
外源激素对热研7–33–97 低割龄橡胶树单株产量 及其构成因素的影响
2015-07-15 05:42吴春太黎瑜聂智毅康桂娟代龙军曾日中
湖南农业大学学报(自然科学版) 2015年5期
吴春太,黎瑜,聂智毅,康桂娟,代龙军,曾日中*
(1.中国热带农业科学院橡胶研究所,海南 儋州 571737;2.农业部橡胶树生物学与遗传资源利用重点实验室,海南 儋州 571737;3.海南省热带作物栽培生理学重点实验室,海南 儋州 571737)
巴西橡胶树(Hevea brasiliensis)的次生代谢是 在长期演化过程中形成的,其与树体的生长发育及对环境的适应密切相关。胶乳为橡胶树次生代谢产物中数量最多的一种多聚萜类化合物[1]。胶乳除抵御生物或非生物胁迫,保护橡胶树外[2],还具有重要的工业使用价值[3–4]。由于许多逆境因子都会诱导巴西橡胶产生抗逆反应,反应的主要表现是伤口处流胶[5]。流出的天然胶乳是橡胶树为了保护其自身而产生的重要防御物质,主体为聚顺式1,4–异戊二烯,因此,抗逆反应发生后树体可能会启动次生代谢而合成大量胶乳。据报道,切割树皮和施用乙烯是天然橡胶生产的两种主要刺激因子,其中乙烯对产量的影响主要通过促进排胶来实现[6–7]。为对橡胶树目的次生代谢产物的生物合成途径进行调控,橡胶生产中通常采用单方、复方乙烯利作为诱导子,而且同一品种在不同割龄段施用不同浓度的乙烯利,同一割龄段的不同类型品种所适应的浓度也存有差异。现今施用乙烯利作为橡胶增产的重要手段已普遍应用于橡胶树主要栽培品种,并取得了良好的生产效果。黄瑾等[8]研究表明,不同排胶特性的高割龄橡胶树在受乙烯利刺激后胶乳产量均高于刺激前的产量,而其增产幅度存在着品系差异,表现为PR107、GT1、RRIM600的产量依次递减。吴明等[9]对低割龄(2~3年)热研7–33–97早期应用刺激剂的产量效应进行了研究,结果表明,采用乙烯利刺激割胶的热研7–33–97成龄幼树试验第1年表现出高产效应,但第2年却表现出低产效应。罗世巧等[10]以2~4年割龄热研7–33–97为试材,研究了开割幼树对乙烯利刺激低频割制的适应性,试验发现该品种的成龄幼树对乙烯利刺激低频割胶表现出胶乳再生能力强、排胶速度快、生长受抑制的程度轻等生理特性。2007年,Jetro和Simon发现乙烯刺激对胶乳产量和胶乳生理的影响作用随季节而变化[11]。2011年,Njukeng等[12]提出刺激的频率和浓度应作为品种、树龄和割制的一个方面被采用。综上可见,乙烯利是影响橡胶产量的重要因素。同时,施用乙烯利也被认为是提高天然橡胶生产效率和缓解胶工缺乏的最有效途径[13]。此外,割胶时单株胶乳体积的多少一直是衡量单位面积橡胶产量重要的标志之一[14]。
