
伞形科藁本属20种植物的果实特征及其分类学意义
2015-07-04 09:22陈俊佩马祥光何兴金
西北植物学报 2015年8期
陈俊佩,刘 爽,马祥光,何兴金
(四川大学 生命科学学院,生物资源与生态环境教育部重点实验室,成都610064)
藁本属(LigusticumL.)是伞形科(Apiaceae)芹亚科(Apioideae)中一个重要的分类群[1]。全世界约有60 余种,主要分布在欧洲、亚洲和北美洲等北温带地区。中国现有40余种(35种为特有种),主要分布于西南地区高山及亚高山地带,少数种类分布于黄河流域以北山区[2]。
藁本属的植物由于形态多样,营养器官分化较大,分生果的形状介于两侧扁压与背腹扁压之间,使该属与其邻近属如亮蛇床属(SelinumL.)、山芎属(ConioselinumFisch.ex Hoffm.)、羌活属(No-topterygium de Boiss.)、岩茴香属(TilingiaRegel&Tiling)和拟藁本属(LigusticopsisLeute)等属间的分类界限不清,种的出入频繁[2-4],而争议比较大的是藁本属与拟藁本属的属间界限和系统定位[5-8]。其中拟藁本属是Leute在1969年将藁本属中萼齿发育较好的类群分出所成立的一个属,包含14 个种[8]。实际上在Leute所述的藁本属中,羽苞藁本[L.daucoides(Franch.)Franch.]和多苞藁本(L.involucratumFranch.)等也具有明显的萼齿,因此Leute所成立的拟藁本属与藁本属之间缺乏明显的分属界限,难以区别开来,而拟藁本属也未被其他学者所采纳。
近年来,国内外学者在植物化学[9]、孢粉学[10-12]、细胞学[13-16]、叶表皮微形态[17]、叶柄解剖[18]、分支分类学[19]和分子系统学[20-27]等方面对藁本属进行了相关研究。在伞形科传统的经典分类学研究中,果实由于其形态和结构的独特性和稳定性,一直被分类学家视为分类和鉴定的重要依据[1,28]。有关藁本属果实特征的研究,主要集中在果实解剖方面,如国外学者Pimenov M.G.等对藁本属部分种类进行了研究,并对一些种的分类系统进行调整[29-31],将尖瓣藁本(L.weberbauerianumFedde ex H.Wolff)等归入羌活属,而在之后对山芎属的修订中,根据果实中维管束的位置等特征将藁本属中辽藁本[L.jeholense(Nakai &Kitagawa)Nakai &Kitagawa]和尖叶藁本(L.acuminatumFranch.)等8个种转隶于山芎属中;国内,虽然孟德玉对藁本属5种植物果实进行对比观察,但涉及到的种类较少,缺乏系统的解剖学研究[32]。在果实微形态的研究方面,前人对伞形科其他类群的研究结果表明,果实微形态特征在属下的分类中具有重要意义,而对藁本属果实表面微形态的研究尚未见报道[33-37]。
