
亚东鲑人工繁殖与野生群体的形态和微卫星多态性分析
2015-10-20 12:23李福贵但唐兴林绍南邹曙明
江苏农业科学 2015年5期
关键词:形态特征
李福贵+但唐兴+林绍南+邹曙明
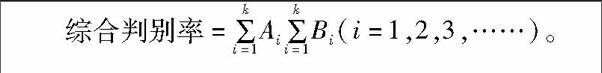

摘要:开展亚东鲑野生群体与人工繁殖后代的形态特征和微卫星遗传多态性分析。首先采用方差、判别、主成分分析方法研究了2个群体,包括7个可数、11个可量与20个框架数据,结果表明亚东鲑人工繁殖后代与野生群体在鳍式、鳞式、可量和框架结构性状等方面均无显著差异。进一步采用9对微卫星引物评估了亚东鲑人工繁殖与野生群体问的遗传变异,在10个座位中,共检测到55个等位基因,平均有效等位基因分别为4.8、4.0个,平均每个座位的等位基因数为5.5个;平均座位观测杂合度分别为0.6267、0.6417,平均基因多样性均为0.5719,群体遗传分化较小,2个群体的微卫星DNA多态性均呈现低水平。结果表明,亚东鲑人工繁殖与野生群体在遗传上差异不大,人工繁殖群体可用于开展进一步的保种繁育和保护性放流工作。
关键词:亚东鲑;野生群体;人工繁殖群体;形态特征;微卫星多态性
中图分类号:S932.4 文献标志码:A 文章编号:1002-1302(2015)05-0226-04
亚东鲑(Salmo trutta fraio Linne)属鲑形目鲑科鲑属,别称河鲑(俗称花点鲑),是冷水性珍稀鱼类,具有较高的营养价值和经济价值。关于亚东鲑的研究较少,张春霖等先后对亚东鲑的来源和资源情况进行过分析;蒲德永等对亚东鲑消化系统的形态学和组织学进行过观察研究;豪富华等对亚东鲑的性腺发育生长年龄与繁殖进行过研究。在群体遗传学方面,豪富华利用线粒体和微卫星标记对亚东鲑种群和丹麦引进棕鳟种群进行过群体遗传多样性研究,孟玮等基于线粒体C01基因序列对亚东鲑进行了DNA条形码研究。然而目前关于亚东鲑人工繁殖后代和野生群体的群体遗传学比较研究尚未开展。
亚东鲑自19世纪由英国人从欧洲引种后,仅栖息于我国西藏自治区日喀则地区亚东县海拔约3000m的亚东河,1992年已被列入西藏自治区二级保护水生动物,尤其是近些年来,野生亚东鲑资源量有加快衰退的趋势。开展亚东鲑的人工繁殖和放流有利于资源的恢复,须要选择具有与原种相似遗传多样性的亲本,鉴于当前野生亚东鲑资源量萎缩,从人工繁殖后代中培育亲鱼是解决亲本来源的有效途径。有关于野生和人工繁殖亚东鲑群体间形态差异及微卫星多态性的研究未见报道,本研究通过分析人工繁殖与野生西藏亚东鲑2个群体间的形态差异和微卫星DNA多态性,旨在了解2个群体的遗传分化特征,为亚东鲑保种繁育和保护性放流工作提供理论资料。
1.材料与方法
1.1试验材料
于2012年从西藏自治区日喀则地区亚东县亚东河采集野生亚东鲑,先后捕获30尾,体长、体质量分别为(20.42±2.70)cm、(155.01±72.72)g,未进行年龄鉴别;亚东鲑人工繁殖岳代1~2龄鱼取自西藏亚东鲑渔场,样本数量也为30尾,体长、体质量均值分别为(16.34±4.26)cm、(92.79±87.98)g,其亲本系由亚东河采集的野生个体培育而成。剪鱼鳍保存于无水乙醇中,用于抽提基因组DNA,于-20℃冰箱保存。
1.2形态特征分析
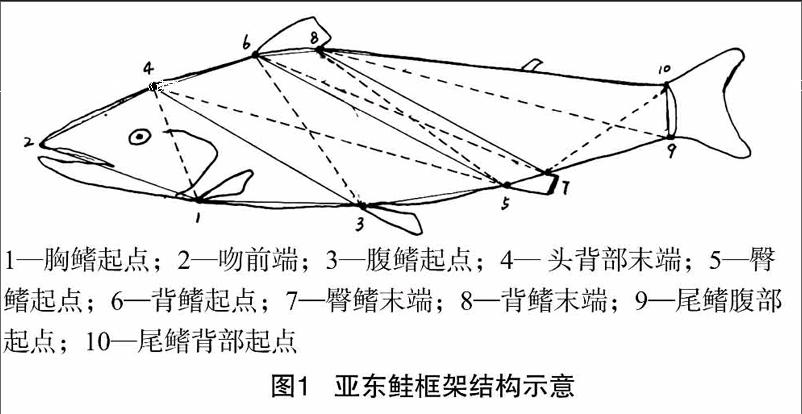
1.2.1数据采集测量的数据分为2类:一类是传统形态学数据,包括可数、可量性状共18项,其中可数性状包括背鳍、胸鳍、腹鳍、臀鳍鳍条、侧线鳞、侧线上鳞、侧线下鳞;可量性状有头长、吻长、眼径、眼间距、尾柄长、尾柄高、体长、全长、体高、背棘长、体质量。另一类为框架数据,共20项,20个框架数据是10个定位点之间的距离,例如D1-2表示定位点1与2之间的距离。框架测量定位点的选择参照图1。
1.2.2数据处理和多元统计分析为消除鱼体规格大小对形态分析的影响,每尾鱼的所有实测可量性状(除体质量外)均用体长进行校正。再分别求出各组样本每个参数的平均比值,用于多元统计分析。分析数据包括7个可数性状、111个可量性状及20个框架性状,分别用方差分析进行差异性检验,如果方差分析检验为差异显著(P<0.05)或极显著(P<0.01),则用Duncan's多重比较进行分析。
