planorbis beds (Neophylliteszone) at Watchet in England. Stocker and Butler (2013) expressed skepticism about the stratigraphic provenance of this record, and it may be a reworked fossil, a possibility difficult to test. We accept that it documents phytosaur persistence into the earliest Jurassic, contradicting the longstanding assumption of their extinction at the TJB.
There are no demonstrable aetosaur or rauisuchian body fossils in Rhaetian strata (Desojoet al., 2013; Lucas,2010b; Nesbittet al., 2013), though Nesbittet al.(2013)mentioned a possible Early Jurassic rauisuchian from southern Africa, based on a single snout fragment that they also stated may be of a large crocodylomorph. Aetosaur and rauisuchian body fossil records thus suggest their extinction before Rhaetian time.
Nevertheless, the footprint ichnogenusBrachychirothe‑riumis known from Rhaetian strata, and Lucas and Heckert(2011) argued that an aetosaur was the trackmaker. Note that Smithet al.(2009) reported what they identified as chirothere tracks from the Lower Jurassic Elliot Formation of Lesotho in southern Africa, but as Klein and Lucas(2010) noted, these tracks differ from chirothere footprints in significant ways and are more likely large crocodylomorph tracks. The available stratigraphic data thus suggest a three-step extinction of the “crurotarsans” — first,rauisuchians by the end of the Norian, second, aetosaurs(based on the footprint ichnogenusBrachychirotherium) by the end of the Rhaetian and third, phytosaurs in the early Hettangian (Fig. 6).
Synapsids of the Late Triassic are dicynodonts and cynodonts (e.g., Abdala and Ribeiro, 2010; Lucas and Hunt,1994; Lucas and Wild, 1995). Most Late Triassic dicynodont records are Carnian, with a couple of post-Carnian records as young as late Norian (Dziket al.2008; Lucas, 2015; Lucas and Wild, 1995; Szulcet al., 2015). The Late Triassic cynodont record is more extensive, especially in Gondwana. Traversodontids have a relatively diverse record in the Carnian-Norian and a single Rhaetian record (note that the Gondwana cynodont-bearing strata assigned a Rhaetian age by Abdala and Ribeiro, 2010 are actually Norian: Lucas,2010b). Trithelodontids cross the TJB, and a problematic group of tooth-based taxa, the “dromatheriids,” disappears in the Rhaetian (Lucas and Hunt, 1994). Note, though, that the origin and diversification of mammaliaforms, which likely include the “dromatheriids,” began in the Late Triassic (e.g., Newhamet al., 2014). On present evidence, a TJB synapsid extinction is thus of a handful of taxa, with most taxa extinct by Rhaetian time, but others crossing the TJB.
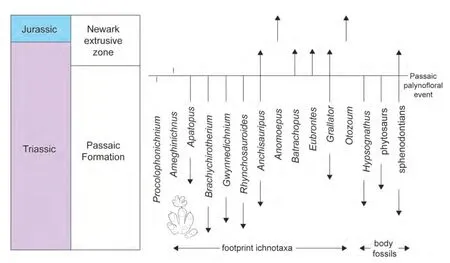
Figure 5 Stratigraphic ranges of tetrapod footprint ichnogenera and body fossil taxa across the Late Triassic (Rhaetian) Passaic palynofloral event in the Newark section. Modified and updated after Olsen et al. (2002a).
5.2 Newark tetrapod record
The recent case for a mass extinction of tetrapods at the TJB has relied heavily on the Newark Supergroup record of tetrapod fossils (e.g., Olsen and Sues 1986; Olsenet al., 1987, 1990, 2002a, 2002b; Szajna and Silvestri,1996; Silvestri and Szajna, 1993) (Fig. 5). However, a close examination of all Triassic tetrapod taxa known from the Newark Supergroup identifies only three body-fossil taxa(indeterminate phytosaurs, the procolophonidHypsogna‑thusand sphenodontians) in the youngest Triassic strata,which are strata of Rhaetian age (Huberet al., 1993; Kozur and Weems, 2010; Lucas and Huber, 2003). Recent collecting has not changed that, and only a few fragmentary tetrapod fossils are known from the Newark extrusive zone and are not age diagnostic (Lucas and Huber, 2003; Lucas and Tanner, 2007b).
The McCoy Brook Formation, which overlies the only CAMP basalt of the Fundy basin in Nova Scotia, yields a tetrapod assemblage generally considered Early Jurassic in age by vertebrate paleontologists (Fedaket al., 2015; Lucas, 1998; Lucas and Huber, 2003; Lucas and Tanner, 2007a,2007b; Olsenet al., 1987; Shubinet al., 1994). However,this assemblage could straddle the marine-defined TJB given that Cirilliet al.(2009) demonstrated that at least the lowermost McCoy Brook strata (lower part of Scots Bay Member) are Rhaetian.
Indeed, very recently, Sues and Olsen (2015) and Fedaket al.(2015) have assigned a latest Triassic (Rhaetian) age to an assemblage of vertebrate fossils from the “fish bed”in the Scots Bay Member of the McCoy Brook Formation that includes the crocodylomorphProtosuchusand the tritylodontOligokyphus. These two taxa are known only from Jurassic strata elsewhere (e.g., Lucas and Tanner, 2007a),and their assignment to the Rhaetian is based on inferred cyclostratigraphic correlations (Sues and Olsen, 2015), not on direct association with Triassic fossils. Therefore, we question the age assignment of the “fish bed” to the Triassic, though we also note that if correctly assigned a Rhaetian age, it pushes the temporal record ofProtosuchusand ofOligokyphusback into the Triassic, thus diminishing the apparent turnover in tetrapod taxa across the TJB.
Because the Newark body fossil record of tetrapods is sparse across the TJB and inadequate to evaluate a possible tetrapod extinction, the tetrapod footprint record in the Newark Supergroup has been used as a proxy (e.g.,Olsen and Sues, 1986; Olsenet al., 2002a,2002b; Szajna and Silvestri, 1996). However, detailed stratigraphic study of the Newark footprint record indicates nothing more than moderate turnover in the footprint assemblage at a within-Rhaetian stratigraphic level below the lowest CAMP basalt sheet (Fig. 5). Similar changes in tetrapod footprint assemblages are also known from the Chinle Group-Glen Canyon Group section of the American Southwest and from the Germanic Basin (e.g., Lucaset al., 2006a; Lucas, 2007).
The footprint turnover in the Newark section (Fig. 5) is supposedly the disappearance of four ichnogenera in the uppermost Passaic Formation, and the appearance of three ichnogenera at that datum (Olsenet al., 2002a, 2002b). The ichnogenera that disappear represent phytosaurs (Apatopus:Klein and Lucas, 2013), aetosaurs (Brachychirotherium: Lucas and Heckert, 2011) and tanystropheids (Gwynnedichni‑um: Lucaset al., 2014). There are single Newark records ofProcolophonichnium(procolophonid: Baird, 1986) just below the turnover level and a single record ofAmeghinichnus(mammaliaform: de Valais, 2009) above the level.
According to Olsenet al.(2002a, 2002b), the ornithischian dinosaur footprint ichnogenusAnomoepusfirst appears at this level, but a later detailed review of the ichnogenus by Olsen and Rainforth (2003) indicated that the lowest stratigraphic record ofAnomoepusis stratigraphically higher, in the Newark extrusive zone (Fig. 5). The crocodylomorph footprint ichnogenusBatrachopusappears at the turnover level, but there are older Triassic body fossil records of crocodylomorphs (Klein and Lucas, 2010;Nesbittet al., 2013). Olsenet al.(2002a, 2002b) showed the prosauropod-dinosaur-footprint ichnogenusOtozoumappearing in the upper Passaic Formation, but a later revision of the ichnogenus by Rainforth (2003) established its stratigraphically lowest record as Jurassic, in the Newark extrusive zone (Fig. 5). The lacertoid footprint ichnogenusRhynchosauroideshas its last Newark record in the upper Passaic Formation, but this ichnogenus has Jurassic records elsewhere (Avanziniet al., 2010).
Thus, what the Newark tetrapod footprint and body-fossil record show is the local extinction of phytosaurs (they have a younger record elsewhere), aetosaurs, tanystropheids and procolophonids (this may be the level of their global extinction). That is the extent of the turnover in tetrapod taxa it documents, and the turnover level in the Newark is at a Rhaetian horizon, not at the TJB (Fig. 5).
