
柏木低效林林窗改造初期边界木细根形态和生物量变异
2016-01-12 08:37苏宇,李贤伟,刘运科等
西北植物学报 2015年3期
*通信作者:李贤伟,教授,主要研究从事森林培育研究。E-mail:lxw@sicau.edu.cn
柏木低效林林窗改造初期边界木细根形态和生物量变异
苏宇1,李贤伟1,2*,刘运科1,李金凤1,尹海峰1,王之南1
(1 四川农业大学 林学院,四川雅安 625014;2 长江上游林业生态工程四川省重点实验室,四川雅安 625014)
摘要:为深入探究川中丘陵区柏木低效林林窗改造初期边界木林细根的生态适应,利用土柱法对34年生柏木低效林在人造小面积开窗(50 m2、100 m2、150 m2)处理1年后的林窗边界木1~5级细根直径、根长、比根长、根长密度和生物量变异进行研究。结果显示:(1)柏木1~5级细根平均直径、根长随着根序的上升而增加,比根长、根长密度随着根序的上升而减小。 (2)与林内对照(无林窗)相比,人工开林窗处理后边界木细根在同一根序上的平均直径变化不显著,根长和比根长增加,根长密度和生物量则呈降低的趋势。(3)开窗 150 m2面积林窗抚育后的柏木边界木更大限度地提高了低级根序的物质分配,提高比根长并降低组织密度,更快速地利用地下林窗空间资源。研究表明,柏木低效林林窗式疏伐得到的土壤资源由于更新草本、灌木的生长,增加了林木根系竞争土壤空间的压力,使得根系进行扩展性生长,细根形态和生物量表现出异质性变化。
关键词:低效林;林窗;边界木;根序;形态;生物量
收稿日期:2014-10-07;修改稿收到日期:2015-01-09
基金项目:国家“十二五”科技支撑计划项目(2011BAC09B05);四川省科技支撑计划项目(2010NZ0049);国家自然科学基金面上项目(30771717)
作者简介:苏宇(1991-),女,在读硕士研究生,主要从事森林生态研究。E-mail:yusu110@163.com
中图分类号:Q948.12 文献标志码:A
Fine Root Morphology and Biomass Characteristics at
Preliminary Stage of Gap Border Trees of Reformed
Low BeneficialCupressusfunebrisForests
SU Yu1,LI Xianwei1,2*,LIU Yunke1,LI Jinfeng1,YIN Haifeng1,WANG Zhinan1
(1 College of Forestry,Sichuan Agricultural University,Ya’an,Sichuan 625014,China;2 Sichuan Province Key Laboratory of Forestry Ecological Engineering in Upper Reaches of Yangtze River,Ya’an,Sichuan 625014,China)
Abstract:In order to adapt the ecological adaptation of fine root of gap border trees at preliminary stage,we studied the 34-year old Cupressus funebris plantation in central Sichuan hilly region,researched the first five orders of fine rootdynamic rules of fine root morphology and biomass ofdifferent size of gaps (50 m2,100 m2 and 150 m2) and C.funebris plantation (Ⅳ) at the first year.The main results were found as follows:(1)The fine root morphology and biomass ofdifferent root orders of C.funebris plantation have obvious change rule.The specific root length(SRL) and root lengthdensity(RLD) of the first order root were the largest,but the averagediameter,average root length and biomass of the first order root were the smallest.The SRL and RLD of the fifth order root were the smallest,but the averagediameter and biomass of the fifth order root were the largest.Fine root biomass and averagediameter of C.funebris significantly increased with increase in order(P<0.05),while SRL and RLDdecreased.(2)Compared with C.funebris plantation (Ⅳ),fine root of gap border trees(GBT) have insignificant averagediameter,increase average root length and SRL,reduce RLD and biomass at the same root order.(3)GBT at the gap of 150 m2 highly raised Matterdistribution of low root orders,with increasing SRL anddecline RLD,to use underground forest gap in a more quality way.In conclusion,with the renew of herbs and shrubs the forest gaps have increased pressure of root to competition soil spaces,expansionary growth and heterogeneous exchange of fine root morphology and biomass.