由于藁本属植物大部分生长于高海拔地区,且果实成熟期不同,在野外收集到成熟果实较为困难。通过近四年的材料积累,我们共获得中国藁本属20种及山芎属1种的成熟果实。本文利用扫描电子显微镜对这21种果实的表面微形态进行了首次观察研究,并对其中15种果实进行解剖观察,了解它们的果实微形态学及比较解剖学结构特征,旨在为藁本属的系统分类和分类群的鉴定提供果实方面的依据。
1 材料和方法
1.1 材 料
本文所研究的中国藁本属植物20 种(其中11种曾被Leute归入到拟藁本属中,表中标*)及山芎属1种的果实均为成熟果实,取自野生植株或腊叶标本。凭证标本保存于四川大学标本馆(馆代码为SZ),具体材料来源见表1。
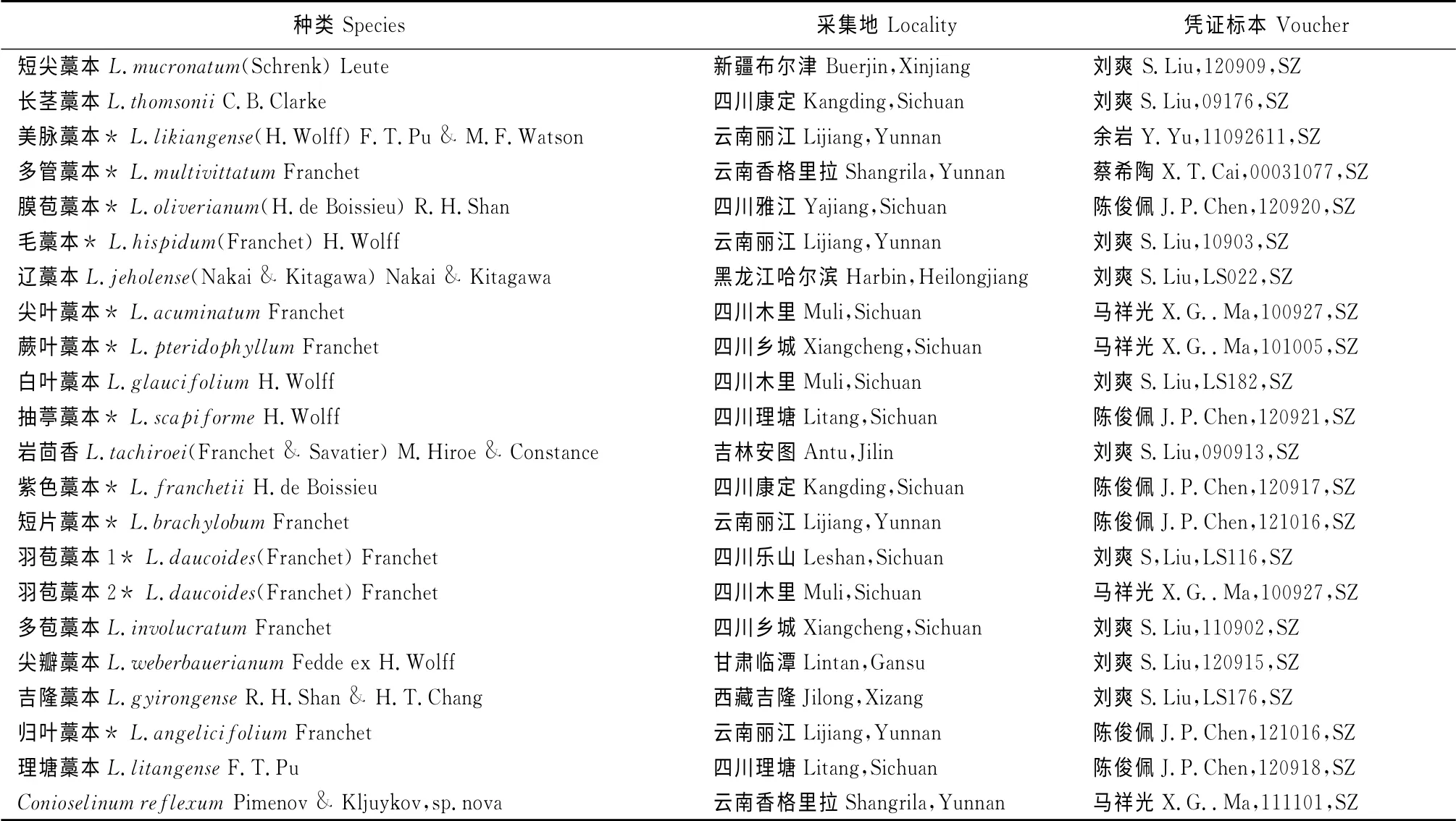
表1 材料来源Table1 Source of materials
1.2 方法
1.2.1 果实形态特征观察 Olympus SZX7解剖镜下检查每份样品5粒以上,选取其中有代表性的果实5~7粒用于实验。在解剖镜下观察果实的形状、颜色和大小等特征,并进行照相、测量和记录。果实形态术语参考刘长江的种子形态描述及相关文献[32-42]。
1.2.2 果实表面微形态特征观察 将果实放入盛有酒精的玻璃小瓶中,超声波震荡清洗2min,取出自然晾干,然后用导电胶将果实粘贴在扫描载物台上进行喷金,之后移入JSM-7500F型扫描电子显微镜仪下观察表面微形态特征并照相,拍照的部位为果实两个背棱槽其中的一个。
1.2.3 果实解剖特征观察 干燥的果实温水浸泡1周,后再经过酒精甘油混合液(体积比例为1∶1)软化处理。采用常规石蜡切片法,切片厚度为10~12μm,经过番红-固绿双重染色,中性树胶封片,光学显微镜下观察果实中部横切面并照相。凭证永久装片保存于四川大学生命科学学院植物系统与进化生物学实验室。
2 结果与分析
2.1 果实外部形态
藁本属分生果形状主要为椭圆形、长圆形或卵圆形,近两侧扁压至背腹扁压,果实表面光滑,背棱突起情况为纤维状突起、显著突起或成窄翅状,侧棱常成翅状,棱槽间多有明显的纵条纹。果实的大小一般为(3.2~5.9)×(1.5~4.6)mm(表2),不同的类群差异较大,较小的如美脉藁本大小为(3.2×2.0)mm,较大的如归叶藁本大小达到(5.9×4.6)mm。
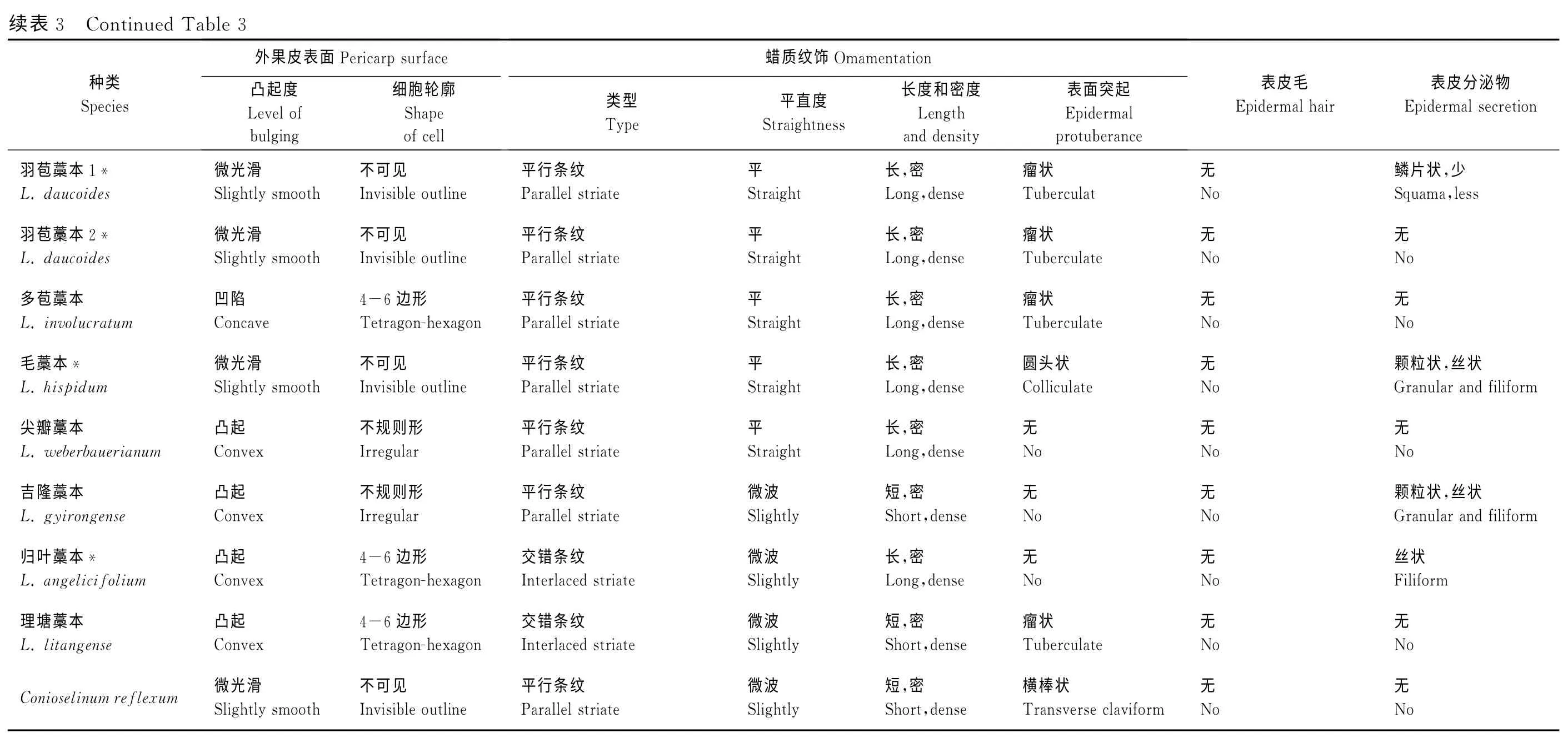
2.2 果实表面微形态特征