Part of the footprint turnover in the Newark section is the local lowest occurrence of the theropod footprint ichnogenusEubrontes(as defined by Olsenet al., 1998,i.e.,tridactyl theropod pes tracks longer than 28 cm). For decades, much was made of this record ofEubrontes. Thus,Olsen and Galton (1984) concluded that the lowest occurrence ofEubrontesmarks the base of the Jurassic, and Olsenet al.(2002a, 2002b) later argued that the sudden appearance ofEubrontesin the “earliest Jurassic” strata of the Newark Supergroup indicates a dramatic size increase in theropod dinosaurs at the TJB. They interpreted this as the result of a rapid (thousands of years) evolutionary response by the theropod survivors of a mass extinction and referred to it as “ecological release” (Olsenet al., 2002a:1307). They admitted that this can be invalidated by the description ofDilophosaurus-sized theropods or diagnosticEubrontestracks in verifiably Triassic-age strata.
Indeed, tracks of large theropod dinosaurs assigned toEubrontes(or its possible synonymKayentapus) are known from the Triassic of Australia, Africa (Lesotho), Europe(Great Britain, France, Italy, Germany, Poland-Slovakia,Scania) and eastern Greenland, invalidating the “ecological release” hypothesis (Bernardiet al.2013; Lucaset al.,2006b; Niedźwiedzki, 2011). A detailed review of these records indicates Carnian, Norian and Rhaetian occurrences of tracks that meet the definition ofEubrontesestablished by Olsenet al.(1998). Also, theropods large enough to have made at least someEubrontes-size tracks have long been known from the Late Triassic body-fossil record (e.g.,Langeret al., 2009). Thus, the sudden abundance of these tracks in the Newark Supergroup cannot be explained simply by the rapid evolution of small theropods to large size following a mass extinction. The concept of a sudden appearance ofEubrontestracks due to “ecological release”at the TJB thus proposed by Olsenet al.(2002a, 2002b) can be abandoned.
5.3 Conclusions
With new and more detailed stratigraphic data, the long perceived TJB tetrapod extinction looks mostly like an artifact of coarse temporal resolution, the compiled correlation effect of Lucas (1994). Thus, Colbert (1958) knew,and we still know, that Late Triassic tetrapod assemblages in Laurussian locales are dominated by phytosaurs, with lesser numbers of aetosaurs, rauisuchians and dinosaurs.Some Gondwanan Late Triassic assemblages are cynodont(mostly traversodontid) and rhynchosaur dominated. In contrast, well known Early Jurassic tetrapod assemblages are dinosaur dominated, with lesser numbers of crocodylomorphs and cynodonts (tritylodontids), and no phytosaurs,aetosaurs or rauisuchians. But, the former are Norian, and the latter are Sinemurian age assemblages, separated in time by at least 6 million years.
Given the lack of evidence for TJB plant and arthropod extinctions, what could have caused a TJB tetrapod extinction? Aetosaurs, some of the cynodonts and all of the dicynodonts perceived of as suffering extinction are inferred herbivores, whereas the phytosaurs and rauisuchians are inferred predators. Given that many herbivores — notably the prosauropod dinosaurs — continue through the TJB, a sudden trophic collapse of vertebrate communities on land cannot be supported. Instead, the extinctions of tetrapods during the Late Triassic appear to be a prolonged series of events that began in the Norian and extended into the Hettangian (Fig. 6).
6 Ecological severity
McGheeet al.(2004, 2013) argued that mass extinctions need to be evaluated not just as biodiversity crises(crashes), but also in terms of their ecological severity. In their scheme of ecological severity, they considered the TJB nonmarine extinction to be in their category I or IIa.Category I means that ecosystems before the extinction were replaced by new ecosystems post-extinction, whereas category IIa means that the extinctions caused permanent loss of major ecosystem components. Clearly, as we have pointed out earlier (Lucas and Tanner, 2008), this is a gross overestimate of the ecological significance of whatever extinctions took place on land across the TJB.
McGheeet al.(2004) concluded that the TJB involved a rapid ecological replacement of Triassic mammal-like reptiles and rhynchosaurs by dinosaurs. However, the stratigraphically highest rhynchosaurs are Revueltian (earlymiddle Norian), and the group was largely gone by the end of the Carnian (Hunt and Lucas, 1991; Lucaset al., 2002;Spielmannet al., 2013). Late Norian dicynodonts are now known (Dziket al., 2008; Szulcet al., 2015) and document a small presence in the post Carnian, unless a putative Cretaceous record (with problematic provenance) from Australia is verified (Thulborn and Turner, 2003). The other principal group of Late Triassic mammal-like reptiles, the cynodonts, were of low diversity after the Carnian-Norian(Abdala and Ribeiro, 2010; Lucas and Hunt, 1994). Dinosaurs appeared as body fossils in the Carnian and began to diversify substantially in some parts of Pangea by the late Norian (e.g., Langer, 2014; Langeret al., 2009).
Thus, the ecological severity of the end-Triassic tetrapod extinction is relatively low (Category IIb on the McGheeet al., 2004 classification), and the plant extinctions do not look like they were ecologically severe either (see above).Clearly, there was no substantial disruption of the terrestrial ecosystem (excluding some local perturbations) across the TJB.
7 The role of CAMP volcanism
In the marine realm, the end-Triassic drop in diversity was selective but notable for the rapid loss of specific taxa,such as conodonts, radiolarians, infaunal bivalves and ammonoids, suggesting physical processes that strongly affected ocean bioproductivity (Tanneret al., 2004). For many years, the hypothesis was advanced that the extinctions were associated with a single catastrophic cause, specifically, the Manicouagan bolide impact, the remnant crater of which spans ~ 70 km in southern Quebec, Canada (Olsenet al., 1987, 2002a, 2002b). However, this hypothesis was firmly disproved by radioisotopic dating of the Manicouagan structure, which showed that the impact occurred ~ 14 Ma prior to the TJB (Hodych and Dunning, 1992; Onoueet al.,2012; Ramezaniet al., 2005).
Subsequently, attention on the TJB “extinctions” has focused primarily on the environmental effects of the eruption of the flood basalts of the Central Atlantic Magmatic Province (CAMP). Formerly, the onset of CAMP eruptive activity was assumed to post-date the TJB (Whitesideet al.,2007), but it is now well-established that these eruptions span the boundary (Cirilliet al., 2009; Kozur and Weems,2005, 2007, 2010; Lucas and Tanner, 2007b). Within the basins of the Newark Supergroup, the eruptions proceeded in three main episodes, separated by eruptive hiatuses during which sediments accumulated, with the majority of the total volume ejected during the initial eruptive episode (Marzoliet al., 2011).
The total duration of the CAMP eruptions has been estimated at ~600 ky by cyclostratigraphy (Olsenet al., 1996).The initial estimate of the size of the province, by McHone and Puffer (1996), was ~2.3 × 106km2, but Deckartet al.(1997) and Marzoliet al.(1999) extended the boundaries of the province to include all areas near the rift margin containing mafic intrusions of approximately correlative age,ultimately outlining an area of 11 × 106km2. Defining the vast potential aerial extent of CAMP led to an erupted lava volume calculation of ~2 × 106km3, similar to the volume calculated for the Deccan Traps. As noted by Tanneret al.(2004), however, this calculation entails the assumption that the entire area was covered by lava to an average depth of hundreds of meters. Even in the Newark basin, however,where the entire erupted thickness is preserved, the total thickness varies widely, from > 1 km to as little as tens of meters. Therefore, we regard this assumption as untenable.
It is widely acknowledged that floral and faunal turnover at the end of the Triassic was likely driven by the CAMP eruptions, which caused a significant environmental perturbation through outgassing (e.g., Belcheret al., 2010;Blackburneet al., 2013; Boniset al., 2009; Haworth and Raschi, 2014; Ruhlet al., 2010; Schoeneet al., 2010; van de Schootbruggeet al., 2009; Tanneret al., 2004). Attention has focused largely on the potential for greenhouse warming produced by the outgassed CO2(Berner and Beerling, 2002; McElwainet al., 1999; Steinthorsdottiret al.,2011), but many of these studies ignore the constraints of the likely volume of erupted lava and gas content. Yapp and Poths (1996), for example, estimated a catastrophic 18-fold increase in atmospheric CO2across the TJB, whereas Tanneret al.(2001) calculated a much more modest increase(of several hundred ppm), and more recently, Steinthorsdottiret al.(2011) calculated a CO2doubling (by 2000 to 2500 ppm). The tremendous divergence in the published estimates of atmospheric CO2loading from CAMP outgassing stems from the different approaches used to generate the calculations. McElwain and collaborators (e.g., McElwainet al., 1999; Steinthorsdottiret al., 2011) rely primarily on the stomatal response of plants to elevated CO2levels, while others (Clevelandet al., 2008; Schalleret al., 2012; Tanneret al., 2001) utilize the isotopic composition of pedogenic carbonates. These approaches also yield diverging estimates for the pre-eruption atmospheric composition,knowledge of which is essential to determining the warming effects of CAMP outgassing. Thus, there is as yet no agreement on the mass of CO2released by the CAMP eruptions,much less agreement on the climatic consequences.
