
暖温带-中温带过渡区4种典型森林土壤呼吸的温度敏感性
2015-12-08 06:26张慧东尤文忠魏文俊周梅
生态环境学报 2015年11期
张慧东,尤文忠,魏文俊,周梅
1. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2. 辽宁省林业科学研究院,辽宁 沈阳 110032
暖温带-中温带过渡区4种典型森林土壤呼吸的温度敏感性
张慧东1,2,尤文忠2,魏文俊2,周梅1*
1. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2. 辽宁省林业科学研究院,辽宁 沈阳 110032
温度是全球气候变化最重要的生态因子,过渡区生态系统的能量流动和物质循环过程极易受到气候变化的影响。为揭示暖温带-中温带过渡区森林土壤呼吸对温度变化的响应,选择在暖温带-中温带过渡区分布面积较大的长白落叶松(Larix olgensis)、红松(Pinus koraiensis)、油松(Pinus tabuliformis)人工林和天然阔叶混交林4种森林类型,利用Li-8100红外气体分析仪于2010─2013年连续观测土壤呼吸速率,同时利用森林小气候梯度观测系统连续同步观测大气温度、大气降水和土壤温度等环境因子,系统研究了土壤呼吸速率动态及其温度敏感性。结果表明:长白落叶松、红松、油松和阔叶混交林的年平均土壤呼吸速率(Rs,以CO2计)分别为(2.31±0.01)、(2.07±0.71)、(1.55±0.03)和(2.24±0.02)μmol·m2·s-1;4种森林类型的Rs与10 cm土壤温度(t10)均具有极显著的相关关系(P=0.000<0.01);生长季期间落叶松、红松、油松人工林和天然阔叶混交林的Q10值分别为3.32、4.46、4.12和3.59,其中红松人工林Rs对t10的温度变化最敏感,而天然阔叶林和落叶松人工林的敏感性相对较低。本研究还对非生长季(11月─翌年3月)期间长白落叶松人工林的土壤呼吸进行了连续监测,并依据非生长季期间土壤温度的变化,将非生长季期间的土壤呼吸分为冻结期(t10-mean:10.0~2.0 ℃)、冻融期(t10-mean:0.5~2.0 ℃)和非冻结期(t10-mean:>2.0 ℃)3个阶段。结果显示:长白落叶松人工林在非生长季期间仍有微弱的呼吸作用(以CO2计,0.01~1.38 μmol·m2·s-1),整个非生长季期间Rs与t10有极显著相关关系(r2=0.586 3,P=0.000<0.01),非生长季冻结期的Rs与t10无显著相关关系(P=0.503>0.5),冻融期的Rs变化较剧烈,且较冻结期有较明显的增加;整个非生长季落叶松人工林的Q10值为4.65,是生长季的1.40倍。研究结果对进一步阐明气候带过渡区不同森林类型土壤呼吸对气候变化的响应具有重要意义。
土壤呼吸;温度敏感性;森林;东北地区
ZHANG Huidong, YOU Wenzhong, WEI Wenjun, ZHOU Mei. Temperature Sensitivity of Soil Respiration in Four Forest Types in Temperate Ecotone, Northeast China [J]. Ecology and Environmental Sciences, 2015, 24(11): 1757-1764.
土壤CO2通量作为陆地生态系统CO2通量第二大组成部分,在全球碳循环中扮演着极其重要的角色(Zhou et al.,2009;Luan et al.,2011),占整个陆地生态系统呼吸的 50%~75%(Deforest et al.,2006),土壤碳库的任何细微变化都会对大气 CO2含量产生显著影响(Schimel et al.,1990;Jenkinson et al.,1991)。已有大量的研究表明土壤温度和土壤水分是影响森林土壤呼吸的重要非生物因素(Jenkinson et al.,1991;Hanson et al.,2004;朱教君等,2008;杨阔等,2010),阐明典型森林生态系统土壤呼吸对土壤水热变化的响应和适应规律,对揭示碳收支平衡机理具有重要意义(Burton et al.,2003)。