从电镜观察结果来看,所研究藁本属类群的外果皮表面微形态特征(表3;图版Ⅰ、Ⅱ)主要为:(1)表面凸起度及细胞轮廓:藁本属植物的外果皮表面凸起度为粗糙、微光滑、凹陷和凸起等4种类型,细胞形状多为不可见,其中紫色藁本、短片藁本和归叶藁本的细胞形状为4~6边形,其他则为隐约可见或不规则形(图版Ⅱ,1~4、17、18)。(2)表面蜡质纹饰:果实表面纹饰类型多样,主要为条状纹饰、网状纹饰和表面突起物3种类型。根据纹饰的排列及式样的不同,条状纹饰又可分为平行条纹、簇状条纹和交错条纹,而表面突起物则表现为瘤状、疣状、横棒状、圆头状和多皱等类型。在所研究的类群中,除尖叶藁本外,其他类群果实表面均具有条状纹饰,而网状纹饰则仅在紫色藁本和短片藁本的果实表面观察到(图版Ⅱ,1~4)。多数类群的果实表面具有2种或2种以上纹饰,如短片藁本具有平行条纹、网状纹饰和瘤状突起物(图版Ⅱ,3、4)。(3)表皮毛:供试的类群中只有短尖藁本和长茎藁本具有表皮毛结构,且表皮毛的类型不同,其中前者为密集的尖囊状和带状表皮毛(图版Ⅰ,1、2),后者仅有密集的尖囊状表皮毛(图版Ⅰ,3、4),同时在表皮毛表面均具有密集排列的微波状粗条纹。(4)表面分泌物:所研究的藁本属20种果实,除蕨叶藁本、白叶藁本和多苞藁本等5种果实表面无分泌物外,其他类群均能观察到,且分泌物的类型及数量在各类群之间区别明显。果实表面的分泌物主要为鳞片状、颗粒状、丝状和絮状4种类型,如短尖藁本、长茎藁本和紫色藁本的表面为排列规则的条带形鳞片状分泌物,且数量上前者明显比后二者少;膜苞藁本的表面为类似碎果皮状的大颗粒状分泌物等(图版Ⅰ,Ⅱ)。
2.3 果实横切面解剖特征
根据果实解剖结构的观察结果,可看到藁本属分生果的横切面观为半长圆形、近半圆形和近五边形,其解剖特征在不同种之间差异明显,而在种内则性状相对稳定,果实基本结构由外到内表现为:
果棱:藁本属分生果一般有5条果棱(尖瓣藁本具有3或4条果棱),每条果棱对应1个维管束,而维管束在果棱中所处的位置在种间有所不同,其中在背棱中常位于果棱的末梢处,在侧棱中则多位于棱基部或中部,少数位于末梢处(图版Ⅲ)。从果棱的发育程度看,藁本属果实的侧棱均为翅状,而背棱则差异较大,可分为3类:突起状,表现为背棱微突起或突起于果实背面(图版Ⅲ,5~10);窄翅状,表现为背棱高出果实背面,但明显较侧棱短(图版Ⅲ,11~16);翅状,表现为背棱与侧棱形状相同,且长度与侧棱近相等(图版Ⅲ,17~19)。
果皮与种皮:外果皮均为1层排列整齐的扁平细胞,外壁突起或平直,成熟时外果皮与中果皮分离或在油管处连接,个别种类如尖瓣藁本和理塘藁本的外果皮具有厚的角质层(图版Ⅲ,17~19)。中果皮有3~5层排列无规则的薄壁细胞,在果实成熟时碎裂或者高度紧缩呈线形不可见,数个油管及维管束排列于其中。内果皮多为1层规则排列的长矩形扁平细胞,通常与中果皮和种皮相连,偶有膨胀分离。多数类群的种皮不可见或为1层排列规则的近方形细胞。
?