Key words:low beneficial forests;forest gap;gap border trees;root order;morphology;biomass
森林是陆地生态系统的主体,维持着整个生态系统的物质循环和能量流动[1]。细根作为森林生态系统中的重要组成部分,是碳库平衡和土壤养分循环进入陆地生态系统的基础[2-3],细根在执行土壤资源有效分配的功能时,对其变化能做出快速的反应,并在生物量分布与形态特征上都表现出异质性[4-5],虽然细根(D<2 mm)生物量仅占根系总生物量的3%~30%,但在总碳循环和年初级生产力(NPP)上都占有较大的比重[6-7]。细根形态在根系功能的维持中作用重大,是衡量植物生产力的重要因素[8-10]。细根比根长(specific root length,单位重量的根长)和根长密度(root lengthdensity,单位面积的根长)决定根系吸收养分和水分的能力[11],但根系构型的改变可能并不引起根系总生物量的变化[12]。同样,单一的生物量指标不能作为根系系统中吸收器官潜在功能的指示指标,采用综合的根系形态参数可反映林地养分潜能并有效预测林分生产力[13]。
中国作为世界上人工林面积最大的国家,人工林地力衰退问题一直阻碍着林业可持续发展,地力衰退是一个复杂的生态学过程,而本质上就是林地林木生长养分吸收与土壤养分供给之间的不平衡。在人工林的抚育过程中,林窗干扰拓展了林隙空间、释放了有效资源,使得生物多样性增加、林下次生生长恢复,已有的研究多集中于林窗中光照、微地形、小气候、生物多样性和资源利用率等方面[14-15],而针对林窗边缘边界木的研究则缺少相应的关注[16-17],在小面积林窗条件下,大部分的林窗面积被边界效应影响,边界木对林窗环境因子的响应极为显著[18],林冠林窗出现后伴随着上层优势苗木的死亡和倒伏,根系的生物量也随之减少,形成不同于林内地下生物量的地下林窗微环境。地下林窗也同林冠林窗一样是森林资源有效改善的重要部分,现有很少从林下土壤及根系生长的角度进行研究。
本研究以川中低山丘陵区柏木低效林为研究对象,采用小面积林窗采伐的人为干扰方式进行抚育管理,在控制水土流失、缩短植被恢复期的情况下,有效改善林分结构、促进林木生长、维持系统的生物多样性。在人工开窗一年后针对不同面积林窗边界木的地下根系进行研究,探讨地下空窗对于边界木细根生长与生物量的影响,充分认识林窗的基本特征,了解林窗边界木地下竞争机制,加强对林木地上以及地下生长趋向性发育的探索,将有助于进一步揭示林窗对森林更新群落组成、结构和功能的动态影响及其维持机制。