Tanneret al.(2004, 2007) drew attention to the lack of consideration of the effects of SO2outgassing by the CAMP eruptions, and further demonstrated that high atmospheric SO2loading also can force changes in plant stomatal frequency, an effect eventually noticed by other workers (Baconet al., 2013). The environmental effects of large sulfur emissions during prolonged flood basalt eruptions are not clear, but the formation of H2SO4aerosols, which in addition to causing acidic precipitation, are known to increase atmospheric opacity and result in reduced short-wave radiant heating, causing global cooling (Sigurdsson, 1990). Fluorine and chlorine volatile emissions during the CAMP eruptionsalso would have contributed to the acidification of surface waters and terrestrial environments (Guexet al., 2004; Lucas and Tanner, 2008; van de Schootbruggeet al., 2009).
Thus, a temporary loss of phytoplankton productivity in marine surface waters is an entirely reasonable consequence of the CAMP eruptions. The subsequent “trickledown” effects through the marine trophic system are clear,even without climate change. Primary producers would have been most profoundly affected by the changes in water chemistry, but higher level consumers would also have felt ecological pressure, although perhaps less strongly. The compounding effects of climate change-rapid, short-term cooling forced by H2SO4aerosols and slower, longer-term warming driven by CO2- would have exacerbated the environmental stresses on the ecosystems. Thus, floral and faunal turnover at the end of the Triassic was likely driven by the CAMP eruptions, which caused significant environmental perturbations (cooling, warming, acidification) through outgassing, but the effects on the nonmarine biota appear to have been palaeogeographically localized, transient and not catastrophic.
8 End Triassic nonmarine biotic events
The Late Triassic was a time of elevated extinction rates and low origination rates in many biotic groups (e.g., Bambachet al., 2004; Kiesslinget al., 2007; Lucas and Tanner,2008). Most of the evidence for this comes from the marine biota, with multiple Late Triassic, and often substantial extinctions of radiolarians, conodonts, bivalves, ammonoids and reef-building organisms. In the marine realm,these events are: (1) early-middle Carnian boundary (the“Carnian crisis”), which included major extinctions of crinoids (especially the Encrinidae), echinoids, some bivalves(scallops), bryozoans, ammonoids, conodonts and a major change in the reef ecosystem; (2) a substantial extinction of conodonts, ammonoids and some bivalves (especially pectinids) at the Carnian-Norian boundary; (3) within and at the end of the Norian, when there are several extinction events that had a particularly profound effect on conodonts, marine bivalves and ammonoids; and (4) a final set of stepwise extinctions, particularly of ammonoids and conodonts, within and at the end of the Rhaetian. (e.g.,Flügel, 2002; Hallam, 1995; Hornunget al., 2007; Johnson and Simms, 1989; Lucas and Tanner, 2008; Schäfer and Fois,1987; Tanneret al., 2004)
On land, plant and tetrapod extinctions were much less dramatic. Within the Carnian most workers envision a humid phase (or “pluvial”) as a possible cause of some turnover of terrestrial taxa, at least in Europe (e.g., Hornunget al., 2007; Rigoet al., 2007; Simms and Ruffell, 1989,1990), though this has been disputed (e.g., Visscheret al.,1994). There is an end-Carnian event in the terrestrial tetrapod record, with some evolutionary turnover across the Carnian-Norian boundary (Benton, 1986, 1991, 1993; Lucas,1994). Extinctions on land closer to the TJB look even less significant. The amphibian, archosaur and synapsid extinctions of the Late Triassic are not concentrated at the TJB,but instead occur stepwise, beginning in the Norian and extending into the Jurassic (Fig. 6). Changes in the land plants are complex, diachronous and likely climate driven evolutionary changes over background extinction rates, not a serious extinction or set of extinctions.
We conclude that no significant extinction took place on land at the TJB. Indeed, the idea of a single mass extinction on land at the TJB has led to a search for the cause of the “mass extinction” and drawn attention away from what were actually a series of biotic events that took place throughout the Late Triassic (Fig. 6). Research should now focus on these multiple extinctions and their causes, not on a single extinction event. Perhaps the most interesting question not yet addressed by most researchers is why a prolonged (at least 30 million years) interval of elevated extinction rates occurred during the Late Triassic.
Acknowledgements
We are grateful to Baas van de Schootbrugge for encouraging us to publish this article. We also thank Professor Li,Jin-Geng Sha, Shu-Zhong Shen and two anonymous reviewers for their helpful reviews of this article.
1. Abdala, F., Ribeiro, A. M., 2010. Distribution and diversity patterns of Triassic cynodonts (Therapsida, Cynodontia) in Gondwana.Palaeogeography,Palaeoclimatology,Palaeo‑ecology, 286, 202-217.
2. Ash, S., 1986. Fossil plants and the Triassic-Jurassic boundary, in: Padian, K. (Ed.), The Beginning of the Age of Dinosaurs.Cambridge University Press, Cambridge, pp. 21-30.
3. Avanzini, M., Pinuela, L., Garcia-Ramos, J. C., 2010. First report of a Late Jurassic lizard-like footprint (Asturias, Spain).Journal of Iberian Geology, 36, 175-180.
4. Bacon, K. L., Belcher, C. M., Haworth, M., McElwain, J. C.,2013. Increased atmospheric SO2detected from changes in leaf physiognomy across the Triassic-Jurassic boundary interval of East Greenland. PLOS One, 8(4), e60614.
5. von Baczko, M. B., Ezcurra, M. D., 2013. Ornithosuchidae:A group of Triassic archosaurs with a unique ankle joint, in:Nesbitt, S. J., Desojo, J. B., Irmis, R. B., (Eds.), Anatomy,phylogeny and palaeobiology of Early Archosaurs and their kin.Geological Society,London,Special Publications, 379,187-202.
6. Baird, D., 1986. Some Upper Triassic reptiles, footprints, and an amphibian from New Jersey.The Mosasaur, 3, 125-153.
7. Balini, M., Lucas, S. G., Jenks, J. M., Spielmann, J. A., 2010.Triassic ammonoid biostratigraphy: An overview, in: Lucas, S.G. (Ed.), The Triassic timescale.Geological Society,London,Special Publications, 334, 221-262.
8. Bambach, R. K., Knoll, A. H., Wang, S. C., 2004. Origination,extinction, and mass depletion of marine diversity.Paleobiol‑ogy, 20, 522-542.
9. Bandyopadhyay, S., Sengupta, D. P., 2006. Vertebrate faunal turnover during the Triassic-Jurassic transition: An Indian scenario.New Mexico Museum of Natural History and Science Bulletin, 37, 77-85.
10. Belcher, C. M., Mander, L., Rein, G., Jervis, F. X., Haworth,M., Hesselbo, S. P., Glasspool, I. J., McElwain, J. C., 2010.Increased fire activity at the Triassic/Jurassic boundary in Greenland due to climate-driven floral change.Nature Geo‑science, 3, 426-429.
11. Benton, M. J., 1986. More than one event in the Late Triassic mass extinction.Nature, 321, 857-861.
12. Benton, M. J., 1991. What really happened in the Late Triassic?Historical Biology, 5, 263-278.
13. Benton, M. J., 1993. Reptilia, in: Benton, M. J., (Ed.), The Fossil Record 2. Chapman and Hall, London, pp. 681-715.
14. Benton, M. J., 1994. Late Triassic to Middle Jurassic extinctions among continental tetrapods: Testing the pattern, in:Fraser, N. C., Sues, H-D., (Eds.), In the Shadow of the Dinosaurs. Cambridge University Press, Cambridge, pp. 366-397.
15. Bernardi, M., Petti, F. M., Porchetti, S. D., Avanzini, M., 2013.Large tridactyl footprints associated with a diverse ichnofauna from the Carnian of the Southern Alps.New Mexico Mu‑seum of Natural History and Science Bulletin, 61, 48-54.
16. Berner, R. A., Beerling, D. J., 2007. Volcanic degassing necessary to produce CaCO3undersaturated ocean at the Triassic-Jurassic boundary.Palaeogeography,Palaeoclimatology,Pal‑aeoecology, 244, 368-373.
17. Blackburn, T J., Olsen, P. E., Bowring, S. A., McLean, N. M.,Kent, D. V., Puffer, J., McHone, G., Rasbury, E. T., Et-Touhami, M., 2013. Zircon U-Pb geochronology links the end-Triassic extinction with the Central Atlantic Magmatic Province.Sci‑ence, 340, 941-945.
18. Bonis, N. R., Kürschner, W. M., 2012. Vegetation history, diversity patterns, and climate change across the Triassic/Jurassic boundary.Paleobiology, 38, 240-264.