油管:藁本属果实背棱槽及侧棱槽内油管数多为2~3,稀为1或4;合生面油管数多为5~6,少数为2、3或4,个别种可达到8。根据棱槽内油管的数目可分为3种类型:单油管型-背棱槽和侧棱槽油管数目均为1,该类型仅包含岩茴香1种(图版Ⅲ,11);少油管型-背棱槽和侧棱槽内油管数目较少,多为2~3(图版Ⅲ,5~10);多油管型-背棱槽和侧棱槽油管数目较多,为3及以上(图版Ⅲ,12~19)。
胚乳:藁本属果实胚乳的横切面形状多样,多为长圆形或近半圆形,如短尖藁本和美脉藁本(图版Ⅲ,5~6);少数为五边形或马蹄形,如岩茴香和理塘藁本(图版Ⅲ,11、19)。胚乳合生面处的的凹陷程度在种间也有一定的差异,如短尖藁本、羽苞藁本和多苞藁本等合生面为平直型(图版Ⅲ,5、9~10),美脉藁本和膜苞藁本为微凸或微凹的近平直型(图版Ⅲ,6~7),而归叶藁本、尖瓣藁本和理塘藁本的合生面则为深内凹型,凹陷程度可达胚乳的1/3~1/2 处(图版Ⅲ,16~19)。
山芎属C.reflexum的果实形态及横切特征为:果实近背腹扁压,背棱呈窄翅状,背棱槽处有3个油管,侧棱槽油管数目为3~5,合生面油管数目为8,胚乳横切面形状为长圆形(图版Ⅲ,20);其果实微形态特征为:外果皮表面微光滑,细胞轮廓不可见,具平行条纹和横棒状突起,无分泌物(图版Ⅱ,19~20)。
3 讨 论
3.1 藁本属果实特征的分类学意义
目前,关于伞形科果实微形态及解剖特征的研究表明,在种内水平上果实微形态及解剖特征具有相当好的稳定性,种间则表现出明显的差异性[32-41]。本文通过扫描电镜及石蜡切片方法对羽苞藁本两个居群的果实进行观察,发现果实微形态及解剖特征在种内具有良好的稳定性,比如:果实表面凸起度、表面纹饰、分生果及胚乳横切面形状、棱槽及合生面油管数目等特征在两个居群间较为稳定,而这些特征在种间则表现出较大的差异性,为藁本属的分类鉴定和种间关系的探讨提供了重要的形态学依据。
通过扫描电镜的对比观察,藁本属不同类群在果实微形态特征方面,除尖叶藁本外,其余种类均具有蜡质条纹,因此,该纹饰特征基本为所研究种类的共性特征。结合已有的研究[35-38],伞形科其他属如独活属(Heracleum)、棱子芹属(Pleurospermum)和前胡属(Peucedanum)等也具有条状纹饰,即果实微形态的蜡质条纹纹饰特征不能作为藁本属与其他属的区别特征。除此之外的其他特征如表面凸起度、细胞轮廓、表面突起物及表皮毛等特征与其他属也存在交叉,并不是藁本属独有的特征。同时该属果实的解剖结构特征形态多样,与其他近缘属的结构特征有一定的相似性,如当归属(Angelica)果实具有背腹扁压情况,羌活属果实具有明显的果棱[39],棱子芹属果实也存在单油管和多油管现象等[41]。因此通过对藁本属果实微形态学及比较解剖学特征的综合分析,从果实微形态特征和比较解剖学物特征角度,其研究结果不支持藁本属为一个自然类群,这也与前人相关的形态与分子等研究的藁本属不为单系群的结果相符合[4,21-24]。
自1754年林奈在其植物属志中建立藁本属以来,其与拟藁本属LigusticopsisLeute的属间界限及系统定位一直存在争议[5-8]。Leute根据藁本属植物中萼齿发育情况,将其中萼齿发育良好的14种移出藁本属,建立了拟藁本属[8],而由于Leute所述的藁本属中多苞藁本等也具有明显的萼齿,因此拟藁本属与藁本属之间缺乏明显的形态间断,即缺乏明显属级分类界定特征,Leute的该分类观点也未被其他学者所认同;王萍莉等通过对21种花粉形态的研究,支持将拟藁本属归入藁本属[10],而《中国植物志》、中国藁本属修订和《Flora of China》等也支持该结论[1-3];孙纳等通过对藁本属分支分类及叶表皮微形态研究,支持拟藁本属归入藁本属中[17,19]。