采用逐步判别法对所有的形态数据进行筛选,去除判别效果不显著的参数,计算判别准确率和互相证实准确率,建立判别方程。判别准确率P1=判别正确的尾数/实际尾数×100%。互相证实准确率P2:在共有Ⅳ个样品中,每次留下一个样品作为新样品,由N-1个样品建立判别函数,然后将留下的这个样品代入判别函数,判别其归属,其计算方法同P1。式中:Ai为第i个群体判别正确的尾数,尾;Bi为第i群体的实际尾数,尾;K为群体数。
主成分分析采用降维计算,是将多个指标转化为少数几个综合指标的统计方法。
上述所有数据均用SPSS13.0软件进行统计分析。
1.3亚东鲑微卫星DNA多态性分析
1.3.1简单重复序列(simple sequence repeat,SSR)引物序列
参考已报道的相关文献,共设计9对SSR引物,由生工生物工程(上海)股份有限公司合成,引物序列与PCR反应条件见表1。
1.3.2全基因组DNA提取试验前将鱼鳍从乙醇中取出,用蒸馏水洗净、吸水纸吸干、剪碎,按照海洋动物组织DNA抽提法[由天根生化科技(北京)有限公司提供的试剂盒]抽提全基因组DNA,用1.2%的琼脂糖凝胶电泳检测抽提的DNA的质量、浓度。
1.3.3SSRPCR反应体系与程序、PCR产物检测PCR反应体系为25μL:2.5μL10×buffer,1μLdNTP,0.5μLTaq-E,1μL模板DNA。反应程序:95℃预变性5min;95℃变性30s,54℃退火30s,72℃延伸45s,35个循环;72℃再延伸10min;4℃保存。以上反应均设置阴性对照以排除DNA污染。PCR产物经1.2%琼脂糖凝胶电泳、EB染色检测扩增产物的有无与量,酌量取PCR产物4~5μL以8%的非变性聚丙烯酰胺凝电泳,0.5×TBE电泳缓冲液,恒压120V(约2h)进行电泳、银染、显色、拍照。endprint
1.3.4SSR数据统计和分析结合软件AdobePhotoshopCS5和AlphaView凝胶图像分析软件分析微卫星条带大小,根据每个个体产生的条件位置确定基因型。用Popgene1.32软件进行分析,计算微卫星座位分别在2个群体中的等位基因数、期望杂合度、观测杂合度、平均基因多样性指数,并检验各个座位、各个群体的哈代一温伯格平衡。
2.结果与分析
2.1亚东鲑形态特征数据多元分析
如表2所示,对亚东鲑人工繁殖后代与野生群体的7项可数性状求平均值及标准差,经卡方检验,亚东鲑2个群体间的卡方值是x2=0.004;其侧线鳞数、侧线上鳞、侧线下鳞、背鳍条数、胸鳍条数、腹鳍条数、臀鳍条数相互差异均不显著;全长/体长、体长/体高、体长/头长、头长/吻长、头长/眼径、头长/眼间距、体长/尾柄长、尾柄长/尾柄高、背棘长/体长等9个比例性状的测量结果也不存在显著差异。
采用方差、判别和主成分等方法分析了2个群体包括7个可数、111个可量、20个框架数据的形态特征。结果表明,9个比例性状在野生和人工繁殖2个群体间差异均不显著,相似性状比例为100%;29个可量和框架数据在野生和人工繁殖两群体间差异均不显著,相似性状比例为100%。主成分分析得到因子1-11的方差累积贡献率在85%以上,但各自特征根极小,且对群体间总方差的贡献率小,无法用相互独立的因子来概括这2个群体鱼的形态差异。用逐步判别程序对经过校正的29个形态参数进行筛选,淘汰掉26个形态参数,得到D4-1/体长、眼径/体长、背棘长/体长等3个判别效果稍微显著的形态参数。计算得到2个群体判别准确率及互相证实准确率较低,分别为84.99%、85.04%(表3),即亚东鲑野生和人工繁殖后代2个群体易发生错判,也进一步证实了它们间不存在显著的形态差异。
2.2亚东鲑野生和人工繁殖群体的微卫星DNA多态性分析