近年来的研究发现,植物体内的茉莉酸及其衍生物不仅是植物受外界刺激后反应最快的信号分子,且还是植物次生代谢物积累的前提,如能诱导或抑制植物次生代谢产物积累等[15–17],因其重要的基础研究价值,茉莉酸类物质也越来越受到橡胶学家们的高度关注[18],而茉莉酸类物质对巴西橡胶树胶乳积累的影响和机理方面的研究尚少见报道。尽管乙烯利对刺激后的橡胶树的产量形成有较大影响,基于植物次生代谢产物合成的代谢途径多样性,人们想尝试通过不同的诱导子刺激代谢途径来增加橡胶树目标次生产物的合成量,但目前还很少有关于橡胶生产上应用茉莉酸的报道。以往的研究侧重乙烯利对产量的影响,而不同激素和刀次对产量及其构成因素的影响则研究较少,而且在近几年胶价持续低迷的国内各植胶区,由于胶工缺乏和产量与效益矛盾突出等原因,胶树闲置的现象十分普遍,高产、高效和安全采胶已成为现代橡胶产业的当务之急。为探明不同激素对橡胶生产的影响效果,结合中国橡胶生产特点,本试验中以热研7–33–97为对象,对不同激素刺激下橡胶树单株不同刀次产量及其构成性状的表现进行了研究,试图得到适宜于低割龄橡胶树高产所需的激素,旨在为大面积高产、高效生产应用提供理论指导。
1 材料与方法
1.1 供试材料的概况
试验材料为2005年种植的高产、抗风橡胶品种热研7–33–97成龄幼树。种植规格7 m×3 m,田间管理同常规胶园。2012年开割,试验当年割龄为3年。
1.2 试验设计
试验于2014年在中国热带农业科学院(儋州)试验场十二队的橡胶林段进行。激素涂施前、后采胶试验全部采用半树围,3 d一刀(S/2 、d/3)割胶制度。激素施用后,试验分别采用3种割制对样树进行割胶:1) 采用单阳线非激素刺激的割制割胶;2) 采用单阳线单方乙烯利(0.5%ET) 刺激的割制,涂药时间选择在割胶前24 h,统一使用液剂,在割线下方涂施,每株来回刷2次;3) 采用单阳线复方茉莉酸(0.3% JA+羊毛脂) 刺激的割制,羊毛脂实为茉莉酸的载体,统一使用糊剂,涂施前刮去橡胶树割线下方的外周木栓层,每株涂2g,割胶前24 h完成涂药。各个处理的割胶株数均为6株。每种处理设置3次重复。
1.3 调查内容与方法
于2014年9月4日、9月7日和9月11日按单株测产的方式对样树进行3个割次的试前产量鉴定;分别于10月10日、10月13日、10月16日、10月19日和10月22日对样株进行试后第1、2、3、4、5次采胶测产。所有测次胶乳产量采用量筒测定。干胶含量测定采用标准烘箱干燥法。
1.4 数据处理