在本次研究的20个藁本属类群中包含Leute所述的拟藁本属11种(表中标*),通过果实表面微形态及解剖结构特征的观察结果可知,拟藁本属与藁本属无法通过果实特征区分开,如膜苞藁本(曾经为拟藁本属)和岩茴香果实表面均微光滑、细胞不可见、具有平行条纹等,羽苞藁本(曾经为拟藁本属)和多苞藁本的果实形状及横切面形状、胚乳形状及果棱凸起度等结构相似,同样蕨叶藁本(曾经为拟藁本属)和白叶藁本解剖结构也相似,因此无法找到有效的衍征将拟藁本属与藁本属明显区别开。相关的分子系统发育的研究中[21-22,24],拟藁本属并未形成一个单系分支,而是分别插入到藁本属的各级分支中。因此,结合前人的研究结果,支持将拟藁本属归入藁本属。
3.2 藁本属一些种的分类问题
Pimenov等在对山芎属的修订中,将藁本属8个种转隶于山芎属中[30],本文包括其中4 种:辽藁本、尖叶藁本、蕨叶藁本和白叶藁本。此外,我们在野外采集到山芎属的Conioselinumreflexum,该种是Pimenov等在同一篇文章中发表的新种,模式标本采集于云南香格里拉[30]。通过对C.reflexum的果实特征进行研究,发现其近背腹扁压、外果皮表面微光滑、细胞轮廓不可见、具平行条纹和横棒状突起、背棱呈窄翅状、棱槽有3~5个油管、合生面油管数目为8、胚乳横切面形状为长圆形等。果实微形态特征方面,尖叶藁本与C.reflexum较为相似:二者均为外果皮表面微光滑、细胞轮廓不可见和具有横棒状突起等,同时二者在解剖结构特征上也具有一定的相似性,不同的是本文中尖叶藁本果实侧棱处维管束位于棱基部,而C.reflexum是在棱末梢处,而这一特征也是Pimenov重建山芎属的重要依据。在相关的分子系统研究中[26,27],通过构建nrDNA ITS及cpDNA 序列进化树发现尖叶藁本与藁本属的羽苞藁本、多苞藁本和抽葶藁本等相距较远,而与山芎属的鞘山芎[Conioselinumvaginatum(Spreng.)Thell.]具有较密切的关系。蕨叶藁本和白叶藁本与C.reflexum具有相似果实解剖结构:背棱均为窄翅状、背棱槽油管数为3、合生面油管较多、维管束位于果棱的末端、胚乳横切面都为长圆形等,同时三者在微形态上也具有一定的相似性。结合植物外部形态[2]、分子系统学[26-27]、果实解剖及果实微形态研究,表明尖叶藁本、蕨叶藁本和白叶藁本与山芎属具有密切的关系,支持将蕨叶藁本和白叶藁本转入山芎属,而尖叶藁本是否转入山芎属需要更多的证据来验证。此外,对于本文研究中涉及到的辽藁本,由于其微形态特征与C.reflexum具有一定的相似性,但尚无法将其与藁本属其他种明显区分开,因此其与山芎属的关系仍需要进一步的研究。
研究的藁本属果实中,尖瓣藁本的果实与其他类群区别明显,主要表现为尖瓣藁本分生果具有3条或4条果棱,而其他类群为5条,并且果棱的形状明显不同。前人对尖瓣藁本的分类有不同的处理意见,其中Pimenov将其归入羌活属[29],而《Flora of China》将其仍处理为藁本属的一种[2]。结合王幼平、Pimenov等关于羌活属的研究[29,39],发现本研究中尖瓣藁本的果实形态结构与羌活(Notopterygiumincisum)和羽苞羌活(Notopterygiumpinnatiinvolucellatum)均较为相似:羌活具有4条果棱,羽苞羌活为3条果棱;同时它们的果实解剖结构也较相似:胚乳腹面内凹,果棱扩展成翅状,外果皮被角质层,维管束位于果棱基部近中央位置,同时径向伸长(图版Ⅲ,17~18)。