1研究地区与研究方法
1.1 研究区概况
研究区位于德阳市旌阳区和新镇(104°15′~104°33′E,31°5′~31°20′N),地处川中丘陵区北部边缘,属于典型的低山丘陵地貌。该区属四川盆地中亚热带湿润气候区,气候特征四季分明,气候温和,雨量充沛,日照较少,年平均气温16~17 ℃,年总降水量880~940 mm,年平均无霜期270~290d。该区域森林土壤主要为石灰型紫色土,土层浅薄,通常不超过40 cm。样地区域土壤基本养分中全碳为39.11 g/kg,全氮为1.71 g/kg,全磷为0.27 g/kg,水解氮76.34 mg/kg,有效磷7.58 mg/kg,速效钾105.34 mg/kg。
1.2 样地设置与研究方法
该研究区域柏木人工林为34年生林分,区内林木尚处于中幼龄林阶段,其平均树高6.5 m,平均胸径8.0 cm,郁闭度大于0.8,单位蓄积量约为44.7 m3/hm2,远低于全国平均水平83.63 m3/hm2[19],属于典型的低效林。针对实验区林分的生境条件与生长情况,2012年3月,选择在山体南坡向,中坡位,坡度20°~30°,生长状况及经营管理水平相似的地段进行小面积林窗干扰采伐,面积分别为50 m2、100 m2和150 m2,每种采伐面积3个重复,各处理样地的基本情况见表1。人造林窗均为南北向长、东西向短的近似椭圆形,且各林窗之间间隔15 m以上的过渡带。清除枯枝和杂灌,建立铁丝围栏,防治人畜干扰,使其自然恢复。林窗面积的计算采用椭圆面积公式:A=πLW/4,式中,L为林窗中最长的直径,W为与L垂直的最大直径,其相交点作为林窗的中心点[20]。
2013年5月,在每个林窗周边选取8株柏木边界木,同时在未进行林窗改造的对照林分随机选取8株样木作为研究对象。在面向林窗方向距林窗边界木树基50 cm内用土块法采集细根样品,土块大小为15 cm×15 cm×15 cm。土块放入低温保鲜盒中带回实验室。在实验室,把未分离土块放入盛有低温(2~4 ℃)去离子水中,小心的将根系取出。在10倍放大镜下,将根系上剩余的土、有机物质和已死的细根片段洗净。土块得到的细根样使用用Fitter的根系序列位置命名方法对细根进行分级[21],位于根轴最远端没有分枝的根称为1级根,1级根着生在2级根上,依次分到5级根,着生于高级根上没有分枝的根也算作1级根。