19. Bonis, N. R., Kürschner, W., Krystyn, L., 2009. A detailed palynological study of the Triassic-Jurassic transition in key sections of the Eiberg basin (Northern calcareous Alps, Austria).Review of Palaeobotany and Palynology, 156, 376-400.
20. Brugman, W. A., 1983. Permian-Triassic Palynology. State University Utrecht, Utrecht, pp. 1-121.
21. Burgoyne, P. M., van Wyk, A. E., Anderson, J. M., Schrire,B. D., 2005. Phanerozoic evolution of plants on the African plate.Journal of African Earth Sciences, 43, 13-52.
22. Cascales-Miñana, B., Cleal, C. J., 2012. Plant fossil record and survival analysis.Lethaia, 45, 71-82.
23. Cirilli, S., 2010. Uppermost Triassic-lowermost Jurassic palynology and palynostratigraphy: A review, in: Lucas, S. G.,(Ed.), The Triassic timescale.Geological Society,London,Special Publications, 334, 285-314.
24. Cirilli, S., Marzoli, A., Tanner, L. H., Bertrand, H., Buratti,N., Jourdan, F., Bellieni, G., Kontak, D., Renne, P. R., 2009.Late Triassic onset of the Central Atlantic Magmatic Province (CAMP) volcanism in the Fundy Basin (Nova Scotia): New stratigraphic constraints.Earth and Planetary Science Let‑ters, 286, 514-525.
25. Cleal, C. J., 1993a. Pteridophyta, in: Benton, M. J., (Ed.),The Fossil Record 2. Chapman and Hall, London, pp. 779-794.
26. Cleal, C. J., 1993b. Gymnospermophyta, in: Benton, M. J.,(Ed.), The Fossil Record 2. Chapman and Hall, London, pp.795-808.
27. Cleveland, D. M., Nordt, L. C., Dworkin, S. I., Atchley, S. C.,2008. Pedogenic carbonate isotopes as evidence for extreme climatic events preceding the Triassic-Jurassic boundary: Implications for the biotic crisis?GSA Bulletin, 120, 1408-1415.
28. Colbert, E. H., 1949. Progressive adaptations as seen in the fossil record, in: Jepsen, G. L., Mayr, E., Simpson, G. G.,(Eds.), Genetics, Paleontology and Evolution. Princeton University Press, Princeton, pp. 390-402.
29. Colbert, E. H., 1958. Triassic tetrapod extinction at the end of the Triassic Period.Proceedings National Academy of Science USA, 44, 973-977.
30. Cornet, B., 1977. The Palynostratigraphy and Age of the Newark Supergroup. Ph.D. Thesis. Pennsylvania State University,University Park, PA, 1-505.
31. Cornet, B., Olsen, P. E., 1985. A summary of the biostratigraphy of the Newark Supergroup of eastern North America with comments on provinciality, in: Weber, R., (Ed.), III Congreso Latinoamericano de Paleontologia Mexico, Simposio Sobre Floras del Triasico Tardio, su Fitogeografia y Paleoecologia,UNAM Instituto de Geologia, Mexico City, pp. 67-81.
32. Dam, G., Surlyk, F., 1993. Cyclic sedimentation in a large wave- and storm-dominated anoxic lake, Kap Stewart Formation (Rhaetian-Sinemurian), Jameson Land, East Greenland.International Association of Sedimentologists Special Publi‑cation, 18, 419-448.
33. Deckart, K., Fèraud, G., Bertrand, H., 1997. Age of Jurassic continental tholeiites of French Guyana, Surinam and Guinea:Implications for the initial opening of the Central Atlantic Ocean.Earth and Planetary Science Letters, 150, 205-220.
34. Desojo, J. B., Heckert, A. B., Martz, J. W., Parker, W. G.,Schoch, R. R., Small, B. J., Sulej, T., 2013. Aetosauria: A clade of armoured pseudosuchians from the Upper Triassic continental beds, in: Nesbitt, S. J., Desojo, J. B., Irmis, R. B.,(Eds.), Anatomy, phylogeny and palaeobiology of early archosaurs and their Kin.Geological Society,London,Special Pub‑lications, 379, 203-240.
35. Dzik, J., Sulej, T., Niedzwiedzki, G., 2008. A dicynodonttheropod association in the latest Triassic of Poland.Acta Pal‑aeontologica Polonica, 53, 733-738.
36. Edwards, D., 1993. Bryophyta, in: Benton, M. J., (Ed.), The Fossil Record 2. Chapman and Hall, London, pp. 775-778.
37. Fedak, T. J., Sues, H.-D., Olsen, P. E., 2015. First record of the tritylodontid cynodont Oligokyphus and cynodont postcranial bones from the McCoy Brook Formation of Nova Scotia,Canada.Canadian Journal of Earth Sciences, 52, 244-249.
38. Fisher, M. J., Dunay, R. E., 1981. Palynology and the Triassic/Jurassic boundary.Review of Palaeobotany and Palynology,34, 129-135.
39. Flügel, E., 2002. Triassic reef patterns, in: Kiessling, W.,Flügel, E., Golonka, J., (Eds.), Phanerozoic reef patterns,SEPM Special Publication, 72, 391-463.
40. Fowell, S. J., Olsen, P. E., 1993. Time calibration of Triassic-Jurassic microfloral turnover, eastern North America.Tec‑tonophysics, 222, 361-369.
41. Fowell, S. J., Traverse, A., 1995. Palynology and age of the upper Blomidon Formation, Fundy basin, Nova Scotia.Review of Palaeobotany and Palynology, 86, 211-233.
42. Fowell, S. J., Cornet, B., Olsen, P. E., 1994. Geologically rapid Late Triassic extinctions: palynological evidence from the Newark Supergroup.Geological Society of America Special Paper, 288, 197-206.
43. Fraser, N. C., 1994. Assemblages of small tetrapods from British Late Triassic fissure deposits, in: Fraser, N. C., Sues, H-D.,(Eds.), In the Shadow of the Dinosaurs. Cambridge University Press, Cambridge, pp. 214-226.
44. Galli, M. T., Jadoul, F., Bernasconi, S. M., Weissert, H., 2005.Anomalies in global carbon cycling and extinction at the Triassic/Jurassic boundary: Evidence from a marine C-isotope record.Palaeogeography,Palaeoclimatology,Palaeoecology,16, 203-214.
45. Giordano, N., Rigo, M., Ciarapica, G., Bertinelli, A., 2010.New biostratigraphical constraints for the Norian/Rhaetian boundary: Data from Lagonegro basin, Southern Appenines,Italy.Lethaia, 43, 573-586.
46. Gómez, J. J., Goy, A., Barrón, E., 2007. Events around the Triassic-Jurassic boundary in northern and eastern Spain: A review.Palaeogeography,Palaeoclimatology,Palaeoecology,244, 89-110.
47. Grimaldi, D., Engel, M. S., 2005. Evolution of the Insects.Cambridge University Press, Cambridge, pp. 1-755.
48. Guex, J., Bartolin, A., Atudorei, V., Taylor, D., 2004. Highresolution ammonite and carbon isotope stratigraphy across the Triassic-Jurassic boundary at New York Canyon (Nevada).Earth and Planetary Science Letters, 225, 29-41.
49. Hallam, A., 1995. Major bio-events in the Triassic and Jurassic, in: Walliser, O., (Ed.), Global Events and Event Stratigraphy. Springer, Berlin, pp. 265-283.
50. Hallam, A., 2002. How catastrophic was the end-Triassic mass extinction?Lethaia, 35, 147-157.
51. Hallam, A., Wignall, P. B., 1997. Mass Extinctions and Their Aftermath. Oxford University Press, Oxford, pp.1-320.
52. Harris, T. M., 1937. The fossil flora of Scoresby Sound East Greenland, Part 5: Stratigraphic relations of the plant beds.Meddelelser om Grønland, 112, 1-112.
53. Haworth, M., Raschi, A., 2014. An assessment of the use of epidermal micro-morphological features to estimate leaf economics of Late Triassic-Early Jurassic fossil Ginkgoales.Review of Palaeobotany and Palynology, 205, 1-8.
54. Hesselbo, S. P., Robinson, S. A., Surlyk, F., Piasecki, S., 2002.Terrestrial and marine extinction at the Triassic-Jurassic boundary synchronized with major carbon-cycle perturbation: A link to initiation of massive volcanism?Geology, 30,251-254.