此外,本文研究的尖瓣藁本与Pimenov报道的尖瓣藁本除果棱数目不同外,其他特征也较相似,但Pimenov认为尖瓣藁本与羌活为同一种[29]。果实的比较解剖研究表明,尖瓣藁本与羌活属在果实的解剖结构特征上很相似,同时本文果实微形态特征也表明尖瓣藁本与藁本属其他类群具有明显区别,因此我们支持将尖瓣藁本转入羌活属。鉴于尖瓣藁本的果棱在解剖结构上存在变化,其与羌活是否为同一物种,以及它与羽苞羌活的分类地位和系统关系等问题尚有待进一步的研究来证实。本文中我们也发现在果实微形态和解剖特征方面,理塘藁本和尖瓣藁本具有非常相似的结构特征,而与其他类群区别较大,比如果实近两侧扁压、果实表面凸起、背棱明显且与侧棱同形、胚乳横切面为马蹄形、外果皮具有厚的角质层、木薄壁细胞未完全充满整个果棱等。结合前人的研究[29,39],支持将理塘藁本转入羌活属。
3.3 藁本属属下种间亲缘关系探讨
在伞形科的系统分类中,果实作为繁殖器官,由于其本身特性不易受到环境影响,一直受到分类学者的重视[1,28],不仅将其作为分类鉴定的主要依据,果实的形态、背腹压扁程度、果棱的发达程度等还是确定属下系统位置的重要依据。在研究的藁本属20种果实中,短尖藁本与长茎藁本具有相似的果实微形态特征:果实表面粗糙有表皮毛,细胞轮廓不可见,具簇状条纹、表皮毛及规则的鳞片状分泌物。二者的区别为:短尖藁本具有尖囊状和带状表皮毛;后者仅具有尖囊状的表皮毛。在植株形态上,据《Flora of China》描述,二者形态特征非常相似,仅靠果实棱槽内及合生面的油管数目来区别。地理分布上,前者分布于新疆北部,后者为甘肃、青海东南部、四川西部、西藏及云南西北部[1,2]。因此,从宏观形态、地理分布和果实微形态结构来看,二者关系较为密切。此外,本文研究的其他藁本属果实均无表皮毛结构,因此,表皮毛作为重要的鉴定特征,可将二者与藁本属其他种明显区别开。
羽苞藁本和多苞藁本具有较相似的果实微形态及解剖结构特征:均具有长且密的平行条纹和瘤状突起物,果实大小和形状、果棱形状、胚乳横切面形状等相似,维管束位置相同等。二者的不同在于棱槽内的油管数目差别较大。从地理分布来看,二者均分布于云南和四川地区。生境上,前者生于海拔
2 500~4 000 m 的山坡草地,后者为海拔2 800~4 300m 的林下、灌丛、草地及石隙间。宏观形态上,二者具有较多相似特征,如全株被毛、根茎被纤维状枯萎叶鞘、叶片羽状全裂、小总苞片羽状分裂等[1-2]。相关 分 子 系 统 研 究 中[26-27],周静通过nrD-NA ITS及cpDNA 序列构建系统发育树,二者聚为支持率较高的一支(52%BS,93%PP)。由此可见,羽苞藁本和多苞藁本在果实微形态、宏观形态及分子系统方面都具有较高的相似性,显示出较为密切的亲缘关系。

图版Ⅰ 电镜下藁本属果实表面微形态特征PlateⅠ Micromorphological features of pericarp surfaces of Ligusticum L.under SEM

图版Ⅱ 电镜下藁本属及山芎果实表面微形态特征PlateⅡ Micromorphological features of pericarp surfaces of Ligusticum L.and C.reflexumunder SEM
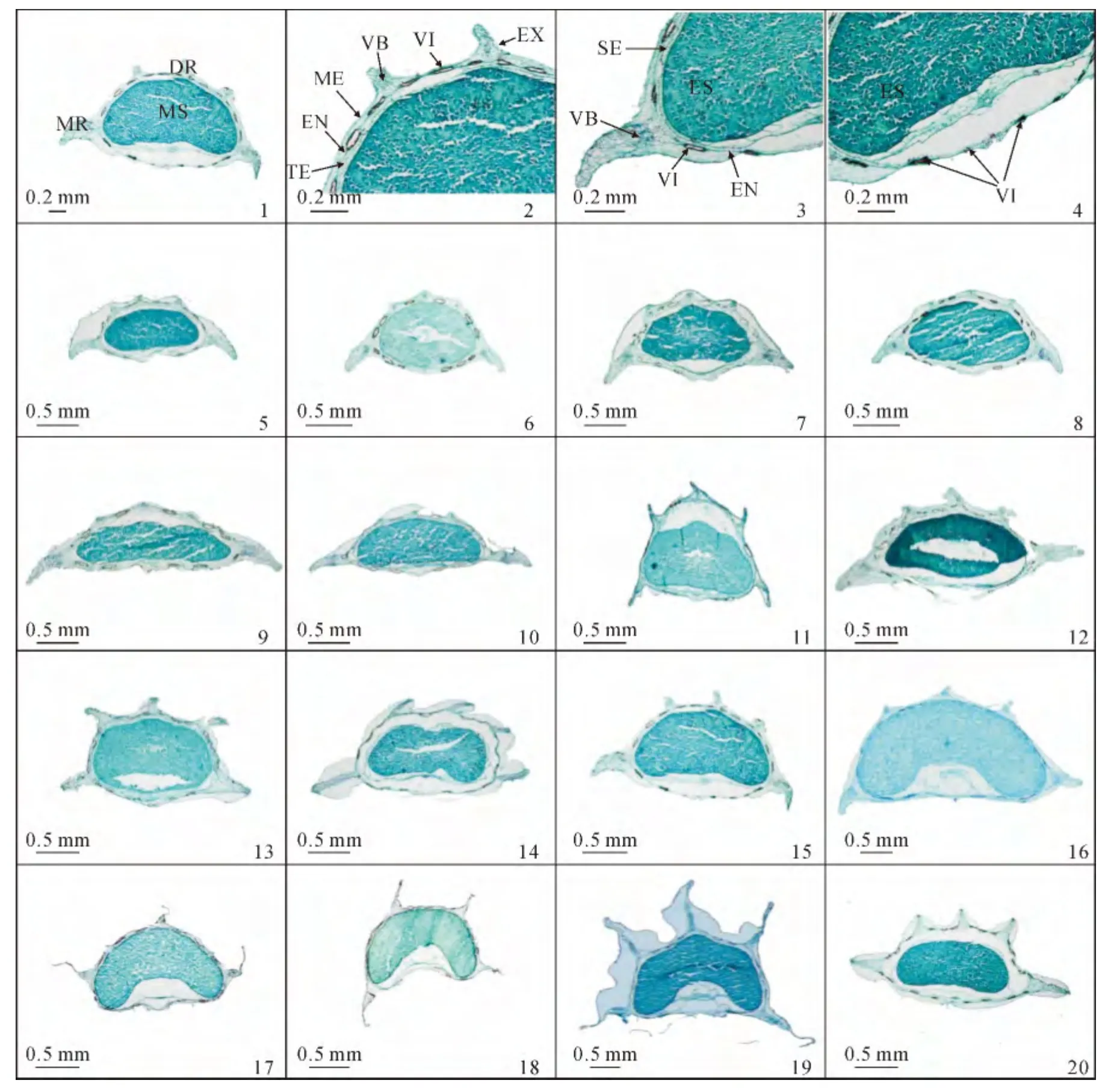
图版Ⅲ 藁本属与山芎属C.reflexum 的果实解剖特征示意图及果实横切面结构PlateⅢ Fruit anatomical features and transection of Ligusticum L.and C.reflexum.