多数研究者认为:土壤温度是与土壤呼吸关系最密切的非生物因子,在一定的温度范围内二者呈现正相关关系(方精云等,2001;Liu et al.,2002);但也有研究认为,土壤温度对土壤呼吸的影响不显著,二者相关性较弱(闫敏华等,2001);Hanson et al.(2004)通过对全球13个不同生态类型生态过程的模拟发现,土壤呼吸速率与土壤温度及水分的相关系数r2在0.04~0.64之间。可见,陆地生态系统土壤呼吸对温度变化的响应存在着很大的不确定性。全球气候变化主要通过影响土壤温度和含水量来影响土壤呼吸(Maier et al.,2000),过渡区生态系统对全球气候变化影响极为敏感,生态环境极脆弱,森林植被极易受到气候变化的影响。因此,开展暖温带与中温带过渡区不同森林类型土壤CO2通量对温度敏感性的响应,对于准确评估土壤碳固存对生态系统碳收支平衡的贡献和科学预测生态系统碳循环对全球变化的响应具有极其重要的意义。
目前东北地区关于森林土壤呼吸已进行了较多的研究,包括了小兴安岭、长白山等区域的蒙古栎(Quercus mongolica)林、兴安落叶松(Larix gmelinii)林、长白落叶松(Larix olgensis)林、红松(Pinus koraiensis)林、云冷杉(Picea asperata-Abies fabri)林、硬阔叶混交林、杨桦(Populus davidiana-Betula platyphylla)林、杂木林等主要森林类型(You et al.2013;史宝库等,2012;张慧东等,2011;杨阔等,2010;杨金艳等,2006;朱教君等,2008),对不同森林类型土壤呼吸的特征、组成、影响因素及其对气候变化的响应等进行了较全面的研究。但是,对温度敏感性响应较强烈的暖温带-中温带过渡区典型森林的土壤呼吸与温度敏感性关系的研究还较少,尤其是缺乏长时间尺度的定位观测研究。这给区域森林土壤碳库的估算带来了较大的不确定性,因此开展东北森林生态系统的土壤呼吸及其与土壤温度的关系的研究,不但直接关系到本地区森林生态系统碳循环的源汇强度的评估,而且关系到全球变化条件下该生态系统碳循环的预测。本文研究了东北地区4种典型的森林类型的土壤呼吸对温度的敏感性的研究,对提高温带森林土壤生态学过程的理解具有重要的意义,同时能为更好地评价和预测气候变化对不同森林生态系统碳循环的影响提供科学依据。
1 材料与方法
1.1研究区概况及样地选择
本研究在位于辽宁省西丰县境内的国家林业局所属辽宁冰砬山森林生态系统国家定位观测研究站(N42°33′37″,E125°03′15″,以下简称冰砬山森林生态站)进行。该地区属温带大陆性气候,春季气温回升迅速,夏季雨量集中,秋霜较早,冬季寒冷。7月份气温最高,平均 23.2 ℃,极端高温35.2 ℃;1月份气温最低,平均-17.2 ℃,极端低温-41.1 ℃;具有典型的山区气候特征,年均气温5.2 ℃,年均降水量684.8 mm,年均蒸发量1379.8 mm,无霜期133 d,是我国暖温带-中温带过渡区。
该区地貌属低山丘陵,平均海拔500~600 m,最高峰海拔870 m。土壤以暗棕色森林土为主,其次为棕色森林土,土壤质地多为粉沙壤质、壤质,土层深厚,有机质含量高,pH值6~7。原生植被为红松阔叶林,现已被次生阔叶混交林、长白落叶松人工林、日本落叶松(Larixkaempferi)人工林、红松人工林、油松(Pinus tabuliformis)人工林等所取代。
于2010年7月,分别在长白落叶松人工林、红松人工林、油松人工林和天然阔叶混交林中选择坡面平坦、均质性强的坡中地段各建立 3块 20 m×30 m观测样地,对每块样地进行每木检尺,样地基本情况见表 1。天然阔叶混交林主要由核桃楸(Juglans mandshurica)、黄菠萝(Phellodendron amurense)、枫桦(Betulacostata)等优势树种组成,林冠下有槭属(Acer)、榆属(Ulmus.)等伴生树种,灌木主要有接骨木(Sambucus williamsii)、稠李(Prunuspadus)、卫矛(Euonymus alatus)、珍珠梅(Sorbaria sorbifolia)等,草本主要有白花碎米荠( Cardamine leucantha)、 猴 腿 蹄 盖 蕨(Athyriummultidentatum) 、悬钩子(Rubuscorchorifolius)、白屈菜(Chelidonium majus)、堇菜(Viola verecunda)等,盖度85%。长白落叶松林、红松林和油松林均为人工纯林,林龄46~60 a,郁闭度0.8以上,林下灌木稀少,草本主要为羊胡苔草(Carex rigescens),盖度85%。4种森林类型土壤情况见表2。