以不使用激素的空白为对照,计算出激素刺激割胶所得产量及其构成因子数据的净增产(长)量和净增产(长)率[13,19]。净增产(长)量=试后处理组产(含)量–试前处理组产(含)量×(试后对照组产(含)量/试前对照组产(含)量);净增产(长)率=(试后处理组产(含)量×试前对照组产(含)量/(试后对照组产(含)量×试前处理组产(含)量))×1–1。所获数据经Excel 2003整理后,采用SAS8.0对数据进行方差分析,并采用新复极差法(Duncan’s)进行多重比较。
2 结果与分析
2.1 常规割制下橡胶树单株产量及其构成性状的表现
在施用激素前,对不同组热研7–33–97开割幼树进行了3个割次的胶乳测产与数据分析(表1)。结果表明,同一组样树试前测产期每割次株产胶乳量存在一定变化,其中,变化最大的是ETH试验区,其每割次胶乳最高产量为其最低值(148.00mL)的1.34倍,变化最小的是JA试验区,其每割次胶乳最高与最低产量相差34.32mL,两者差异显著(P<0.05);不同组之间第2、3割次的产量差异明显,同一割次株产胶乳产量均以ETH试验区最高,其次为JA试验区,而且ETH试验区的3个割次平均株产胶乳量高于对照试验区和JA试验区,ETH试验区的值分别为JA试验区的1.07倍、CK试验区的1.41倍,其中仅与CK试验区的差异达显著水平。
从表1 还可以看出,同一组样树不同割次干胶含量变化较小,其中,变化较大的是ETH 试验区,其每割次干胶含量最高水平为其最低值(30.07%)的1.08 倍,变化最小的是CK,其每割次干胶含量最高与最低水平仅相差2.13%;不同组之间同一割次干胶含量和3个割次平均干胶含量差异均不显著。
同一组样树不同割次株产干胶量存在一定变化,其中,变化最大的是CK 试验区,其每割次株产干胶最高产量为其最低值(34.16g)的1.37 倍,变化最小的是JA 试验区,其每割次株产干胶最高与最低产量相差11.14g,两者差异显著(P<0.05);不同组之间同一割次株产干胶量和3个割次平均株产干胶量差异均不显著。
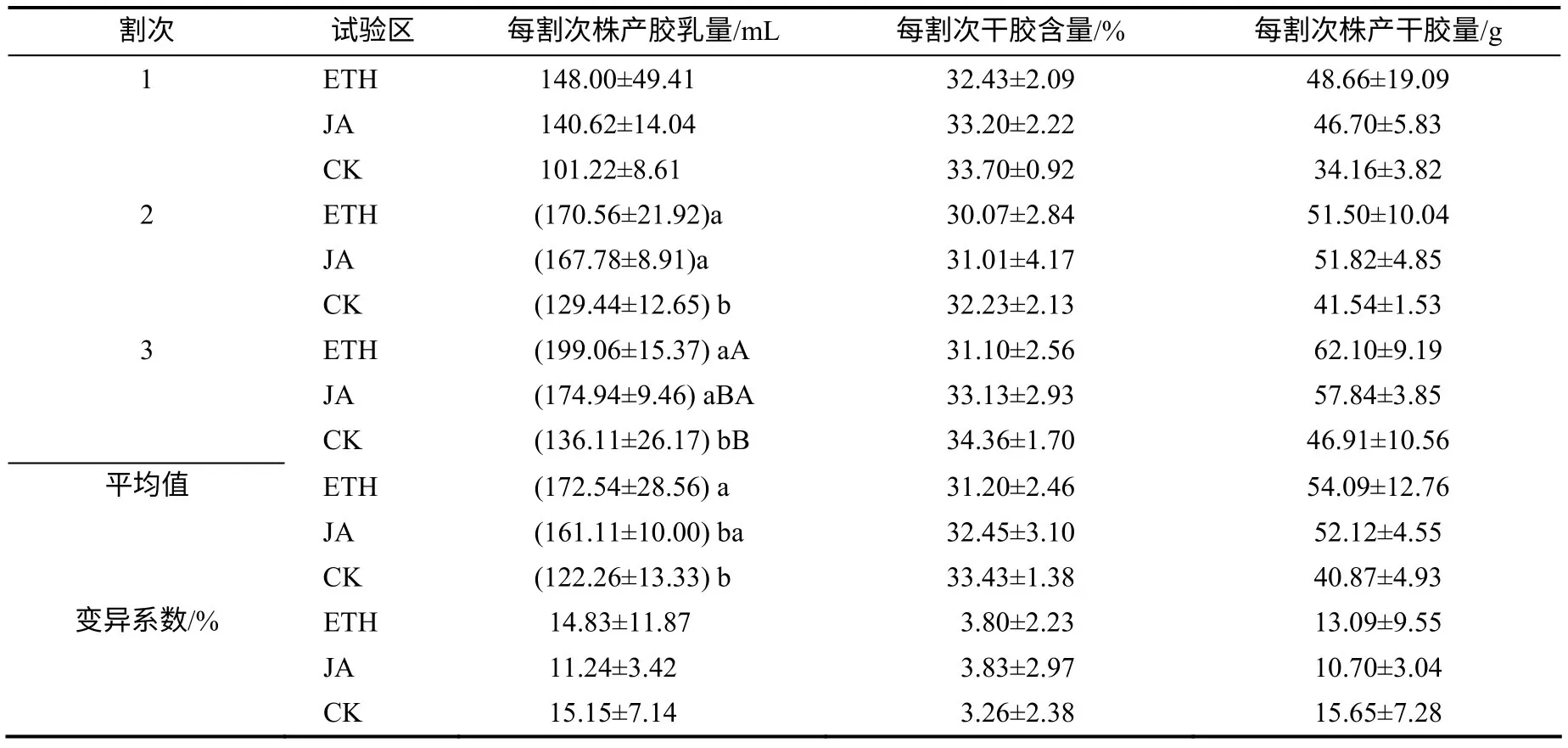
表1 激素处理前橡胶树的产量及产量性状 Table 1 Yield performance and yield component traits of Hevea brasiliensis before treatment with hormone
2.2 不同外源激素刺激下橡胶树单株产量及其构成因素的分析