应用Epson数字化扫描仪(Expression 10000XL 1.0)和Win RHIZO 2012根系图像分析系统软件对不同处理的1~5级柏木细根形态特征指标(直径、根长、表面积等)进行定量分析。然后在65 ℃烘干至恒重,用电子天平称重(精确到0.000 1 g)测定各级细根质量。得到各处理细根比根长(SRL),根长密度(RLD)以及生物量数据,计算公式为:
SRL=L/M;RLD=L/V;B=M/d2×104
式中,SRL为比根长(m·g-1);L为根系长度(m);M为根系干质量(g);RLD为根长密度(m·m-3);V为土块体积(m3);B为细根生物量(g·m-2);d为土块边长(cm)。
1.3 数据处理与统计分析
数据采用Excel 2010和SPSS 20.0软件做统计处理和分析,采用单因素方差分析(one-way ANOVA)结合Duncan检验分析比较不同开窗面积处理下边界木不同根序间细根直径、根长、比根长、根长密度的差异(P<0.5)。
2结果与分析
2.1 不同林窗面积下边界木各级根序形态特征
由图1,A可以看出,各处理样地柏木细根平均
直径随着根序的上升而增加,前5级细根的直径在0.36~1.50 mm之间,其中1、2级根之间差异不显著(P>0.05),2~5级根之间则呈现显著性差异(P<0.05),且各面积开窗边界木1~5级细根平均直径大小均无显著影响;此外,各处理样地柏木1~5级细根平均直径呈现不均匀增长状态,并在3级根出现快速增长的拐点,且3~5级根的平均直径分别是1级根的1.3倍、2.1倍和3.6倍。
各处理柏木细根根长方差分析显示(图1,B),不同面积开窗处理对边界木各级根序根长的影响不同。50、100 m2开窗处理后,林窗边界木细根平均根长在1~5级根之间均表现出显著的差异(P<0.05);150 m2开窗后,林窗边界木1~5级细根平均根长则同对照处理的柏木细根变化规律一致,其中,在1、2级,2、3级根序之间均未表现出显著差异性,在3~5级根序间差异显著(P<0.05);50、100 m2开窗处理后,边界木2~4级根序细根平均根长大于150 m2林窗细根;此外,对照组的5级根根长显著小于各面积林窗处理(P<0.05)。