[1] 单人骅,佘孟兰.中国植物志(第55卷第二分册)[M].北京:科学出版社,1985:234-257.
[2] PU F D,WATSON M F.LigusticumL.[A].Flora of China[M].Beijing:Science Press,2005:140-150.
[3] PU F D(溥发鼎).A revision of the genusLigusticum(Umbelliferae)in China[J].ActaPhytotaxonomicaSinica(植物分类学报),1991,29(5):385-393(in Chinese).
[4] VALIEJO-ROMAN C M,SHNEYER V S,SAMIGULLIN T H,etal.An attempt to clarify taxonomic relationships in “Verw and tschaftskre is der GattungLigusticum”(Umbelliferae-Apioideae)by molecular analysis[J].PlantSystematicsandEvolution,2006,257:25-43.
[5] REGEL E,TILING H.Florula Ajanensis[J].NouveauxMémoiresdelaSociétéImpérialedesNaturalistesdeMoscou,1858,1:97.
[6] KOZO-POLJANSKY B M.Sciadophytorum Systematis lineamenta[J].BulletinofMoscowSocietyofNaturalists,1916,2(29):120.
[7] HIROE M,CONSTANCE L.Umbelliferae of Japan(Vol.30)[M].University of California Press,1958:74.
[8] LEUTE G H.Untersuchungen uber den verwandtscnaftskreis der GattungLigusticumL(Umbelliferae).[J].Ann.Nat.Hist.Mus.Wien,1969,78:66-79.
[9] ZHANG B(张 博),SUN J M(孙佳明),CHANG R L(常仁龙),etal.Studies on the chemical constituents of the root and rhizoma ofLigusticumjeholense[J].JournalofChineseMedicinalMaterial(中药材),2009,32(5):710-712(in Chinese).
[10] WANG P L(王萍莉),PU F D(溥发鼎),MA J SH(马建生).Pollen morphology of the genusLigusticumfrom China and its systematic significance[J].ActaPhytotaxonomicaSinica(植物分类学报),1991,29(3):235-245(in Chinese).
[11] POLAND-HEY DAKER F,CERCEAU-LARRIVAL M T.Ultrastructure detectum de pollen d,Ombelliferes[J].GRANA,1978,17:81-89.
[12] 席以珍,孙湘君.中国伞形科花粉形态及其早期演化[C].植物集刊,1983,1:57-84.
[13] DARLINGTON C D,WYLIE A P.Chromosome Atlas of Flowering Plants[M].London:George Allen & Unwin Ltd,1955:251-260.
[14] CONSTANCE L,CHUANG T I,BYE R A.Chromosome numbers in Chihuahuan Umbelliferae[J].BotanicalMuseumLeaflets,HarvardUniversity,1976,24(8):241-247.
[15] ROSTOVTSEVA T S.Chromosome numbers of some species of the family Apiaceae[J].BotZH(Leningr),1979,64(2):227-232.
[16] PAN Z H(潘泽惠),QIN H ZH(秦慧贞),WU ZH J(吴竹君),etal.A report on the chromosome numbers of Chinese Umbelliferae[J].ActaPhytotaxonomicaSinica(植物分类学报),1985,23(2):97-102(in Chinese).
[17] SUN N,HE X J,ZHOU S D.Epidermal morphology ofLigusticum(Apiaceae)from China[J].AnnalesBotaniciFennici,2010,47(4):261-279.
[18] PAN Z H(潘泽惠),WU ZH J(吴竹君),PU F D(溥发鼎).Anatomical studies on petiole ofLigusticumfrom China[J].ActaBotanica Yunnanica(云南植物研究),1992,14(2):143-149(in Chinese).
[19] SUN N,HE X J,ZHOU S D,Morphological cladistic analysis ofLigusticum(Umbelliferae)in China[J].NordicJournalofBotany,2008,26:118-128.
[20] FUSHIMI H,KOMATSU K,ISOBE M,etal.A new approach for the identification of a Chinese traditional medicine,Chuanxiongby 18S ribosomal RNA gene sequences[J].Phytomedicine,1996,(3):387-389.
[21] DOWNIE S R,RAMANATH S,KATZ-DOWNIE D S,etal.Molecular systematics of Apiaceae subfamily Apioideae:Phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacer and plastid rpoC1intron sequences[J].AmericanJournalofBotany,1998,85:563-591.
[22] KATZ-DOWNIE D S,VALIEJO-ROMAN C M,TERENTIEVA E I,etal.Towards a molecular phylogeny of Apiaceae subfamily Apioideae:additional information from nuclear ribosomal DNA ITS sequences[J].PlantSystematicsandEvolution,1999,216(3-4):167-195.
[23] PLUNKETT G M,DOWNIE S R.Major lineages within Apiaceae subfamily Apioideae:A comparison of chloroplast restriction site and DNA sequence data1[J].AmericanJournalofBotany,1999,86(7):1 014-1 026.