对施用不同激素的热研7–33–97 开割幼树5个割次的胶乳产量观测数据(表2)进行分析,结果表明,同一处理单个周期每割次株产胶乳量存在一定变化,其中,变化最大的是ETH,其每割次胶乳最高产量为其最低值(147.05mL)的1.83 倍,两者差异显著(P<0.05),变化最小的是CK,其每割次胶乳最高与最低产量相差 65.89mL,两者差异显著(P<0.05);不同处理间第2、3、5 割次的产量差异明显,同一割次株产胶乳产量均以ETH 最高,其次为CK,而且,ETH 的5个割次平均株产胶乳量高于对照和JA,与JA、CK 的差异分别达极显著和显著水平, ETH 的胶乳产量分别为JA 的1.61 倍、CK 的1.33 倍。
从表2 可以看出,同一处理不同割次干胶含量变化较小,其中,变化最大的是ETH,其每割次干胶含量最高值为其最低值(29.52%)的1.18 倍,变化最小的是JA,其每割次干胶含量最高与最低水平仅相差1.51%;不同处理间第4、5 割次的干胶含量差异明显,其中,第4、5 割次平均干胶含量分别以CK、JA 最高,而且,对照、JA 的5个割次平均干胶含量高于ETH,与ETH 的差异分别达极显著和显著水平,其中,CK、JA 的干胶含量分别为ETH的1.11 倍、1.09 倍。
同一处理不同割次株产干胶量存在一定变化,其中,变化最大的是JA,其每割次株产干胶最高产量为其最低值(33.07g)的1.83 倍,两者差异极显著(P<0.01),变化最小的是CK,其每割次株产干胶最高与最低产量相差21.70g,两者差异显著(P<0.05);不同处理间第1、2、3 割次的产量差异明显,同一割次株产干胶量均以ETH 最高,其次为CK,而且,ETH 的5个不同割次平均株产干胶量均高于对照和JA,ETH 的值分别为JA 的1.48 倍,CK 的1.21 倍,其中,仅与JA 的差异达极显著水平。
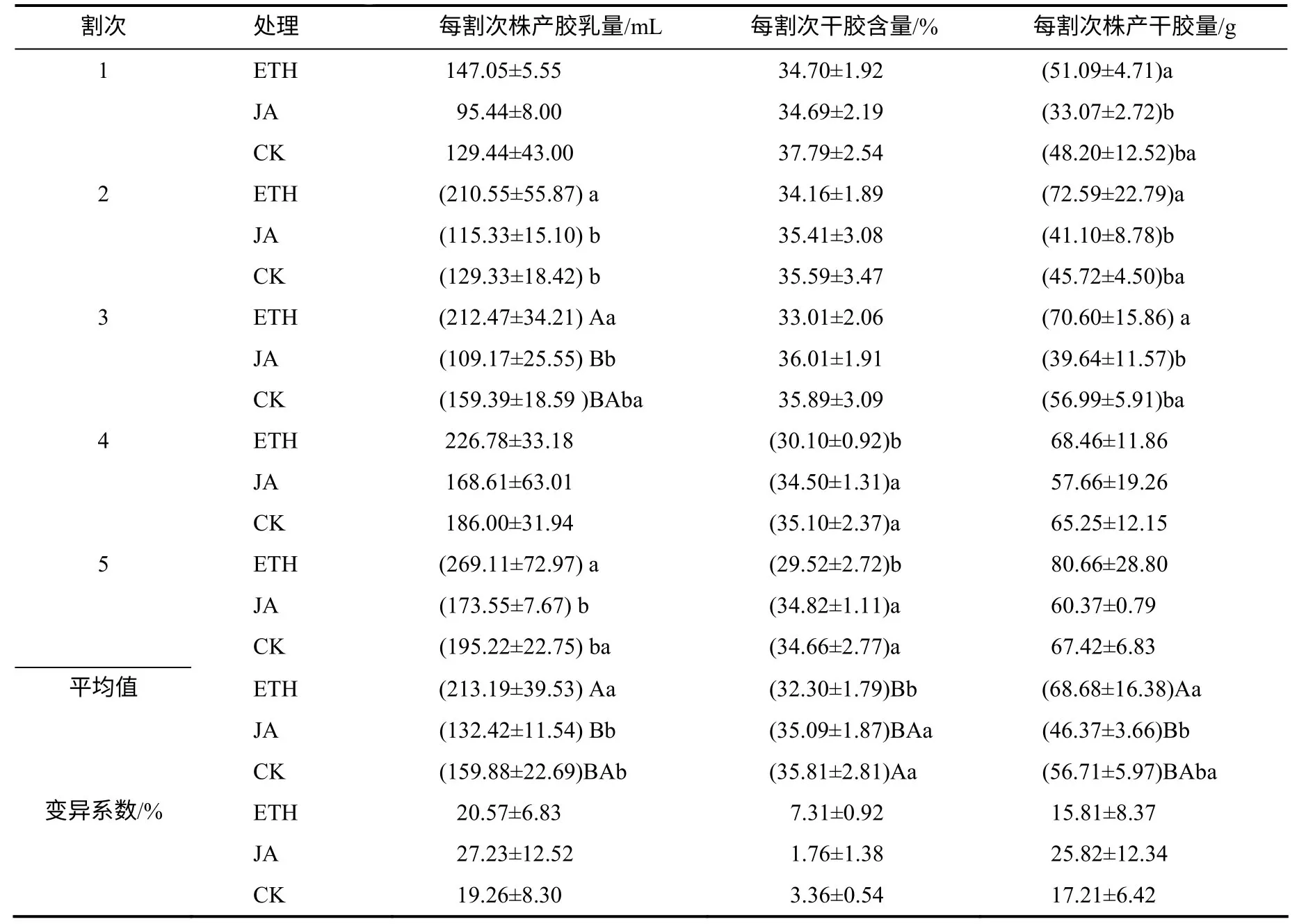
表2 不同激素处理后的干胶产量及产量性状 Table 2 Change of dry rubber yield and yield component traits in CATAS 7–33–97 after treatment with different hormones
2.3 不同割制下的橡胶树单株产量及其构成因素的比较
3种处理的低割龄热研7–33–97试验前后产量与产量构成因子的差异显著性分析结果表明,同一处理组不同割制的单株胶乳、干胶产量和干胶含量存在一定变化。橡胶树采用乙烯利刺激后,除第1刀次外,第2~5刀次的每割次株产胶乳量均高于其试前采用常规割制的量;其试后5个割次的平均干胶含量高于试前3个割次的均值,但差异不显著(P>0.05);除第1刀次外,第2~5刀次的每割次株产干胶量均高于试前采用常规割制的量。