55. Hillebrandt, A. v., Krystyn, L., Kürschner, W. M., Bonis, N. R.,Ruhl, M., Richoz, S., Schobben, M. A. N., Ulrichs, M., Bown, P.R., Kment, K., McRoberts, C. A., Simms, M., Tomãsovych, A.,2013. The global stratotype section and point (GSSP) for the base of the Jurassic System at Kuhjoch (Karwendel Mountains,Northern Calcareous Alps, Tyrol, Austria).Episodes, 36, 162-198.
56. Hodych, J. P., Dunning, G. R., 1992. Did the Manicouaga impact trigger end-of-Triassic mass extinction?Geology, 20, 51-54.
57. Hornung, T., Brandner, R., Krystyn, L., Jaochimski, M. M.,Keim, L., 2007. Multistratigraphic constrainst on the NW Tethyan “Carnian crisis”.New Mexico Museum of Natural History and Science Bulletin, 41: 59-67.
58. Hounslow, M. W., Muttoni, G., 2010. The geomagnetic polarity timescale for the Triassic: Linkage to stage boundary definitions, in: Lucas, S. G., (Ed.), The Triassic timescale.Geo‑logical Society,London,Special Publications, 334, 61-102.
59. Hounslow, M. W., Posen, P. E., Warrington, G., 2004. Magnetostratigraphy and biostratigraphy of the Upper Triassic and lowermost Jurassic succession, St. Audrie’s Bay, UK.Palaeo‑geography,Palaeoclimatology,Palaeoecology, 213, 331-358.
60. Huber, P., Lucas, S. G., Hunt, A. P., 1993. Vertebrate biochronology of the Newark Supergroup Triassic, eastern North America.New Mexico Museum of Natural History and Science Bulletin, 3, 179-186.
61. Hunt, A. P., 1993. A revision of the Metoposauridae (Amphibia:Temnospondyli) of the Late Triassic with description of a new genus from the western United States.Museum of Northern Arizona Bulletin, 59, 67-97.
62. Hunt, A. P., Lucas, S. G., 1991. A new rhynchosaur from West Texas (USA) and the biochronology of Late Triassic rhynchosaurs.Palaeontology, 34, 191-198.
63. Johnson, L. A., Simms, M. J., 1989. The timing and cause of Late Triassic marine invertebrate extinctions: Evidence from scallops and crinoids, in: Donovan, K., (Ed.), Mass Extinctions:Processes and Evidence, Columbia University Press, New York,pp. 174-194.
64. Kelber, K.-P., 1998. Phytostratigraphische aspekte der makrofloren des süddeutschen Keupers.Documenta Naturae, 117,89-115.
65. Kiessling, W., Aberhan, M., Brenneis, B., Wagner, P. J., 2007.Extinction trajectories of benthic organisms across the Triassic-Jurassic boundary.Palaeogeography,Palaeoclimatology,Palaeoecology, 244, 201-222.
66. Klein, H., Lucas, S. G., 2010. The Triassic footprint record of crocodylomorphs — A critical re-evaluation.New Mexico Museum of Natural History and Science Bulletin, 51, 55-60.
67. Klein, H., Lucas, S. G., 2013. The Late Triassic tetrapod ichnotaxonApatopus lineatus(Bock 1952) and its distribution.New Mexico Museum of Natural History and Science Bulletin,61, 313-324.
68. Knoll, A. H., 1984. Patterns of extinction in the fossil record of vascular plants, in: Nitecki, M., (Ed.), Extinction. University of Chicago Press, Chicago, pp. 21-68.
69. Kozur, H. W., Bachmann, G. H., 2005. Correlation of the Germanic Triassic with the international scale.Albertiana, 32,21-35.
70. Kozur, H. W., Bachmann, G. H., 2008. Updated correlation of the Germanic Triassic with the Tethyan scale and assigned numeric ages.Berichte Geologische Bundesanstalt, 76, 53-58.
71. Kozur, H. W., Weems, R. E., 2005. Conchostracan evidence for a late Rhaetian to early Hettangian age for the CAMP volcanic event in the Newark Supergroup, and a Sevatian (late Norian) age for the immediately underlying beds.Hallesches Jahrbuch Geowissenschaft, B27, 21-51.
72. Kozur, H. W., Weems, R. E., 2007. Upper Triassic conchostracan biostratigraphy of the continental rift basins of eastern North America: Its importance for correlating Newark Supergroup events with the Germanic Basin and the international geologic time scale.New Mexico Museum of Natural History and Science Bulletin, 41, 137-188.
73. Kozur, H. W., Weems, R. E., 2010. The biostratigraphic importance of conchostracans in the continental Triassic of the northern hemisphere, in: Lucas, S. G., (Ed.), The Triassic timescale.Geological Society,London,Special Publications,334, 315-417.
74. Krystyn, L., Boquerel, H., Kuerschner, W., Richoz, S., Gallet,Y., 2007. Proposal for a candidate GSSP for the base of the Rhaetian Stage.New Mexico Museum of Natural History and Science Bulletin, 41, 189-199.
75. Kuerschner, W. M., Bonis, N. R., Krystyn, L., 2007. Carbonisotope stratigraphy and palynostratigraphy of the Triassic-Jurassic transition in the Tiefengraben section — Northern Calcareous Alps (Austria).Palaeogeography,Palaeoclimatol‑ogy,Palaeoecology, 244, 257-280.
76. Kürschner, W. M., Herngreen, G. F. W., 2010. Triassic palynology of central and northwestern Europe: A review of palynofloral diversity patterns and biostratigraphic subdivisions, in:Lucas, S. G., (Ed.), The Triassic timescale.Geological Society,London,Special Publications, 334, 263-283.
77. Labandeira, C. C., Sepkoski, J. J., Jr., 1993. Insect diversity in the fossil record.Science, 261, 310-315.
78. Langer, M. C., 2014. The origins of Dinosauria: Much ado about nothing.Palaeontology, 2014, 1-10.
79. Langer, M. C., Ezcurra, M. D., Bittencourt, J. S., Novas, F. E.,2009. The origin and early evolution of dinosaurs.Biological Reviews, 84, 1-56.
80. Long, R. A., Murry, P. A., 1995, Late Triassic (Carnian and Norian) tetrapods from the southwestern United States.New Mexico Museum of Natural History and Science Bulletin, 4,1-254.
81. Lucas, S. G., 1994. Triassic tetrapod extinctions and the compiled correlation effect.Canadian Society of Petroleum Ge‑
ologists Memoir, 17, 869-875.
82. Lucas, S. G., 1998. Global Triassic tetrapod biostratigraphy and biochronology.Palaeogeography,Palaeoclimatology,Pal‑aeoecology, 143, 347-384.
83. Lucas, S. G., 2007. Tetrapod footprint biostratigraphy and biochronology.Ichnos, 14, 5-38.
84. Lucas, S. G., 2008. Global Jurassic tetrapod biochronology.VoluminaJurassica, 6, 99-108.
85. Lucas, S. G., 2010a. The Triassic timescale: An introduction,in: Lucas, S. G., (Ed.), The Triassic timescale.Geological So‑ciety,London,Special Publications, 334, 1-16.
86. Lucas, S. G., 2010b. The Triassic timescale based on nonmarine tetrapod biostratigraphy and biochronology, in: Lucas, S.G., (Ed.), The Triassic timescale.Geological Society,London,Special Publications, 334, 447-500.
87. Lucas, S. G., 2013a. Plant megafossil biostratigraphy and biochronology, Upper Triassic Chinle Group, western USA.New Mexico Museum of Natural History and Science Bulletin, 61,354-365.
88. Lucas, S. G., 2013b. A new Triassic timescale.New Mexico Mu‑seum of Natural History and Science Bulletin, 61, 366-374.
89. Lucas, S. G., 2015. Age and correlation of Late Triassic tetrapods from southern Poland.Annales Societatis Geologorum Poloniae, in press. doi.org/10.14241/asgp.2015.024.
90. Lucas, S. G., Heckert, A. B., 2011. Late Triassic aetosaurs as the trackmaker of the tetrapod footprint ichnotaxonBrachy‑chirotherium.Ichnos, 18, 197-208.
91. Lucas, S. G., Huber, P., 2003. Vertebrate biostratigraphy and biochronology of the nonmarine Late Triassic, in: LeTourneau,P. M., Olsen, P. E., (Eds.), The great rift valleys of Pangea in eastern North America. Volume 2. Sedimentology, Stratigraphy, and Paleontology. Columbia University Press, New York,pp. 143-191.
92. Lucas, S. G., Hunt, A. P., 1994. The chronology and paleobiogeography of mammalian origins, in: Fraser, N. C., Sues,H-D.,(Eds.), In the Shadow of the Dinosaurs. Cambridge University Press, Cambridge, pp. 335-351.