不同开窗面积对柏木细根不同根序比根长的影响见图1,C。各处理细根SRL均表现为1~5级根依次降低,然而,同一级根序内SRL随着开窗处理面积的增加而增加,对照处理则最低;各处理的同级根序SRL规律表现为:对照<50 m2<100 m2<150 m2;此外,150 m2开窗处理后细根SRL在1~5级根均较对照处理有显著增加(P<0.05),100 m2处理细根SRL仅在3~5级根显著大于对照(P<0.05),而50 m2处理细根SRL与对照比较并没有显著差异(P>0.05)。
由图1,D可发现,各面积开窗处理后,边界木1~3级细根RLD均显著低于对照组(P<0.05),而4、5级根RLD仅在150 m2开窗处理后,其边界木较对照组出现显著降低(P<0.05)趋势。总之,不同面积开窗处理对细根不同根序根长密度影响不一致,1、2级根RLD表现为:50 m2<100 m2<150 m2,而3~5级根RLD表现为:50 m2、100 m2处理均大于150 m2处理。
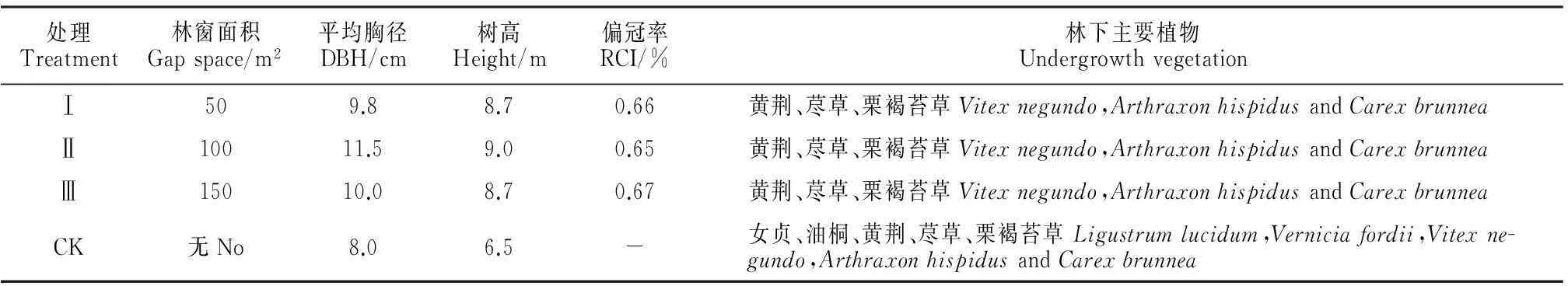
表1 柏木人工林人造林窗处理样地基本概况
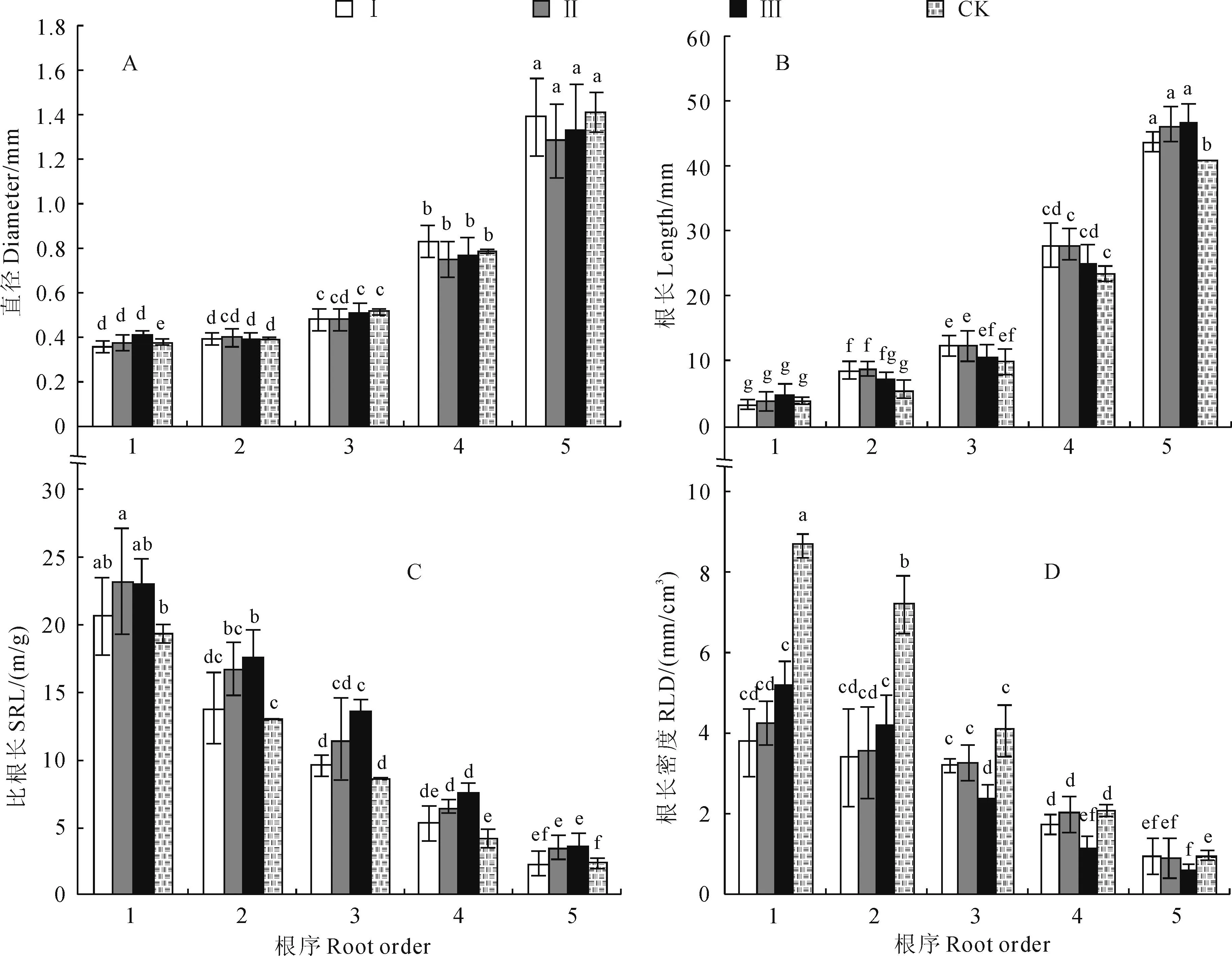
图1 不同林窗面积下边界木各级根序细根直径、根长、比根长、根长密度
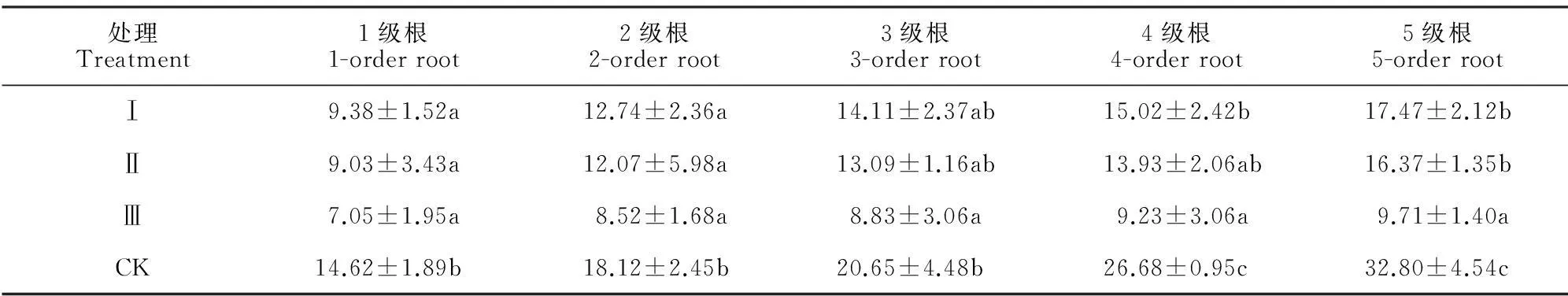
处理Treatment1级根1-orderroot2级根2-orderroot3级根3-orderroot4级根4-orderroot5级根5-orderrootⅠ9.38±1.52a12.74±2.36a14.11±2.37ab15.02±2.42b17.47±2.12bⅡ9.03±3.43a12.07±5.98a13.09±1.16ab13.93±2.06ab16.37±1.35bⅢ7.05±1.95a8.52±1.68a8.83±3.06a9.23±3.06a9.71±1.40aCK14.62±1.89b18.12±2.45b20.65±4.48b26.68±0.95c32.80±4.54c
注:不同小写字母表示同列不同处理柏木细根生物量差异显著(P<0.05)。
Note:Different normal letters within the same column meant significantdifference at 0.05 level.