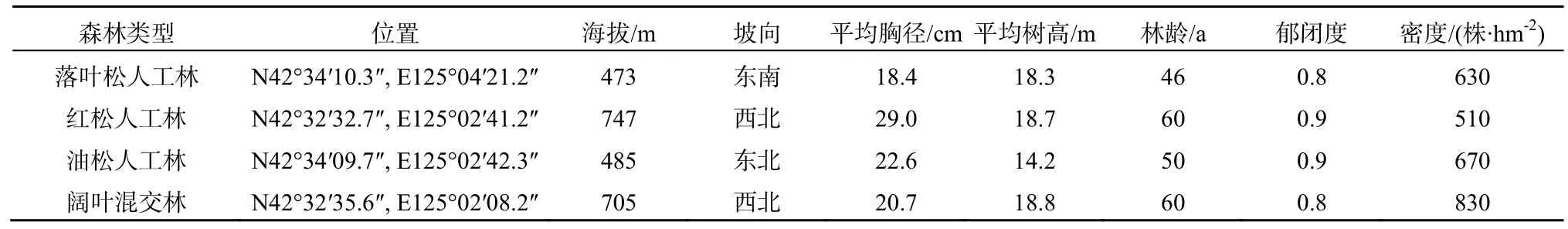
表1 4种森林类型基本情况调查Table 1 General situation of 4 forests
1.2土壤呼吸测定
土壤呼吸速率(Rs)采用 Li-8100(Li-Cor,USA)红外气体分析仪测定。在样地内沿样地对角线等距布设观测样点3个,然后在每个样点设置3个直径21.34 cm、高11.43 cm的PVC土壤隔离环,将PVC环前端削薄插入土壤,地上保留1.0 cm,尽量保持环内的土壤表层枯落物不受到扰动。落叶松人工林土壤呼吸环在2007年9月安放,红松人工林、油松人工林和阔叶次生林土壤呼吸环在 2010年7月初安放。每次测量之前,土壤隔离环内的草本用剪刀紧贴地表剪除。本实验根据研究区气候特征,将土壤解冻植物开始复苏萌芽至秋冬季节各种落叶植物落叶休眠的时间段定义为生长季(4月下旬─10月下旬),将落叶植物休眠至翌年土壤解冻植物开始复苏萌芽前的时间段定义为树木的非生长季(11月初─翌年4月中旬)。2010年7月─2013年7月,生长季期间的每月上、中和下旬选择晴好天气分别对落叶松、红松、油松和阔叶混交林进行土壤呼吸测定。非生长季期间的每月上、中和下旬继续对落叶松跟踪测定土壤呼吸速率。Li-8100红外气体分析仪对观测的土壤呼吸数值自动记录,每个样点重复3次,每次测量的循环周期为生长季5 min、非生长季7 min。
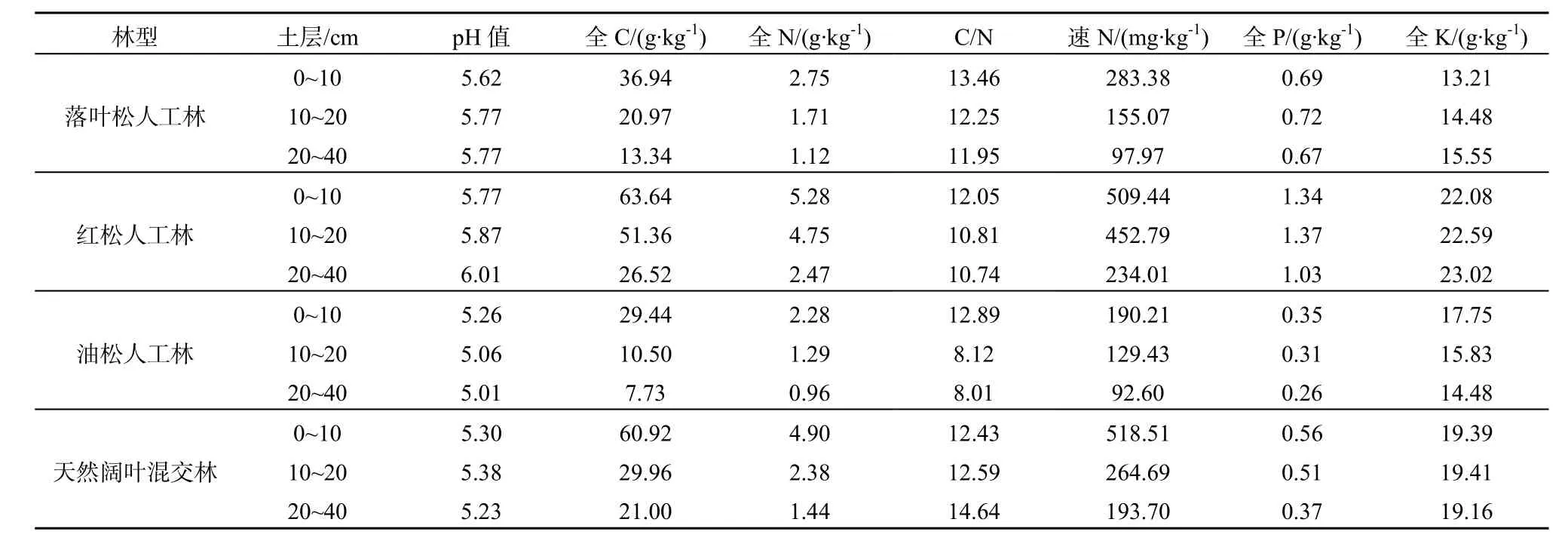
表2 4种森林类型土壤养分状况Table 2 Soil nutrient conditions of 4 forests
1.3环境因子测定
在测量土壤呼吸的同时利用Li-8100附带的土壤温度传感器和土壤湿度传感器在样点附近测量10 cm深土壤的温度(t10);同时,在观测期间利用冰砬山森林生态站森林小气候梯度观测系统对研究区的林内大气温度、相对湿度、大气降水和 5、10、15 cm的土壤温度等进行连续测量。样地土壤理化性质数据来源于2011年固定样地调查数据。
1.4数据分析
对不同森林类型生长季的土壤呼吸速率、温度和湿度的显著性检验,采用重复测量方差分析(Repeat measured ANOVA)进行方差分析和多重比较。用线性和非线性方程拟合土壤呼吸(Rs)与土壤温度(t)之间的单因素和多因素关系。利用土壤呼吸和土壤温度指数关系方程(Rs=aebt)中的温度反应系数b计算土壤呼吸的温度敏感性指数Q10,其定义为Q10=e10b(黄耀,2003)。
2 结果分析
2.1土壤呼吸与环境因子
2010年7月至2013年7月期间土壤呼吸的年际动态变化与土壤温度、大气温度和大气降水的动态变化基本一致,季节变化表现为单峰型;生长季期间研究区内的平均土壤体积含水量变化较小(图1)。在2010─2013年生长季期间4种森林类型的平均土壤呼吸速率(以 CO2计,下同)分别为(2.31±0.01)、(2.07±0.71)、(1.55±0.03)和(2.24±0.02)μmol·m2·s-1。如图1所示,在2011年完整的生长季期间,落叶松、红松、油松人工林和天然阔叶林Rs-max分别出现在6月29日、7月29日、8月 3日和 7月 7日,分别为(5.07±0.12)、(4.45±0.12)、(5.27±0.12)和(3.78±0.12)μmol·m2·s-1;而在 2012年完整的生长季期间,落叶松、红松、油松人工林和天然阔叶林Rs-max分别出现在8月20日、8月6日、8月10日和6月24日,分别为(5.04±0.12)、(4.53±0.13)、(3.39±0.12)和(4.08±0.12)μmol·m2·s-1。
4种森林类型生长季土壤呼吸的年平均速率方差分析结果显示(图2),4种森林类型的土壤呼吸平均速率存在显著性差异(P=0.003,n=12),其中落叶松人工林与红松人工林存在显著差异(P=0.038,n=6),与油松人工林存在极显著差异(P=0.001,n=6),与阔叶混交林无显著差异(P=0.697,n=6;);红松人工林与油松人工林和阔叶混交林均无显著差异(P分别为0.066和0.076,均大于0.05,n=6);油松人工林与红松人工林无显著性差异(P=0.066,n=6),而与落叶松人工林存在极显著差异(P=0.001,n=6),与阔叶混交林存在显著差异(P=0.002,n=6)。