采用JA刺激后的橡胶树,该测产周期5个割次的平均株产胶乳量低于试前3个割次的均值,但差异不显著(P>0.05);其试后5个割次的平均干胶含量高于试前3个割次的平均值,差异达极显著水平(P<0.01);其5个割次的平均株产干胶量低于试前3个割次的均值,但差异不显著(P>0.05)。继续采用常规割制采胶的橡胶树,除第1~2刀次外,第3~5刀次的每割次株产胶乳量均高于其试前的量;其试后5个割次的干胶含量均高于试前3个割次的量,差异达显著水平(P<0.05);除第2刀次外,第1、3、4、5刀次的每割次株产干胶量均高于试前采用常规割制的量。
比较表1、表2中的变异系数可以看出,试前、试后3个处理的性状变异系数大小排序发生了变化:试前3个割次的胶乳量变异系数由大到小的处理分别为CK、ETH、JA,干胶产量变异系数由大到小的处理分别为CK、ETH、JA,干胶含量变异系数由大到小的处理分别为JA、ETH、CK;试后5个割次的胶乳量变异系数由大到小的处理分别为JA、ETH、CK,干胶产量变异系数由大到小的处理分别为JA、CK、ETH,干胶含量变异系数由大到小的处理分别为ETH、CK、JA。
2.4 不同割制干胶产量及其构成因素的净增长量与净增长率比较
由表3、表4 可知,ETH 处理不同割次净干胶含量增长量和增长率存在一定变化。第2 割次净干胶含量增长量依次比第1、3、4、5 割次超出1.61、1.45、3.67、3.84mg/mL,且与第4、5 割次的差异达显著水平;第2 割次净干胶含量增长率分别比第1、3、4、5 割次超出4.65%、4.45%、11.04%、11.93%,且与第4、5 割次的差异均达显著水平;其他割次间的净干胶含量增长量和增长率差异均无统计学意义。
不同激素刺激割制干胶产量及其构成因素的净增长量与净增长率存在一定变化。ETH 的净胶乳量、干胶增产量和净胶乳、干胶增产率分别比JA的超出69.41mL、21.67 kg、33.31%、28.43%,差异达极显著水平;ETH 的5个割次平均净干胶含量增长量、增长率与JA 的差异均无统计学意义。
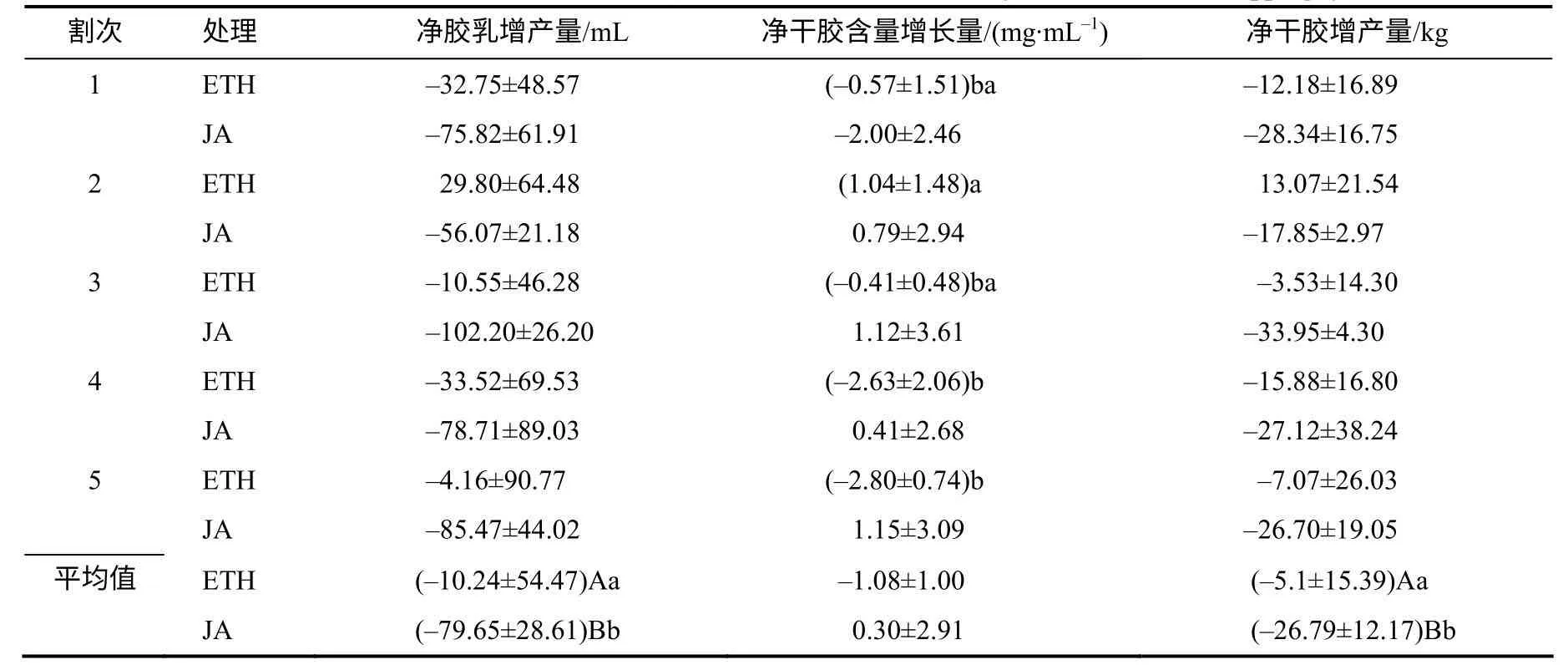
表3 不同割制干胶含量与橡胶产量的净增长量 Table 3 Comparison of the net increased dry rubber content and rubber yield from different tapping systems
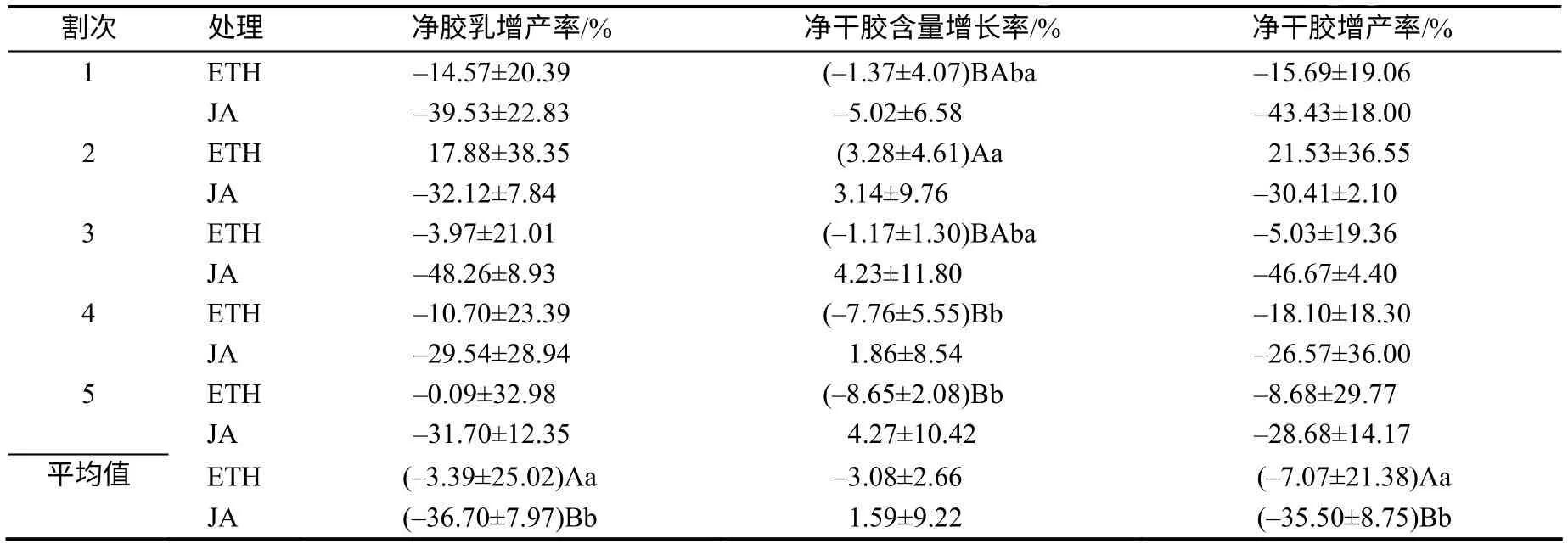