2.2 不同林窗面积下边界木各级根序生物量
不同林窗面积下边界木1~5级根序生物量以及分配比例(表2;图2)。各处理柏木细根生物量分配均表现为1级根最小,5级根最大;150 m2开窗处理边界木各级根序生物量均显著小于对照,50 m2、100 m2处理除3级根外生物量也显著小于对照(P<0.05);此外,随着开窗面积的增加,50 m2、100 m2、150 m2开窗处理的1~3级根生物量占前5级根总生物量的比值均有所增加,1级所占比例分别比对照组增加0.70 %、1.05 %、3.31%。
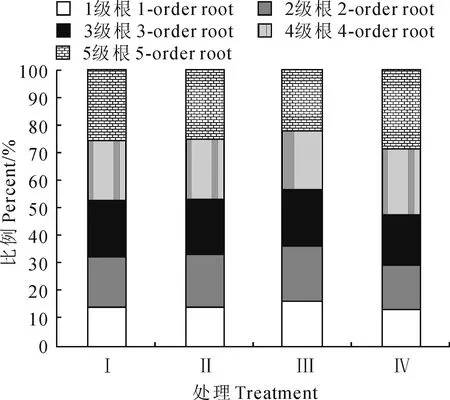
图2 各级根序占总生物量的百分比
3结论与讨论
3.1 不同林窗面积下边界木细根形态特征
根系系统是植物从土壤中吸收水分与养分的主要器官,在地下生态系统的循环中有重要意义[22],细根直径、根长、比根长和根长密度都是反映细根形态和功能的重要指标。本文以人工抚育开窗处理初期反应最快的林窗边界木细根为研究对象,研究柏木细根特征和各形态指标对不同面积开窗处理的响应。本实验对柏木1-5级细根形态的研究表明,1级根直径、根长、比根长最小,根长密度最大,而5级根直径、根长、比根长最大,根长密度最小,这种根序间的变化趋势与差异性说明了细根存在普遍异质性[23-25],细根形态异质性与其生理功能相适应,这些特征同时又受到植物自身遗传特性及立地条件的影响,并对土壤条件及环境变化的反应较为敏感[26-27],故而细根中行使不同生理功能的根序会做出不同的应对策略。
开窗后,边界木细根在同一根序上的平均直径变化不显著,而根长和比根长则有所增加,根长密度呈现降低的趋势。不同面积抚育开窗后的柏木边界木以及对照样地内柏木的细根直径均呈现随着根序增加直径增加的规律,且各处理之间并没有显著的变化,这与“扩展的土壤资源并不能通过显著增加柏木细根的直径来拓展养分利用范围”的结论相吻合[28]。本研究中柏木前4级根序的直径都在1 mm以下,直径较细,具有较强的吸收功能,有利于从土壤中吸收养分和水分[29],同时,开窗处理并没有对周转快速的1级细根根长产生影响,但随着根序的增加细根对开窗响应的敏感度呈现上升趋势,相较于150 m2林窗中较为疏松的边界木分布情况,位于50 m2和100 m2林窗的边界木根长出现了更显著的增加,这使得根系增殖速度快,活力提高,土壤空间的利用效率增加[30-31]。
林窗边界木各级根序细根比根长较对照都有增幅,这种变化随开窗面积的增加而增加,细根中出现较大的比根长表明根系在该根序生物量的投入方面提高了效率;根长密度作为单位面积上根的长度,直观反映树木根系吸收水肥的能力,是树木地下竞争力的体现[32],但是开窗处理减小了边界木细根根长密度,1~3级根和4、5级根序表现出不同的响应机制,低级根序的细根根长密度降低程度尤为突出。此现象可能是由于开窗处理带来的林冠林窗与地下林窗的双向形成和光照增加导致的土壤水分养分变化和林下生物多样性的改变,细根根长密度与土壤有效N呈现显著相关[33],在开窗初期地上部分的伐除导致边界木地下有效养分竞争的增加,边界木细根生长的相对应策略变为增加单位生物量的根系长度,进而通过比根长的增加和根长密度的减少来快速利用土壤空间。