2.2生长季土壤呼吸Rs的温度敏感性
在生长季期间,4种森林类型的土壤呼吸速率Rs与 t10均存在极显著的相关性(P<0.01),Rs-t10指数模型的拟合相关系数 r2分别为落叶松 0.8090(P=0.000<0.01)、红松0.8662(P=0.000<0.01)、油松 0.9347(P=0.000<0.01)和阔叶混交林 0.8090(P=0.000<0.01)(图3)。
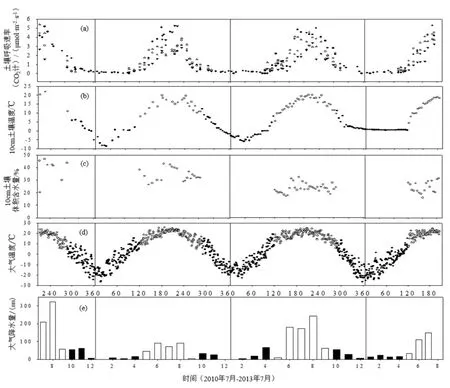
图1 4种森林类型的土壤呼吸速率及其主要环境因子动态变化Fig. 1 Dynamics of soil respiration rate and the major environmental factors
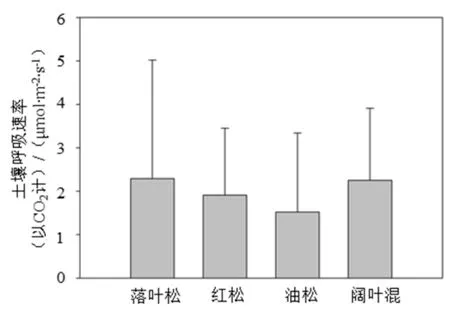
图2 2010—2013年生长季期间4种森林类型平均土壤呼吸速率特征Fig. 2 Characteristics of mean soil respiration rate for 4 forests in growing season during 2010 and 2013
Q10能够反映出不同森林类型对温度变化的敏感性响应,以4种森林类型的Rs-t10指数模型为基础,对反映Rs的温度敏感性指标Q10值进行计算,结果为红松人工林(4.46)>油松人工林(4.12)>天阔叶混交林(3.59)>落叶松人工林(3.32);生长季期间,4种森林类型的Q10值平均为3.87,不同森林类型的Q10的变异系数为0.264。由此看出,不同森林类型对温度的敏感性响应具有显著差异,表现为红松人工林的 Rs对气候变化的土壤温度变化最敏感,其次是油松人工林,而天然阔叶林和落叶松人工林对气候变化的敏感性最弱。
2.3非生长季森林土壤呼吸的温度敏感性
非生长季土壤碳通量是年土壤碳通量的基本组成部分。非生长季落叶松人工林10 cm土壤温度t10保持在-10~10 ℃之间;Rs CO2在 0.01~1.38 μmol·m2·s-1之间变化,Rs与 t10呈指数相关关系(RS=0.2496e0.153 7t10,r2=0.5863,P=0.000<0.01)(图4)。根据土壤表层温度的变化情况,将非生长季期间Rs分为3个阶段:Ⅰ冻结期,即t10-mean保持在-10~0.5 ℃之间,此时落叶松人工林的Rs值保持在0.12~0.24 μmol·m2·s-1间波动,Rs与t10无显著的相关性(P=0.503>0.5,n=40);Ⅱ冻融期:即 t10-mean在0.5~2.0 ℃间,此时Rs较冻结期明显升高,最高可达CO20.73 μmol·m2·s-1,可达冻结期平均Rs的3倍以上,Rs与 t10相关性不显著(P=0.896>0.5,n=20);Ⅲ非冻结期:即t10-mean>2 ℃以后,表层土壤完全解冻,此时Rs与t10的相关性达到极显著,土壤呼吸Rs随t10呈指数函数增长(P=0.000<0.01,n=48)(图4)。
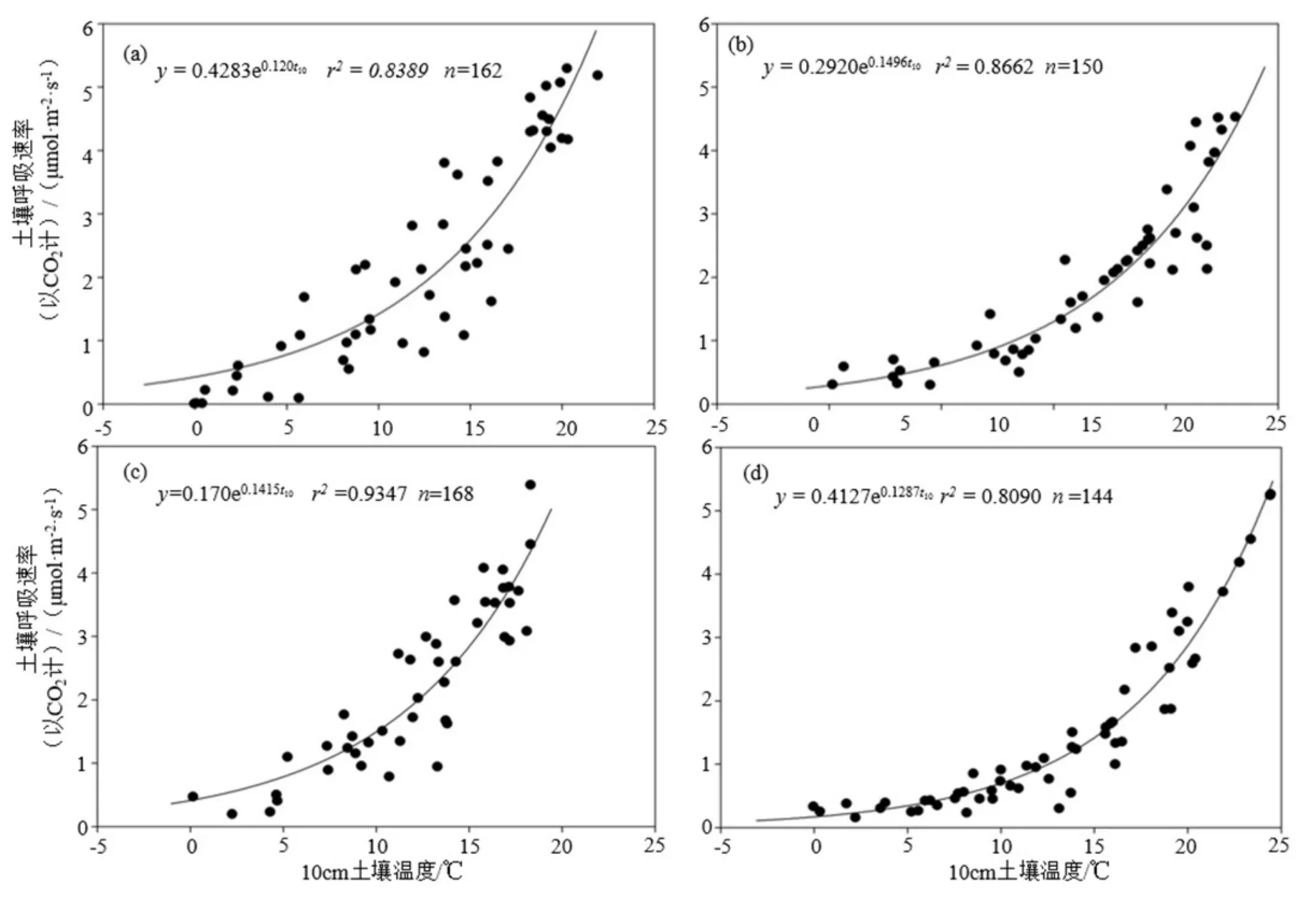
图3 生长季4种森林类型土壤呼吸速率(Rs)对10 cm土壤温度(t10)的响应Fig. 3 Response of soil temperature at the depth of 10 cm on soil respiration rate in 4 forests
通过对落叶松人工林Rs连续3年的观测,根据生长季、非生长季和完整生长季的土壤呼吸速率Rs与 t10分别建立 Rsg-t10、Rsd-t10和 Rsg+d-t103个Rs-t10模型(见图3a、图4左和图4右)。根据建立的模型对落叶松人工林生长季、非生长季和全年的Q10值进行了计算,结果显示:落叶松人工林在生长季、非生长季和全年的 Q10值分别为 3.32、4.65和3.64,生长季的Q10最低,非生长季的Q10是生长季的1.40倍。
3 讨论
3.1不同森林类型Rs特征及对土壤温度的响应