表4 不同割制干胶含量与橡胶产量的净增长率 Table 4 Comparison on the net inerease rate of dry rubber content and rubber yield from different tapping systems
3 小结与讨论
本研究结果表明,应用于热研7–33–97 开割幼树的外源激素增产效果不一,0.5%乙烯利对未使用乙烯利刺激割胶的3 割龄胶树产量有显著影响,而0.3%茉莉酸的影响效果则不明显。这说明乙烯利诱导产量的效应明显强于茉莉酸,其原因可能是所用茉莉酸浓度过高,高浓度的茉莉酸类物质对植物次生代谢产物合成没能起到促进作用,反而起着抑制作用[20]。本试验中,以10月13日割胶、ETH 处理时产量最高,达72.59g/株。ETH 处理的平均实收产量分别比对照和JA 处理增产21.53%、51.94%。其第2 刀次(10月13日)的平均实收产量比第1、3、4、5 刀次分别增产37.22%、26.56%、39.63%和30.21%,说明热研7–33–97 涂施乙烯利的最适增产期为第2 刀次左右。
本试验发现热研7–33–97 成龄幼树产量构成因素对乙烯利和茉莉酸的响应也有差异。乙烯利对胶乳产量影响较大,以第2 刀次净胶乳增产率最高,达17.88%,其他刀次的净胶乳增产率均呈负增长,但不同刀次间净胶乳增产率差异不明显。乙烯利对干胶含量影响也较大,不同刀次间净干胶含量增长率呈显著差异。以第2 刀次净干胶含量增长率最高,达3.28%。随其后刀次增加,净干胶含量增长率下降。通过比较第2 刀次的净胶乳增产率与净干胶含量增长率大小,可以推测乙烯利实现增产效应的主要方式可能是促进胶乳排胶, 而非促使胶乳再生。茉莉酸对干胶含量影响较大,随着割次增多(第1割次除外),茉莉酸处理的干胶含量有增加的趋势。这可能是由于橡胶树本身的调节功能引起,当每割次株产胶乳量减少时,植株分配到橡胶烃上的营养成份增加,干胶含量相应增加。但由于每割次株产胶乳量下降,干胶产量也随之下降,因此,采用上述2 种割制采胶时,胶乳产量是影响低割龄热研7–33–97 干胶产量的主要因素,干胶含量对干胶产量影响相对较小。
热研7–33–97 开割幼树应用不同浓度乙烯利采胶增产效果不一。早在1998年吴明等[9]采用半树围,4 d 一刀割制,并以0.5%、1.0% ET 2 种浓度对2年割龄热研7–33–97 进行刺激割胶试验,其全年的干胶产量分别比对照增产41.1%、53.8%,但干胶含量依次比对照下降3.9%、4.8%。