93. Lucas, S. G., Tanner, L. H., 2004. Late Triassic extinction events.Albertiana, 31, 31-40.
94. Lucas, S. G., Tanner, L. H., 2007a. Tetrapod biostratigraphy and biochronology of the Triassic-Jurassic transition on the southern Colorado Plateau, USA.Palaeogeography,Palaeocli‑matology,Palaeoecology, 244, 242-256.
95. Lucas, S. G., Tanner, L. H., 2007b. The nonmarine Triassic-Jurassic boundary in the Newark Supergroup of eastern North America.Earth Science Reviews, 84, 1-20.
96. Lucas, S. G., Tanner, L. H., 2008. Reexamination of the end-Triassic mass extinction, in: Elewa, A. M. T., (Ed.), Mass Extinction. Springer Verlag, New York, pp. 66-103.
97. Lucas, S. G., Wild, R., 1995. A Middle Triassic dicynodont from Germany and the biochronology of Triassic dicynodonts.Stuttgarter Beiträge zur Naturkunde, 220, 1-16.
98. Lucas, S. G., Heckert, A. B., Hotton, N. III., 2002. The rhynchosaurHyperodapedonfrom the Upper Triassic of Wyoming and its global biochronological significance.New Mexico Mu‑seum of Natural History and Science Bulletin, 21, 149-156.
99. Lucas, S. G., Lockley, M. G., Hunt, A. P., Milner, A. R. C., Tanner, L. H., 2006a. Tetrapod footprint biostratigraphy of the Triassic-Jurassic transition in the American Southwest.New Mexico Museum of Natural History and Science Bulletin, 37,105-108.
100. Lucas, S. G., Klein, H., Lockley, M. G., Spielmann, J. A., Gierlinski, G., Hunt, A. P., Tanner, L. H., 2006b. Triassic-Jurassic stratigraphic distribution of the theropod footprint ichnogenusEubrontes.New Mexico Museum of Natural History and Science Bulletin, 37, 86-93.
101. Lucas, S. G., Hunt, A. P., Heckert, A. B., Spielmann, J. A.,2007a. Global Triassic tetrapod biostratigraphy and biochronology: 2007 status.New Mexico Museum of Natural History and Science Bulletin, 41, 229-240.
102. Lucas, S. G., Taylor, D. G., Guex, J., Tanner, L. H., Krainer,K., 2007b. The proposed global stratotype section and point for the base of the Jurassic System in the New York Canyon area, Nevada, USA.New Mexico Museum of Natural History and Science Bulletin, 40, 139-168.
103. Lucas, S. G., Tanner, L. H., Donohoo-Hurley, L. L., Geissman,J. W., Kozur, H. W., Heckert, A. B., Weems, R. E., 2011. Position of the Triassic-Jurassic boundary and timing of the end-Triassic extinctions on land: Data from the Moenave Formation on the southern Colorado Plateau, USA.Palaeogeog‑raphy,Palaeoecology,Palaeoclimatology, 302, 194-205.
104. Lucas, S. G., Tanner, L. H., Kozur, H. W., Weems, R. E., Heckert, A. B., 2012. The late Triassic timescale: Age and correlation of the Carnian-Norian boundary.Earth Science Reviews,114, 1-8.
105. Lucas, S. G., Szajna, M. J., Lockley, M. G., Fillmore, D. L.,Simpson, E. L., Klein, H. Boyland, J., Hartline, B. W., 2014.The Middle-Late Triassic tetrapod footprint ichnogenusGw‑ynnedichnium.New Mexico Museum of Natural History and Science Bulletin, 62, 135-156.
106. Maisch, M. W., Kapitzke, M., 2010. A presumably marine phytosaur (Reptilia: Archosauria) from the pre-planorbis beds(Hettangian) of England.Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 257, 373-379.
107. Mander, L., Kürschner, W. M., McElwain, J. C., 2010. An explanation for conflicting records of Triassic-Jurassic plant diversity.Proceedings of the National Academy of Science USA,107, 15351-15356.
108. Mander, L., Wesseln, C. J., McElwain, J. C., Punyasena, S. W.,2012. Tracking taphonomic regimes using chemical and mechanical damage of pollen and spores: An example from the Triassic-Jurassic mass extinction.PLOS ONE, 7(11), e49153.
109. Mander, L., Kürschner, W. M., McElwain, J. C., 2013. Palynostratigraphy and vegetation history of the Triassic-Jurassic transition in East Greenland.Journal of the Geological Society London, 170, 37-46.
110. Marzoli, A., Renne, P. R., Piccirillo, E. M., Ernesto, M., Bellieni, G., DeMin, A., 1999. Extensive 200-million-year-old continental flood basalts of the central Atlantic Magmatic province.Science, 284, 616-618.
111. Marzoli, A., Bertrand, H., Knight, K. B., Cirilli, S., Buratti,N., Verati, C., Nomade, S., Renne, P. R., Youbi, N., Martini,R., Allenbach, N., Neuwerth, R., Rapaille, C., Zaninetti, L.,Zaninetti, L., Bellieni, G., 2004. Synchrony of the Central Atlantic magmatic province and the Triassic-Jurassic boundary climate and biotic crisis.GSA Bulletin, 32, 973-976.
112. Marzoli, A., Jourdan, F. Puffer, J. H., Cuppone, T., Tanner,L. H., Weems, R. E., Bertrand, H., Cirilli, S., Bellieni, G.,De Min, A., 2011. Timing and duration of the Central Atlantic magmatic province in the Newark and Culpeper basins, eastern U.S.A.Lithos, 122, 175-188.
113. McCune, A. R., Schaeffer, B., 1986. Triassic and Jurassic fishes: Patterns of diversity, in: Padian, K., (Ed.), The Beginning of the Age of Dinosaurs. Cambridge University Press, Cambridge, pp. 171-181.
114. McElwain, J. C., Punyasena, S. W., 2007. Mass extinction events and the plant fossil record.Trends in Ecology and Evo‑lution, 22, 548-557.
115. McElwain, J. C., Beerling, D. J., Woodward, F. I., 1999. Fossil plants and global warming at the Triassic-Jurassic boundary.Science, 285, 1386-1390.
116. McElwain, J. C., Popa, M. E., Hesselbo, S. P., Haworth, M.,Surlyk, F., 2007. Macroecological responses of terrestrial vegetation to climatic and atmospheric change across the Triassic/Jurassic boundary in East Greenland.Paleobiology, 33,547-573.
117. McElwain , J. C., Wagner, P. J., Hesselbo, S. P., 2009. Fossil plant relative abundances indicate sudden loss of late Triassic biodiversity in East Greenland.Science, 324, 1554-1556.
118. McGhee, G. R., Jr., Sheehan, P. M., Bottjer, D. J., Droser,M. L., 2004. Ecological ranking of Phanerozoic biodiversity crises: Ecological and taxonomic severities are decoupled.Palaeogeography,Palaeoclimatology,Palaeoecology, 211,289-297.
119. McGhee, G. R., Jr., Clapham, M. E., Sheehan, P. M., Bottjer,D. J., Droser, M. L., 2013. A new ecological-severity ranking of major Phanerozoic biodiversity crises.Palaeogeography,Palaeoclimatology,Palaeoecology, 370, 260-270.
120. McHone, J. G., Puffer, J. H., 1996. Hettangian flood basalts across the Pangaean rift. Connecticut State Geological and Natural History Survey Natural Resources Center Miscellaneous Report, 1, 29.
121. Milner, A. R., 1993. Amphibian-grade Tetrapoda, in: Benton,M. J., (Ed.), The Fossil Record 2. Chapman and Hall, London,pp. 665-679.
122. Milner, A. R., 1994. Late Triassic and Jurassic amphibians: Fossil record and phylogeny, in: Fraser, N. C., Sues, H-D., (Eds.),In the Shadow of the Dinosaurs. Cambridge University Press,Cambridge, pp. 5-22.
123. Milner, A. R. C., Kirkland, J. I., Birthisel, T. A., 2006. The geographic distribution and biostratigraphy of Late Triassic-Early Jurassic freshwater fish faunas of the western United States.New Mexico Museum of Natural History and Science Bulletin,37, 522-529.
124. Nesbitt, S. J., 2011. The early evolution of archosaurs: Relationships and the origin of major clades.Bulletin of the American Museum of Natural History, 352, 1-292.
125. Nesbitt, S. J., Brusatte, S. L., Desojo, J. B., Liparini, A., De Franca, M. A. G., Weinbaum, J. C., Gower, D. J., 2013. Rauisuchia, in: Nesbitt, S. J., Desojo, J. B., Irmis, R. B., (Eds.),Anatomy, phylogeny and palaeobiology of early archosaurs and their kin.Geological Society,London,Special Publica‑tions, 379, 241-274.