有研究认为,林分的结构特征是影响土壤呼吸季节动态变化的重要因子(Humphreysa et al.,2006;Wang et al.,2006)。本研究中4种森林类型的土壤呼吸速率在生长季期间表现差异较大,落叶松、红松、油松、阔叶混交林 4种森林类型的年平均 Rs分别为 (2.31±0.01)、(2.07±0.71)、(1.55±0.03)、(2.24±0.02) μmol·m2·s-1。本研究结果与朱教君等(2008)、Matzner et al.(2008)、杨阔等(2010)、You et al.(2013)对东北地区不同森林类型的土壤呼吸速率的研究结果基本一致。
多数研究表明,森林土壤呼吸与土壤表层温度呈现良好的相关性(Fahnestock et al.,1999;Mikan et al.,2002;Wu et al.,2010)。土壤呼吸速率的季节变化通常与温度变化密切相关,在土壤湿度不是限制因子的条件下,土壤温度可以解释土壤呼吸速率变异的60%~80%(Matzner et al.,2008)。本文的研究结果也与多数研究的结果相一致,4种森林的Rs与土壤表层10 cm深的温度具有极显著相关关系(P<0.01),并且呈指数函数关系,相关系数r2均达到0.8以上。大量的研究结果显示,表征森林土壤呼吸对温度敏感性的Q10值在时间和空间上存在着巨大的变异,并且随生态系统的类型、地理位置和环境因子等影响因素的变化而变化(You et al.,2013;Pavelka et al.,2007;Zhou et al.,2009)。本研究结果表明Q10值在不同森林类型间有显著的差异,而且在同一森林类型的不同季节土壤呼吸对温度的敏感性也不一致。因此,应在不同空间和时间尺度上进一步丰富森林 Rs对气候变化,尤其是温度敏感性的研究,提高对森林生态系统在应对气候变化中的作用。
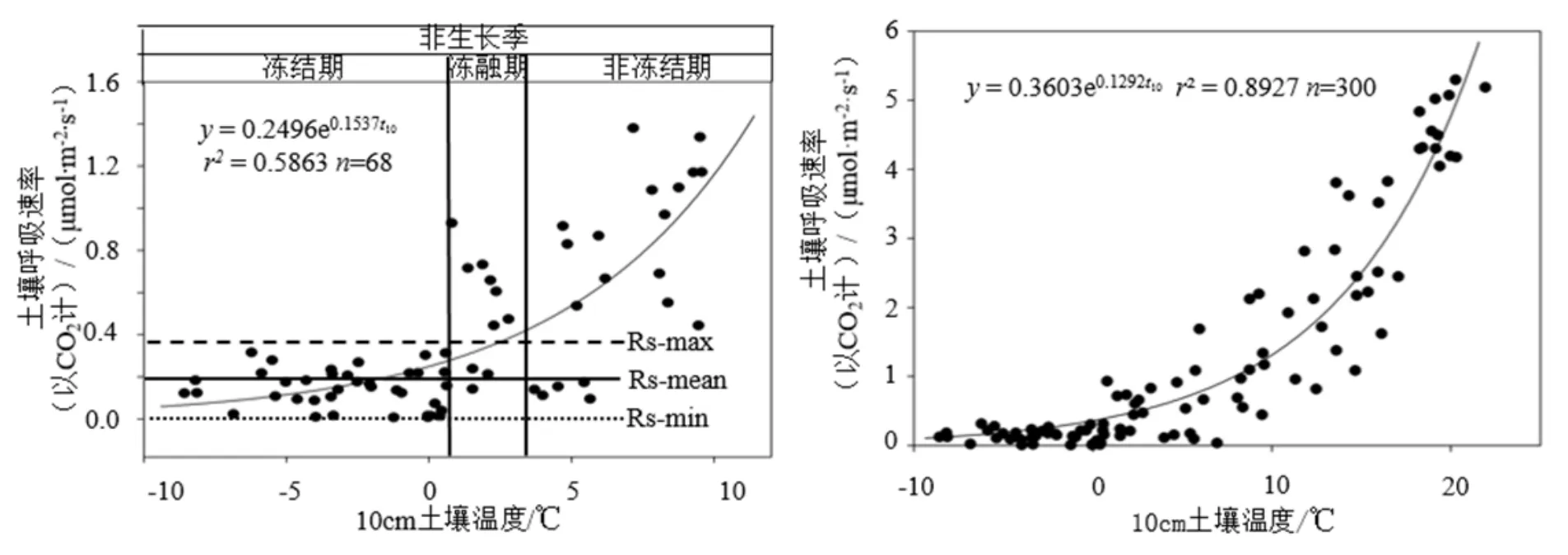
图4 非生长季和全年落叶松人工林Rs与t10相关关系Fig. 4 The relationship between soil respiration rate and soil temperature at depth of 10 cm in Larch plantation during nongrowing season and in the whole year
3.2非生长季Rs特征及对土壤温度的敏感性
森林非生长季土壤CO2通量是区域碳收支非常重要的组成部分,大量的科学研究证实了中、高纬度和高海拔低温生态系统非生长季仍然存在着相当数量的呼吸作用(Fahnestock et al.,1999;Grogan et al.,2006;Schindlbacher et al.,2007),非生长季生物活动和生物地球化学碳循环仍然十分活跃并且发挥着不可忽视的作用。尤其冻融交替期作为季节性冻土区非生长季内经历的一个重要时期,其CO2排放量可占全年土壤 CO2通量的 3%~50%(Liptzin et al.,2009;Lafleur et al.,2001;Wickland et al.,2001)。土壤冻融交替对土壤CO2的排放有不可忽视的影响(Wu et al.,2010;王琼等,2010)。在东北地区非生长季期间的 11─12月的土壤冻结期,t10随时间的变化表现为逐渐降低,日变化不明显;而在翌年4月生长前期的解冻期,t10的日变化比较明显。如图4a所示,冻融期Rs峰值出现在2013年 4月 11日,当日 10 cm土壤温度的日变化为-0.007~2.98 ℃,平均土壤温度为0.70 ℃时,Rs突然增大到0.73 μmol·m2·s-1,是冻结期平均土壤呼吸速率的3倍。本研究结果也表明,温带地区森林土壤在非生长季期间仍保持着微弱的土壤呼吸,在不同的阶段表现不同;同时通过对整个非生长季期间土壤呼吸的分段解析发现,在土壤冻融期,森林土壤的CO2通量存在增大的现象,这一结果不仅进一步证明落叶松人工林在非生长季的 Rs对气候变化的响应更敏感,而且说明如果仅用生长季的Q10分析气候变化对森林土壤呼吸的影响,将会低估森林对气候变化的敏感性,约低9.64%。因此,在温带地区开展非生长季土壤呼吸研究对于准确评估森林生态系统的碳循环及其森林土壤呼吸对气候变化的响应具有重要意义。这一研究结果与国内外的一些同样在野外原位进行的观测结果一致(Mikan et al.,2002;Skogland et al.,1988;Priemé et al.,2001;Bubier et al.,2002;Kato et al.,2005;宋长春等,2005)。