罗世巧等[10]根据2~4年割龄热研7–33–97 对乙烯利刺激低频割制的适应性试验结果表明,低割龄热研7–33–97 采胶不宜使用高浓度乙烯利刺激,因为在相同割制下, 施用1.0%乙烯利的干胶产量比施用0.5%乙烯利的减产6.7%,其干胶含量相应下降。即使采用双短线低强度(S/4+S/4↑ d/6+1.5%~2.0% ET)割制割胶,其割胶强度约比(S/4+S/4↑ d/4+0.5%~1.0% ET)下降17%,但高浓度乙烯利刺激并没有比低浓度处理获得更高的产量,因此,本研究选用的乙烯利浓度(0.5%)是科学的,并取得了良好的效果。第2 刀次时,乙烯利刺激采割的干胶产量大幅度增加,同一刀次的干胶含量也上升,说明该品种的乳管系统具有较大的胶乳再生潜力。但其他刀次的胶乳量、干胶产量和干胶含量均呈负增长,且整个药效周期内所有刀次的平均胶乳量、干胶产量和干胶含量均下降,说明热研7–33–97 在初产期对乙烯利刺激响应速度慢,高产期短,且产量不稳定,这与罗世巧等[10]于1999年采用多种割制对3年割龄热研7–33–97 进行乙烯利刺激割胶均难以获得良好的产量效应吻合,其原因可能与品种的特性和过早使用刺激剂有关。
茉莉酸刺激条件下,第2~5刀次胶乳的干胶含量均上升,但试后所有刀次的胶乳量、干胶产量和第1刀次的干胶含量均下降,这表明外施茉莉酸可以提高胶乳的干胶含量,与前人的外施茉莉酸甲酯可以提高胶乳的干胶含量结果相似[21–22]。然而,茉莉酸带来的干胶含量提高是以产量降低为代价,这与前人的茉莉酸类物质能够促进现有乳管内橡胶的生物合成结论不一致,因此,干胶含量升高是否是因茉莉酸类物质促进有效乳管中的天然橡胶生物合成,或该类物质抑制了有效乳管的排胶有待进一步研究。
本试验仅仅就不同种类激素对热研7–33–97 成龄幼树不同刀次的产量及其构成因素的影响作了初步研究,而对产量的形成规律未作探索。对激素浓度和最适割龄期的选择,以及乙烯利和茉莉酸生产上的适应性还需进行多年、多点试验。
[1] Duan C,Argout X,Gébelin V,et al. Identification of the Hevea brasiliensis AP2/ERF superfamily by RNA sequencing[J].BMC Genomics,2013,14:30.
[2] Konno K.Plant latex and other exudates as plant defense systems:Roles of various defense chemicals and proteins contained therein[J].Phytochemistry,2011,72:1510–1530.