126. Newham, E., Benson, R., Upchurch, P., Goswani, A., 2014.Mesozoic mammaliaform diversity: The effect of sampling corrections on reconstructions of evolutionary dynamics.Pal‑aeogeography,Palaeoclimatology,Palaeoecology, 412, 32-44.
127. Niedźwiedzki, G., 2011. A Late Triassic dinosaur-dominated ichnofauna from the Tomanová Formation of the Tatra Mountains, central Europe.Acta Palaeontologica Polonica, 56,291-300.
128. Niklas, K. J., Tiffney, B. H., Knoll, A. H., 1983. Patterns in vascular land plant diversification: A statistical analysis at the species level.Nature, 303, 614-616.
129. Ogg, J. G., 2012a. Triassic. In: Gradstein, F. M., Ogg, J. G.,Schmitz, M. D., Ogg, G. M., (Eds.), The Geologic Timescale 2012. volume 2. Elsevier, Amsterdam, pp. 681-730.
130. Ogg, J. G., 2012b. Jurassic, in: Gradstein, F. M., Ogg, J. G.,Schmitz, M. D., Ogg, G. M., (Eds.), The Geologic Timescale 2012. volume 2. Elsevier, Amsterdam, pp. 731-791.
131. Ogg, J. G., Huang, C., Hinnov, L., 2014. Triassic timescale status: A brief overview.Albertiana, 41, 3-30.
132. Olsen, P. E., Galton, P. M., 1984. A review of the reptile and amphibian assemblages from the Stormberg of South Africa,with special emphasis on the footprints and the age of the Stormberg.PalaeontologicaAfricana, 25, 87-110.
133. Olsen, P. E., Rainforth, E. C., 2003. The Early Jurassic ornithischian dinosaur ichnogenusAnomoepus, in: LeTourneau,P. M., Olsen, P. E., (Eds.), The Great Rift Valleys of Pangea in Eastern North America. volume 2. Sedimentology, Stratigraphy, and Paleontology. Columbia University Press, New York,pp. 314-368.
134. Olsen, P. E., Sues, H-D., 1986. Correlation of continental Late Triassic and Early Jurassic sediments, and patterns of the Triassic-Jurassic tetrapod transition, in: Padian, K., (Ed.),The Beginning of the Age of Dinosaurs. Cambridge University Press, Cambridge, pp. 321-351.
135. Olsen, P. E., Shubin, N. H., Anders, M. H., 1987. New Early Jurassic tetrapod assemblages constrain Triassic-Jurassic tetrapod extinction event.Science, 237, 1025-1029.
136. Olsen, P. E., Fowell, S. J., Cornet, B., 1990. The Triassic/Jurassic boundary in continental rocks of eastern North America; a progress report.Geological Society of America Special Paper, 247, 585-593.
137. Olsen, P. E., Schlische, R. W., Fedosh, M. S., 1996. 580 ky duration of the Early Jurassic flood basalt event in eastern North America estimated using Milankovitch cyclostratigraphy.Mu‑seum of Northern Arizona Bulletin, 60, 11-22.
138. Olsen, P. E., Smith, J. B., McDonald, N. G., 1998. Type material of the type species of the classic theropod footprint generaEubrontes,Anchisauripus, andGrallator(Early Jurassic, Hartford and Deerfield basins, Connecticut and Massachusetts, U.S.A.).Journal of Vertebrate Paleontology, 18, 586-601.
139. Olsen, P. E., Kent, D. V., Sues, H. D., Koeberl, C., Huber, H.,Montanari, A., Rainforth, E. C., Powell, S. J., Szajna, M. J.,Hartline, B. W., 2002a. Ascent of dinosaurs linked to an iridium anomaly at the Triassic-Jurassic boundary.Science, 296,1305-1307.
140. Olsen, P. E., Koeberl, C., Huber, H., Montanari, A., Fowell,S. J., Et-Touhani, M., Kent, D. V., 2002b. The continental Triassic-Jurassic boundary in central Pangea: Recent progress and preliminary report of an Ir anomaly.Geological Society of America Special Paper, 356, 505-522.
141. Olsen, P. E., Kent, D. V., Whiteside, J. H., 2011. Implications of the Newark Supergroup-based astrochronology and geomagnetic polarity time scale (Newark-APTS) for the tempo and mode of the early diversification of the Dinosauria.Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 101, 201-229.
142. Olsen, P. E., Reid, J. C., Taylor, K. B., Whiteside, J. H., Kent,D. V., 2015. Revised stratigraphy of Late Triassic age strata of the Dan River basin (Virginia and North Carolina, USA) based on drill core and outcrop data.Southeastern Geology, 51,1-31.
143. Onoue, T., Sato, H., Nakamura, T., Noguchi, T., Hidaka, Y.,Shirai, N., Ebihara, M., Osawa, T., Hatsukawa, Y., Toh, Y.,Koizumi, M., Harada, H., Orchard, M. J., Nedachi, M., 2012.Deep-sea record of impact apparently unrelated to mass extinction in the Late Triassic. Proceedings of the National Academy of Science, 109, 19134-19139.
144. Orbell, G., 1973. Palynology of the British Rhaeto-Liassic.Bul‑letin of the Geological Society of Great Britain, 44, 1-44.
145. Orchard, M. J., 2010. Triassic conodonts and their role in stage boundary definition, in: Lucas, S. G., (Ed.), The Triassic timescale.Geological Society,London,Special Publications,334, 139-161.
146. Orchard, M. J., 2013. Five new genera of conodonts from the Carnian-Norian boundary beds of Black Bear Ridge, northeast British Columbia, Canada.New Mexico Museum of Natural History and Science Bulletin, 61, 445-457.
147. Orchard, M. J., 2014.Conodonts from the Carnian-Norian boundary (Upper Triassic) of Black Bear Ridge, northeastern British Columbia, Canada.New Mexico Museum of Natural History and Science Bulletin, 64, 1-139.
148. Pedersen, K. R., Lund, J. J., 1980. Palynology of the plantbearing Rhaetian to Hettangian Kap Stewart Formation,Scoresby Sund, East Greenland.Review of Palaeobotany and Palynology, 31, 1-69.
149. Pieńkowski, G., Niedźwiedzki, G., Branski, P., 2014. Climatic reversals related to the Central Atl—antic magmatic province caued the end-Triassic biotic crisis Evidence from continental strata in Poland.Geological Society of America Special Paper, 505, 263-286.
150. Pieńkowski, G., Niedźwiedzki, G., Waksmundzka, M., 2011.Sedimentological, palynological and geochemical studies of the terrestrial Triassic-Jurassic boundary in northwestern Poland.Geological Magazine, 149, 308-332.
151. Pott, C., McLoughlin, S., 2009. Bennettitalean foliage in the Rhaetian-Bajocian (latest Triassic-Middle Jurassic) floras of Scania, southern Sweden.Review of Palaeobotany and Pa‑lynology, 158, 117-166.
152. Rainforth, E. C., 2003. Revision and re-evaluation of the Early Jurassic dinosaurian ichnogenusOtozoum.Palaeontology, 46,803-838.
153. Ramezani, J., Bowring, S. A., Pringle, M., Winslow III, F. D.,Rasbury, E. T., 2005. The Manicouagan impact melt rock: A proposed standard for intercalibration of U-Pb and 40Ar/39/Ar isotopic systems. 15thV.M. Goldschmidt Conference Abstract Volume, A321.
154. Rigo, M., Preto, N., Roghi, G., Tateo, F., Mietto, P., 2007. A rise in the carbonate compensation depth of western Tethys in the Carnian (Late Triassic): Deep-water evidence for the Carnian pluvial event.Palaeogeography,Palaeoclimatology,Palaeoecology, 246, 188-205.
155. Ruckwied, K., Götz, A. E., Pálfy, J., Török, Á., 2008. Palynology of a terrestrial coal-bearing series across the Triassic/Jurassic boundary (Mecsek Mts, Hungary).Central European Geology, 51, 1-15.
156. Ruhl, M., Veld, H., Kuerschner, W. M., 2010. Sedimentary organic matter characterization of the Triassic-Jurassic boundary GSSP at Kuhjoch (Austria).Earth and Planetary Science Letters, 292, 17-26.
157. Schäfer, P., Fois, E., 1987. Systematics and evolution of Triassic Bryozoa.Geologica et Palaeontologica, 21, 173-225.
158. Schaller, E. M., Wright, J. D., Kent, D. V., Olsen, P. E., 2012.Rapid emplacement of the Central Atlantic Magmatic Province as net sink for CO2.Earth and Planetary Science Letters,323-324, 27-39.
159. Schoch, R. R., Milner, A. R., 2000. Stereospondyli.Encyclope‑dia of Paleoherpetology, 3B, 1-203.
160. Schoene, B., Guex, J., Bartolini, A., Schaltegger, U., Blackburn, T. J., 2010, Correlating the end-Triassic mass extinction and flood basalt volcanism at the 100 ka level.Geology, 38,387-390.