冻融循环的次数、强度(Decker et al.,2003;Matzner et al.,2008)可对冻融过程产生影响,进而改变土壤水热性质导致土壤的生物地球化学过程速率发生变化(Matzner et al.,2008;程慧艳等,2008;Grogan et al.,2004;孙辉等,2008;王娇月等,2011)。为了进一步解析冻融期间Rs的动态变化情况,对冻融期的土壤温度日变化进行分析,研究发现表层土壤的冻融过程受春季气温变化的影响非常短暂,在2013年春季仅发生在4月的10─11日,之后t10的日变化均维持在0 ℃以上,这给开展冻融期土壤呼吸特征的研究带来了较大的难度。
4 结论
通过对东北地区4种森林类型土壤呼吸及土壤温度进行的连续3年观测和分析发现:
(1)该研究区域的4种森林类型土壤呼吸的年际动态变化与土壤温度、大气温度的动态变化基本一致,季节变化表现为单峰型,且4种森林类型的平均Rs存在显著性差异。
(2)生长季期间4种森林类型的Rs与t10极显著相关,Rs-t10指数模型可以很好地解释该地区 4种森林类型土壤呼吸的变化情况。4种森林类型的Rs温度敏感性Q10值显示不同森林类型对温度的敏感性响应具有显著差异,表现为红松人工林 Rs对土壤温度变化最敏感,天然阔叶混交林和落叶松人工林在应对气候变化方面的能力相对较强。
(3)通过对非生长季落叶松人工林Rs的观测,进一步证实了中、高纬度森林在非生长季的土壤CO2通量是该区域土壤碳收支的重要组成部分,并通过实测数据将非生长季Rs的动态变化分为冻结、冻融和非冻结3个阶段。对落叶松人工林非生长季、生长季和全年3种情况的Rs温度敏感性分析结果表明,落叶松人工林的Q10值在生长季最低,非生长季的Q10值最高,是生长季的1.40倍,仅用生长季的Q10对不同森林类型的温度敏感性进行解释将低估Rs对温度变化的敏感性。
由于受研究条件、环境条件等因素的限制,目前只能从宏观上对不同森林类型 Rs在生长季和非生长季期间的 Rs变化进行初步的解释。在今后的研究中还应继续加强对非生长季不同森林类型 Rs的研究,进一步完善中、高纬度和高海拔区域 Rs的时空动态变化规律,特别是要从机理、机制方面进一步揭示冻融期森林Rs的变化特征。
BOWDEN R D, NEWKIRK K M, RULLO G M. 1998. Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions [J]. Soil Biology and Biochemistry, 30(12): 1591-1597.
BUBIER J, CRILL P, MOSEDALE A. 2002. Net ecosystem CO2exchange measured by auto chambers during the snow-covered season at a temperate peatland [J]. Hydrological Processes, 16(18): 3667-3682.
COX P M, BETTS R A, JONES C D, et al. 2000. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model [J]. Nature, 408(6809): 184-187.
DEFOREST J L, NOOMETS A, MCNULTY S G, et al. 2006. Phenophases alter the soil respiration - temperature relationship in an oak -dominated forest [J]. International journal of biometeorology, 51(2): 135-144.
FAHNESTOCK J T, JONES M H, WELKER J M. 1999. Wintertime CO2efflux from arctic soils: Implications for annual carbon budgets [J]. Global Biogeochemical Cycles, 13(3): 775-779.