[3] Ko J H,Chow K S,Han K H.Transcriptome analysis reveals novel features of the molecular events occurring in the laticifers of Hevea brasiliensis (para rubber tree)[J].Plant Molecular Biology,2003,53:479–492.
[4] Chow K S,Mat-Isa M N,Bahari A,et al.Metabolic routes affecting rubber biosynthesis in Hevea brasiliensis latex[J].Journal of Experimental Botany,2012,63(5):1863–1871.
[5] 吴春太,黎瑜,代龙军,等.橡胶树甾醇14α–去甲基化酶基因(HbCYP51)的克隆及机械损伤和外源激素对其表达的影响[J].农业生物技术学报,2014,22(12):1525–1535.
[6] Tungngoen K,Kongsawadworakul P,Viboonjun U,et al. Involvement of HbPIP2,1 and HbTIP1,1 aquaporins in ethylene stimulation of latex yield through regulation of water exchanges between inner liber and latex cells in Hevea brasiliensis[J].Plant Physiol,2009,151(2):843–856.
[7] Zhu J,Zhang Z.Ethylene stimulation of latex production in Hevea brasiliensis[J].Plant Signaling & Behavior,2009,4(11):1072–1074.
[8] 黄瑾,校现周.乙烯利和乙烯刺激对橡胶树胶乳中几丁质酶活性和胶乳产量的影响[J].热带作物学报,2003,24(4):1–5.
[9] 吴明,罗世巧,校现周.橡胶树新品种热研7– 33– 97早刺激产量和生理效应研究初报[J].云南热作科技,2000,23(3):1–4.
[10] 罗世巧,校现周,魏小弟,等.热研7–33–97幼龄开割树对低频刺激割制的适应性研究初报[J].热带作物学报,2005,26(4):28–33.
[11] Jetro N N,Simon G M.Effects of 2–chloroethyl- phosphonic acid formulations as yield stimulants on Hevea brasiliensis[J].African Journal of Biotechnology,2007,6:523–528.
[12] Njukeng J N,Muenyi P M ,Ngane B K,et al.Ethephon stimulation and yield response of some Hevea clones in the humid forests of south west Cameroon[J]. [EB/OL]. http://www.hindawi.com/journals/ija/2011/257340/refl.pd f.DOI:10.115512011/257340.
[13] 周敏,黄强,刘学敏,等.橡胶树树干注射乙烯利增产效应研究[J].热带农业科学,2012,32(10):1–5.
[14] 吴春太,马征宇,刘汉文,等.橡胶RRIM600的产量与产量构成因素的通径分析[J].湖南农业大学学报:自然科学版,2014,40(5):476–480.
[15] 朱家红,彭世清.茉莉酸及其信号传导研究进展[J].西北植物学报,2006,26(10):2166–2172.
[16] Devoto A,Ellis C,Magusin A,et al.Expression profiling reveals COI1 to be a key regulator ofgenes involved in wound- and methyl jasmonate-induced secondary metabolism,defence,and hormone interactions[J].Plant Mol Biol,2005,58:497–513.
[17] Browse J.Jasmonate passes muster:A receptor and targets for the defense hormone[J].Annu Rev Plant Biol,2009,60:183–205.
[18] Pirrello J,Leclercq J,Dessailly F,et al. Transcriptional and post-transcriptional regulation of the jasmonate signalling pathway in response to abiotic and harvesting stress in Hevea brasiliensis[J].BMC Plant Biology,2014,14:341.
[19] 杨文凤,牛静明,罗世巧,等.橡胶树气刺割胶中不同割线长度生理效应研究[J].热带作物学报,2012,33(11):1971–1975.
[20] 施江,马成英,吕海鹏,等.外源茉莉酸甲酯改善农产品品质及其机理研究进展[J].食品工业科技,2013 (13):364–368.
[21] 邹智,杨礼富,王真辉,等.橡胶树中橡胶的生物合成与调控[J].植物生理学通讯,2009,45(12):1231–1238.
[22] 于俊红,黄绵佳,田维敏.巴西橡胶树橡胶生物合成调控的研究进展[J].2007,13(12):38–40.
猜你喜欢
热带生物学报(2022年1期)2022-03-09
农业科技与信息(2021年9期)2021-12-07
热带农业科技(2021年3期)2021-07-10
北京信息科技大学学报(自然科学版)(2020年5期)2020-11-19
热带农业科技(2019年4期)2020-01-15
世界热带农业信息(2019年2期)2019-05-17
橡胶工业(2018年11期)2018-07-23
贵州科学(2016年5期)2016-11-29
世界热带农业信息(2016年6期)2016-07-02
橡塑资源利用(2015年1期)2015-12-24