采用9对微卫星引物扩增出10个多态性座位,获得55个等位基因,平均每个座位扩增出5.5个等位基因。不同的微卫星扩增出的等位基因数量不同,其中最多的是引物Strutta58,共扩增出10个等位基因,最少的是引物Str43INRA,共扩增出2个座位及5个等位基因(表4)。另外,不同微卫星引物在2个群体中可扩增出不同的基因数,如引物Strutta58在亚东鲑人工繁殖群体中扩增出8个等位基因,而在野生群体中扩增出5个等位基因,且有的个体中出现了无效等位基因。
从表5中可以看出,座位Str43INRA-1和Str43INRA-2在亚东鲑2个群体中均严重偏离哈代一温伯格平衡,其余8个座位在2个群体中均符合哈代一温伯格平衡。计算得亚东鲑人工繁殖群体和野生群体的平均等位基因数分别是4.8、4.0,平均观测杂合度分别是0.6267、0.6417,平均基因多样性指数均为0.5719。人工繁殖群体平均等位基因数多于野生群体,而人工繁殖群体平均观测杂合度略小于野生群体,但差异都不显著。
3.讨论与结论
3.1亚东鲑人工繁殖后代与野生群体的形态特征
亚东鲑人工繁殖后代与野生群体在形态学上差异不显著。在7个可数性状中,鳍式和鳞式都相似,没有差异;9个比例性状在两群体间差异均不显著;29个可量和框架数据在2个群体间的差异也不显著,相似性状比例为100%;主成分分析和判别分析也进一步证实了它们之间不存在显著性形态差异。亚东鲑渔场建于亚东河边并直接把河水引入池中进行流水养殖和繁育,养殖环境如水质、水温、光照、溶氧、饵料等与亚东河自然条件基本一致,2个群体在形态学上差异不显著,表明人工繁殖后代没有因人工繁殖和养殖操作产生形态遗传特征的改变。
3.2亚东鲑2个群体的微卫星DNA的多态性
微卫星无效等位基因普遍存在于微生物、动物、植物等众多物种中。微卫星侧翼序列变异、大片段等位基因丢失、不同微卫星位点对DNA质量要求差异等都是造成无效等位基因的原因。本研究所用微卫星引物都通过筛选所得,且不存在大片段等位基因丢失现象,因此主要原因可能是由于样品采集路途遥远、保存条件有限而导致DNA样品质量不太符合微卫星位点的要求。另外,微卫星无效等位基因具有隐匿性,且在杂合子个体中表现更为常见,这与本研究中人工繁殖群体杂合度比野生群体低相符,野生群体无效等位基因出现更多。因此,10个微卫星位点中8个位点都出现无效等位基因,导致各位点基因频率在两群体中具有一定的差异,但结合各位点杂合度等分析两群体遗传差异不明显。
引物Str43INRA扩增出2个位点为Str43IRNA-1、Str43IRNA-2。根据鲤鱼和草鱼相关报道可以推测,出现这种情况是因为亚东鲑曾经经历过特有的全基因组重复。杂合度、等位基因多样性和多态性位点的比例等是评估遗传多样性最常用的指标。本研究发现,亚东鲑野生群体和人工繁殖后代中的微卫星杂合子均严重缺乏,位点$sa85在2个群体均严重偏离哈代一温伯格平衡,人工选择、迁移、突变、近交等都有可能产生该现象。亚东鲑最早引入亚东河并栖息下来时的个体数量少,导致遗传瓶颈和漂变的发生可能是杂合子急剧减少和检测到偏离哈代一温伯格平衡的主要原因。本研究表明,亚东鲑人工繁殖群体的平均等位基因数大于野生群体,也从不同角度证明亚东鲑野生资源正在经历一个急剧衰退的过程,需要加强亚东鲑的保护性繁育和增殖。
豪富华等报道亚东鲑野生群体的平均观测杂合度分别是0.7083,而本研究的亚东鲑野生群体、人工繁殖后代的观测杂合度为0.6417、0.6267,均显著低于2006年所取的亚东鲑野生群体的杂合度,这说明亚东鲑种群的遗传多样性处于下降趋势。同时,亚东鲑人工繁殖群体的遗传杂合度略小于野生群体,但2个群体遗传杂合度差异不显著,说明亚东鲑渔场多年来陆续收集和培育的野生亲本群体基本保留了目前野生群体的遗传多样性,应可用于野生资源的恢复和增殖工作。endprint
猜你喜欢
东方教育(2016年9期)2017-01-17
法制与社会(2016年33期)2016-12-15
新闻世界(2016年11期)2016-12-10
现代园艺(2016年4期)2016-03-24
现代园艺(2016年4期)2016-03-24
现代园艺(2016年3期)2016-03-24
现代园艺(2016年2期)2016-03-15
现代园艺(2016年1期)2016-02-23
纺织导报(2015年2期)2015-04-07