161. van de Schootbrugge, B., Quan, T., Lindström, S., Püttmann,W., Heunisch, C., Pross, J., Fiebig, J., Petschick, R., Röhling,H-G., Richoz, S., Rosenthal, Y., Falkowski, P. G., 2009. Floral changes across the Triassic/Jurassic boundary linked to flood basalt volcanism.Nature Geoscience, 2, 589-594.
162. Schuurman, W. M. L., 1979. Aspects of Late Triassic palynology. 3. Palynology of latest Triassic and earliest Jurassic deposits of the northern limestone Alps in Austria and southern Germany, with special reference to a palynological characterization of the Rhaetian stage in Europe.Review of Palaeo‑botany and Palynology, 27, 53-75.
163. Sepkoski, J. J., Jr., 1982. Mass extinctions in the Phanerozoic oceans: A review.Geological Society of America Special Pa‑per, 190, 283-289.
164. Sha, J., Vajda, V., Pan, Y., Larsson, L., Yao, X., Zhang, X.,Wang, Y., Cheng, X., Jiang, B., Deng, S., Chen, S., Peng, B.,2011. Stratigraphy of the Triassic-Jurassic boundary successions of the southern margin of the Junggar basin, northwestern China.Acta Geologica Sinica, 85, 421-436.
165. Sha, J., Olsen, P. E., Pan, Y., Xu, D., Wang, Y., Zhang, X.,Yao, X., Vajda, V., 2015. Triassic-Jurassic climate in continental high-latitude Asia was dominated by obliquity-paced variations (Junggar Basin, Ürümqi, China). Proceedings of the National Academy of Sciences, 112, 3624-3629.
166. Shubin, N. H., Olsen, P. E., Sues, H.-D., 1994. Early Jurassic small tetrapods from the McCoy Brook Formation of Nova Scotia, Canada, in: Fraser, N. C., Sues, H-D., (Eds.), In the Shadow of the Dinosaurs. Cambridge University Press, Cambridge, pp. 242-250.
167. Sigurdsson, H., 1990. Assessment of atmospheric impact of volcanic eruptions.Geological Society of America Special Pa‑per, 247, 99-110.
168. Silvestri, S. M., Szajna, M. J., 1993. Biostratigraphy of vertebrate footprints in the Late Triassic section of the Newark Basin, Pennsylvania: Reassessment of stratigraphic ranges.New Mexico Museum of Natural History and Science Bulletin, 3,439-445.
169. Simms, M. J., Ruffell, A. H., 1989. Synchroneity of climatic change and extinctions in the Late Triassic.Geology, 17, 265-268.
170. Simms, M. J., Ruffell, A. H., 1990. Climatic and biotic change in the Late Triassic.Journal Geological Society(London), 147,321-327.
171. Smith, R. M. H., Marsicano, C. A., Wilson, J. A., 2009. Sedimentology and paleoecology of a diverse Early Jurassic tetrapod tracksite in Lesotho, southern Africa.Palaios, 24, 672-684.
172. Spielmann, J. A., Lucas, S. G., Hunt, A. P., 2013. The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A.New Mexico Museum of Natural History and Science Bulletin, 61, 562-566.
173. Steinthorsdottir, M., Jeram, A. J., McElwain, J. C., 2011. Extremely elevated CO2concentrations at the Triassic/Jurassic boundary.Palaeogeography,Palaeoclimatology,Palaeoecol‑ogy, 308, 418-432.
174. Stocker, M. R., Butler, R. J., 2013. Phytosauria, in: Nesbitt,S. J., Desojo, J. B., Irmis, R. B., (Eds.), Anatomy, phylogeny and palaeobiology of early archosaurs and their kin.Geologi‑cal Society,London,SpecialPublications, 379, 91-118.
175. Sues, H-D., Olsen, P. E., 2015. Stratigraphic and temporal context and faunal diversity of Permian-Jurassic continental tetrapod assemblages from the Fundy rift basin, eastern Canada.Atlantic Geology, 51, 139-205.
176. Szajna, M. J., Silvestri, S. M., 1996. A new occurrence of the ichnogenusBrachychirotherium: Implications for the Triassic-Jurassic mass extinction event.Museum of Northern Ari‑zona Bulletin, 60, 275-283.
177. Szulc, J., Racki, G., Środoń, J., Lucas, S. G., 2015. Norian age of the Lipie Śląskie vertebrate fauna explains its alleged oddity.Acta Palaeontologica Polonica, in press.
178. Tanner, L. H., Hubert, J. F., Coffey, B. P., McInerney, D. P.,2001. Stability of atmospheric CO2levels across the Triassic/Jurassic boundary.Nature, 411, 675-677.
179. Tanner, L. H., Lucas, S. G., Chapman, M. G., 2004. Assessing the record and causes of Late Triassic extinctions.Earth-ScienceReviews, 65, 103-139.
180. Tanner, L. H., Smith, D. L., Allan, A., 2007. Stomatal response of swordfern to volcanogenic CO2and SO2from Kilauea volcano, Hawaii.Geophysical Research Letters, 34, L15807,http://dx.doi.org/10.1029/2007GL030320.
181. Thulborn, T., Turner, S., 2003. The last dicynodont: An Australian Cretaceous relict. Proceedings Royal Society London B, 270, 985-993.
182. Traverse, A., 1988. Plant evolution dances to a different beat.Historical Biology, 1, 277-301.
183. de Valais, S., 2009. Ichnotaxonomic revision ofAmeghinich‑nus, a mammalian ichnogenus from the Middle Jurassic La Matilde Formation, Santa Cruz Province, Argentina.Zootaxa,2203, 1-21.
184. Visscher, H., Van Houte, M., Brugman, W. A., Poort, R. J.,1994. Rejection of a Carnian (late Triassic) “pluvial event”in Europe.Review of Palaeobotany and Palynology, 83, 217-226.
185. Weems, R. E., 1992. The “terminal Triassic catastrophic extinction event” in perspective: A review of Carboniferous through Early Jurassic terrestrial vertebrate extinction patterns.Palaeogeography,Palaeoclimatology,Palaeoecology,94, 1-29.
186. Weems, R. E., Lucas, S. G., 2015. A revision of the Norian conchostracan zonation in North America and its implications for Late Triassic North American tectonic history.New Mexico Museum of Natural History and Science Bulletin, 67, 303-317.
187. Whiteside, J. H., Olsen, P. E., Kent, D. V., Fowell, S. J., Et-Touhami, M., 2007. Synchrony between the Central Atlantic magmatic province and the Triassic-Jurassic mass-extinction event?Palaeogeography,Palaeoclimatology,Palaeoecology,244, 345-367.
188. Whiteside, J. H., Olsen, P. E., Eglinton, T., Brookfield, M. E.,Sambrotto, R. N., 2010. Compound-specific carbon isotopes from Earth’s largest flood basalt eruptions directly linked to the end-Triassic mass extinction. Proceedings National Academy of Science USA, 107, 6721-6725.
189. Wotzlaw, J-F., Guex, J., Bartolini, A., Gallet, Y., Krystyn, L.,McRoberts, C. A., Taylor, D., Schoene, B., Schaltegger, U.,2014. Towards accurate numerical calibration of the Late Triassic: High-precision U-Pb geochronology constraints on the duration of the Rhaetian.Geology, 42, 571-574.
190. Yapp, C. J., Poths, H., 1996. Carbon isotopes in continental weathering environments and variations in ancient atmospheric CO2pressure.Earth and Planetary Science Letters,137, 71-82.
猜你喜欢
小学生学习指导(低年级)(2022年5期)2022-05-31
党员生活(2022年4期)2022-05-09
数学小灵通(1-2年级)(2020年9期)2020-10-27
VOGUE服饰与美容(2020年12期)2020-02-24
东坡赤壁诗词(2019年5期)2019-11-14
意林(2019年17期)2019-10-07
意林(2019年15期)2019-09-03
Coco薇(2017年5期)2017-06-05
中国经济周刊(2016年13期)2016-07-08
人生十六七(2015年2期)2015-02-28
 Journal of Palaeogeography2015年4期
Journal of Palaeogeography2015年4期
- Journal of Palaeogeography的其它文章
- Correction of two Upper Paleozoic stratigraphic units in the Tianshan Mountains region, Xinjiang Uygur Autonomous Region and implications on the Late Paleozoic evolution of Tianshan tectonic complex,Northwest China
- Modeling the Middle Jurassic ocean circulation
- Lithofacies palaeogeography of the Carboniferous and Permian in the Qinshui Basin, Shanxi Province, China