GROGAN P, JONASSON S. 2006. Ecosystem CO2production during winter in a Swedish subarctic region: the relative importance of climate and vegetation type [J]. Global Change Biology, 12(8): 1479-1495.
GROGAN P, MICHELSEN A, AMBUS P, et al. 2004. Freeze-thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms [J]. Soil Biology and Biochemistry, 36(4): 641-654.
HANSON P J, AMTHOR J S, WULLSCHLEGER S D, et al. 2004. Oak forest carbon and water simulations: Model intercomparisons and evaluations against independent data [J]. Ecological Monographs, 74(3): 443-489.
HUMPHREYSA E R, ANDREW BLACKA T, MORGENSTERNA K, et al. 2006. Carbon dioxide fluxes in coastal Douglas-fir stands at different stages of development after clearcutclear cut harvesting [J]. Agricultural and Forest Meteorology, 140(1-4): 6-22.
JENKINSON D S, ADAMS D E, WILD A. 1991. Model estimates of CO2emissions from soil in response to global warming [J]. Nature, 351: 304-306.
KATO T, HIROTA M, TANG Y, et al. 2005. Strong temperature dependence and no moss photosynthesis in winter CO2flux for a Kobresia meadow on the Qinghai Tibetan plateau [J]. Soil Biology and Biochemistry, 37(10): 1966-1969.
LAFLEUR PM, POULET NT, ADMIRAL SW. 2001. Annual cycle of CO2exchange at a bog peatlandpeat land [J]. Journal of Geophysical Research, 106(D3): 3071-3081.
保教工作是一项系统的工作,绝不是简单地在幼儿园开展几项活动就可以实现的。因此,应该强化家园合作,努力提升幼儿的保教质量。如果园区所在社区具备较为丰富的保健资源,那么社区则能够为幼儿园保教工作的开展提供必要的支持。通过将家庭、社区和幼儿园进行有机结合,能够使幼儿园保教工作更为高效地开展,能够更有计划性、更合理地对幼儿进行必要的科学保教,从而让幼儿园一日活动科学保教更能满足幼儿成长的需要。
LIPTZIN D, WILLIAMS M W, HELMIG D, et al. 2009. Process-level controls on CO2fluxes from a seasonally snow-covered subalpine meadow soil, Niwot Ridge, Colorado [J]. Biogeochemistry, 95(1):151-166.
MAIER C A, KRESS L W. 2000.Soil CO2evolution and root respiration in 11 year-old loblolly pine plantations as affected by moisture and nutrient availability [J]. Canadian Journal of Forest Research, 30(3): 347-359
MATZNER E, BORKEN W. 2008. Do freeze-thaw events enhance C and N losses from soils of different ecosystems? A review [J]. European Journal of Soil Science, European Journal of Soil Science, 59(2): 274-284.
MIKAN C J, SCHIMEL J P, DOULE A P. 2002. Temperature controls of microbial respiration in arctic tundra soils above and below freezing [J]. Soil Biology and Biochemistry, 34(11): 1785-1795.
PAVELKA M, ACOSTA M, MAREK M V, et al. 2007. Dependence of the Q10values on the depth of the soil temperature measuring point [J]. Plant and Soil, 292(1-2): 171-179.
PRIEMÉ A, CHRISTENSEN S. 2001. Natural perturbations, drying-wetting and freezing-thawing cycles, and the emission of nitrous oxide, carbon dioxide and methane from farmed organic soils [J]. Soil Biology and Biochemistry, 33(15): 2083-2091.
SCHIMEL D S, PARTON W J, KITTEL T G F, et al. 1990. Grassland biogeochemistry: Links to atmospheric processes [J]. Climatic change, 17(1): 13-25.
SCHINDLBACHER A, BOLTENSTERN S Z, GLATZEL G, et al. 2007. Winter soil respiration from an Austrian mountain forest [J]. Agricultural and Forest Meteorology, 146(3-4): 205-215.
SKOGLAND T, LOMELAND S, GOKSØYR J. 1988. Respiratory burst after freezing and thawing of soil: experiments with soil bacteria [J]. Soil Biology and Biochemistry, 20(6): 851-856.
WANG C K, YANG J Y, ZHANG Q Z. 2006. Soil respiration in six temperate forests in China [J]. Global Change Biology, 12(11): 2103-2114.
WICKLAND K P, STRIEGL R G, MAST M A, et al. 2001. Carbon gas exchange at a southern Roky Mountain wetland, 1996-1998 [J]. Global biochemical cycles, 15(2): 321-335.
WU X, BRGGEMANN N, GASCHE R, et al. 2010. Environmental controls over soil atmosphere exchange of N2O, NO, NO and CO2in a temperate Norway spruce forest [J]. Global Biogeochemical Cycles, 24(2): 1-16.
YOU W Z, WEI W J, ZHANG H D, et al. 2013. Temporal patterns of soil CO2efflux in a temperate Korean Larch (Larix olgensis Herry.) plantation, Northeast China [J]. Trees, 27(5): 1417-1428.
ZHANG J T. 1998. Effects of global climate change on C and N circulation in natural soils [J]. Scientia Geographica Sinica, 18: 463-471.
ZHOU T, SHI P J, HUI D F, et al. 2009. Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implicationsfor carbon-climate feedback [J]. Journal of Geophysical Research, 114(G2): 1-9.
LUAN J W, LIU S R, WANG J X, et al. 2011. Rhizospheric and heterotrophic respiration of a warm-temperate oak chronosequence in China [J]. Soil Biology and Biochemistry, 43,503-512.
BURTON A J, PREGITZER K S. 2003.Field measurements of root respiration indicate little to no seasonal temperature acclimation for sugar maple and red pine [J]. Tree Physiology, 23: 273-280.
LIU X Z, WAN S Q, SU B, et al. 2002. Response of soil CO2 efflux to water manipulation in a tall grass prairie ecosystem [J]. Plant and Soil, 240: 213-223
程慧艳, 王根绪, 王一博, 等. 2008. 黄河源区不同植被类型覆盖下季节冻土冻融过程中的土壤温湿空间变化[J].兰州大学学报(自然科学版), 44(2): 15-21.