[24] DOWNIE S R,WATSON M F,SPALIK K,etal.Molecular systematics of Old World Apioideae(Apiaceae):Relationships among some members of tribe Peucedaneae sensu lato,the placement of several island-endemic species,and resolution within the apioid superclade[J].CanadianJournalofBotany,2000,78:506-528.
[25] DOWNIE S R,KATZ-DOWNIE D S,WATSON M F.A phylogeny of the flowering plant family Apiaceae based on chloroplast DNA rpl16and rpoC1intron sequences:Towards a suprageneric classification of subfamily Apioideae[J].AmericanJournalofBotany,2000,87(2):273-292.
[26] ZH J,GONG X,DOWNIE S R,etal.Towards a more robust molecular phylogeny of Chinese Apiaceae subfamily Apioideae:Additional evidence from nrDNA ITS and cpDNA intron(rpl16and rps16)sequences[J].MolecularPhylogeneticsandEvolution,2009,53(1):56-68.
[27] ZH J,P H,DOWNIE S R,etal.A molecular phylogeny of Chinese Apiaceae subfamily Apioideae inferred from nuclear ribosomal DNA internal transcribed spacer sequences[J].Taxon,2008,57(2):402-416.
[28] LIU M,PLUNKETT G M,LOWRY P P,etal.The taxonomic value of fruit wing types in the order Apiales[J].AmericanJournalof Botany,2006,93(9):1 357-1 368.
[29] PIMENOV M G,KLJUYKOV E V,OSTROUMOVA T A.Towards a clarification in the taxonomy of Sino-Himalayan species ofSelinumL.s.l.(Umbelliferae).The genusOreocomeEdgew[J].Willdenowia,2001,31:101-124.
[30] PIMENOV M G,KLJUYKOV E V,OSTROUMOVA T A.A revision ofConioselinumHoffm.(Umbelliferae)in the Old World[J].Willdenowia,2003,33:353-377.
[31] LAVROVA T V,PIMENOV M G,TIKHOMIROV V N.The anatomy of the petioles as a source of the taxonomic information in the groupLigusticeae(Umbelliferae-Apioideae)[J].BulL.Mosk.Obsc.Isp.Prir.Oid.BioL,1982,87(1):99-111.
[32] MENG D Y(孟德玉).Anatomical studies on fruits and petioles ofLigusticumL.from Sichuan Province[J].ActaBot.Boreal.-Occident.sin.(西北植物学报),2011,31(2):229-235(in Chinese).
[35] DENG X L(邓贤兰),REN H Y(任海燕),PANG Y L(逄云莉),etal.Micromorphological features of pericarp surface ofHeracleum(Apiaceae)in China and its taxonomic significance[J].ActaBot.Boreal.-Occident.sin.(西北植物学报),2009,29(6):1 162-1 169(in Chinese).
[36] MA X G(马祥光),WANG CH B(王长宝),HE X J(何兴金).Micromorphological features of pericarp surface ofBupleurumL.(Apiaceae)in China and its taxonomic significance[J].ActaBot.Boreal.-Occident.sin.(西北植物学报),2010,30(7):1 388-1 396(in Chinese).
[37] GAO P(郜 鹏),LIAO CH Y(廖晨阳),HE X J(何兴金).Micromorphological features of pericarp surface ofPleurospermum(Apiaceae)in China and its taxonomic significance[J].ActaBot.Boreal.-Occident.sin.(西北植物学报),2011,31(9):1 765-1 773(in Chinese).
[38] LI M ZH(李美芝),SONG CH F(宋春凤),LIU Q X(刘启新).Micromorphological features of mericarp surface ofPeucedanumL.(Apiaceae)in China and its taxonomic significance[J].JournalofPlantResourcesandEnvironment(植物资源与环境学报),2012,21(2):19-29(in Chinese).
[39] WANG Y P(王幼平),PU F D(溥发鼎),WANG P L(王萍莉),etal.Studies on systematics of the Chinese endemic genusNotopterygium[J].ActaBotanicaYunnanica(云南植物研究),1996,18(4):424~430(in Chinese).
[40] WANG CH B(王长宝),MA X G(马祥光),HE X J(何兴金).Fruit features of someBupleurumspecies(Apiaceae)and their systematical implication[J].PlantScienceJournal(植物科学学报),2011,29(4):399-408(in Chinese).
[41] WANG H P(王会朋),MA X G(马祥光),GAO P(郜 鹏),etal.Fruit anatomical features of sixteen species and one variety ofPleurospermum(Apiaceae)in China and their taxonomic significance[J].ActaBot.Boreal-Occident.sin.(西北植物学报),2014,34(4):711-719(in Chinese).
[42] LIU CH J(刘长江),LIN Q(林 祁),HE J X(贺建秀).Methods and terminology of study on seed morphology from China[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2004,24(1):178-188(in Chinese).
猜你喜欢
西北植物学报(2022年4期)2022-05-24
科学大众·教师版(2022年6期)2022-05-23
北京航空航天大学学报(2021年9期)2021-11-02
新农民(2021年6期)2021-09-15
生物学通报(2021年4期)2021-03-16
中国中医药现代远程教育(2019年17期)2019-01-30
中成药(2017年10期)2017-11-16
种子科技(2017年4期)2017-01-13
乡村科技(2016年33期)2016-02-19
中国医药指南(2011年30期)2011-02-11