方精云, 朴世龙, 赵淑清. 2001.CO2失汇与北半球中高纬度陆地生态系统的碳汇[J].植物生态学报, 25( 5): 594-602.
黄耀. 2003. 地气系统碳氮交换:从实验到模型[M]. 北京: 气象出版社.
史宝库, 金光泽, 汪兆洋. 2012. 小兴安岭5种林型土壤呼吸时空变异[J].生态学报, 32(17): 5416-5428.
宋长春, 王毅勇, 王跃思, 等. 2005. 季节性冻融期沼泽湿地CO2、CH4和N2O排放动态[J]. 环境科学, 26(4): 7-12.
孙辉, 秦纪洪, 吴杨. 2008. 土壤冻融交替生态效应研究进展[J]. 土壤, 40(4): 505-509.
王娇月, 宋长春, 王宪伟, 等. 2011. 冻融作用对土壤有机碳库及微生物的影响研究进展[J]. 冰川冻土, 33(2): 442-452.
王琼, 范志平, 王满贵, 等. 2010. 冻融交替对科尔沁沙地不同土地利用方式土壤呼吸的影响[J]. 生态学杂志, 29(7): 1333-1339.
闫敏华, 邓伟, 马学慧. 2001. 大面积开荒扰动下的三江平原近45年气候变化[J].地理学报, 56( 2): 159-169.
杨金艳, 王传宽. 2006. 土壤水热条件对东北森林土壤表面CO2通量的影响[J]. 植物生态学报, 30(2): 286-294.
杨阔, 王传宽, 焦振. 2010. 东北东部5种温带森林的春季土壤呼吸[J].生态学报, 30(12): 3155-3162.
张慧东, 尤文忠, 邢兆凯, 等. 2011. 辽东山区天然次生林和落叶松人工林的土壤呼吸[J]. 水土保持学报, 25(3): 198-201, 224.
朱教君, 闫巧林, 范阿南. 2008. 东北山区次生林生态系统土壤碳通量研究[C]//第五届中国青年生态学工作者学术研讨会论文集: 112-120.
Temperature Sensitivity of Soil Respiration in Four Forest Types in Temperate Ecotone, Northeast China
ZHANG Huidong1,2, YOU Wenzhong2, WEI Wenjun2, ZHOU Mei1
1. College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Hohhot 010019, China; 2. Liaoning Academy of Forestry, Shenyang 110032, China
Soil carbon efflux in forests is a significant component in the terrestrial carbon cycle. Temperature is the major factor affecting forest soil respiration and very important in the context of global climate change in ecosystems in transition zones are vulnerable to the effects of climate change. Larch (Larix olgensis), Korean pine (Pinus koraiensis), Chinese pine (Pinus tabuliformis) plantations and natural broadleaved mixed forests, which are the major forest types in Northeast China, play important roles in forest ecosystem carbon cycles in China. In this study, a Li-8100 infrared gas analyzer was used to observe the soil respiration rate (Rs) from 2010 to 2013, and a forest gradient microclimate observation system was applied to continuously observe the air temperature, precipitation, soil temperature and other environmental factors. The relationship between Rs and soil temperature moisture in these four forests was analyzed over three years. The results showed that the mean Rs in Larch, Korean pine, and Chinese pine plantations and broadleaved mixed forest were (2.31±0.01), (2.07±0.71), (1.55±0.03) and (2.24±0.02) μmol·m-2·s-1, respectively. Moreover, Rs and soil temperature at a depth of 10 cm (t10) were significantly correlated (P=0.000<0.01). The Q10values in the four forests during the growing seasons were 3.32, 4.46, 4.12 and 3.59, respectively. The Rs of Korean pine plantation was more sensitive to t10than the other three forests. Rs was also continuously monitored in a larch plantation during the non-growing season (from November to the following March), which revealed that the Rs in the non-growing season was divided into three stages: the frozen period (t10-mean: 10.0~2.0 ℃), freeze-thaw period (t10-mean: 0.5~2.0 ℃) and thawing period (t10-mean:>2.0 ℃). A weak Rs was observed in larch plantations during the non-growing season (0.01~1.8 μmol·m-2·s-1), and the Rs and t10values were significantly correlated throughout this period (r2=0.586 3, P=0.000<0.01). Moreover, Rs and t10were not significantly correlated in the frozen period (P=0.503>0.05), Rs changes were more intense during the freeze-thaw period, and were significantly regulated variations when compared with the frozen period. Rs showed a greater increase during the freeze-thaw period than the frozen period. The Q10value of the larch plantation during the non-growing season was 4.65, which was 1.4 times that of the growing season. Overall, these findings suggest that Rs was more sensitive to global climate change during the non-growing season than the growing season for temperate forests.
soil respiration; temperature sensitivity; forest; Northeast China
10.16258/j.cnki.1674-5906.2015.11.001
S718.55;X171.1
A
1674-5906(2015)11-1757-08
林业公益性行业科研专项(201204101;201404303);国家自然科学基金项目(31370616);辽宁省农业领域青年科技创新人才培养计划(2014015)
张慧东(1981年生),男,高级工程师,博士研究生,主要从事森林生态系统定位观测研究。E-mail: zhanghuidong00@163.com *通信作者:周梅(1960年生),女,教授,博导,主要从事森林生态学研究。Email: dxalzhoumei@yahoo.com.cn
2015-08-07
引用格式:张慧东, 尤文忠, 魏文俊, 周梅. 暖温带-中温带过渡区 4种典型森林土壤呼吸的温度敏感性[J]. 生态环境学报, 2015, 24(11): 1757-1764.
猜你喜欢
南方农业(2021年5期)2021-12-27
今日农业(2021年10期)2021-11-27
山东林业科技(2019年2期)2019-06-03
现代营销(创富信息版)(2018年2期)2018-08-15
中国粮油学报(2018年12期)2018-03-19
中国公路(2017年10期)2017-07-21
辽宁林业科技(2017年4期)2017-06-22
中国计划生育学杂志(2017年3期)2017-06-01
浙江大学学报(工学版)(2016年2期)2016-06-05
中国房地产业(2016年24期)2016-02-16