3.2 不同林窗面积下边界木细根生物量
细根是林木从土壤中吸收养分和水分并维持自身正常生理活性的重要器官,其生物量与立地条件、土壤类型、群落结构、气候、树种等因素有关[34]。川中丘陵区森林分布范围内土壤类型主要为新冲积物的灰棕和紫色冲积土,土层浅薄,低于40 cm,根据实地土壤剖面确定柏木根系中的细根分布在15 cm以内的浅表层土壤中(占总数生物量的85%以上),Makita等[35]的研究同样指出表层土壤中根系的65%为细根。本研究对柏木细根生物量的研究就集中在15 cm以内,结果表明柏木细根生物量随根序上升增加,然而前人的研究表明不同树种的细根生物量随着根序变化的规律并不一致,这说明细根生物量在各根序的分布会由于树种和立地条件的不同而不同[24,36-37]。
开窗处理伴随着边界木细根生物量的减少,在50 m2开窗的边界木细根上缩减39.12%,开窗150 m2后更是缩减61.60%,但由于在开窗的初期,抚育采伐强度的增大,使得单位面积上各级根序的生物量随着开窗面积的增加而减少,且早期树木生长的扩展性更多的表现在细根不同根序生物量的分配上。本研究发现,开窗后柏木低级根序所占总生物量的比值有不同程度的提高,特别是在150 m2开窗处理下,不同根序细根生物量所占比值的变化反应出根系功能分配的变化,造成此结果的原因可能是植物细根可塑性对潜在的养分、水分吸收和物种本身指定的觅食策略的响应使得低级根序在生物量总量中所占的比值增加,优先扩展活性强的器官,同时伴随降低根系组织密度拓展利用地下林窗空间,高效平衡其生长所需的资源[38]。同时,低级根的高比根长可以保持细根的吸收能力,且由于低级根生物量较小,细根构建和维持单位长度细根消耗的碳水化合物较少,由这种比根长与生物量不对称的比例关系可知,异质性的物质分配对细根的吸收功能有利[39]。
本研究使用的小面积林窗抚育间伐方式,快速干扰了已处于退化状态的柏木纯林,增加了林内光照,改变了土壤条件,影响了植物地上部分的生长,而这些因素均与细根的生长有密切的联系,林下植被物种数和多样性随开窗面积的增加而增加,更新的林下灌木和草本都是浅根系植物,这势必增加了乔木树种根系竞争土壤资源的压力[40],使得乔木树种的根系需要更加快速的进行扩展性、养分诱导性生长[41]。在开窗抚育的初期,林窗边界木作为首先受到林窗影响的生态边界,其根系在形态和生物量分布上都做出了快速的竞争策略,在直径基本不变的基础上,通过增加根长,降低组织密度,迅速占领营养空间。研究根系分布在环境改变响应上的敏感性,有利于完整掌握在人为干扰下的森林生态系统养分库养分循环和碳分配流动。林窗改造恢复过程是一个动态的过程,所形成的界面是非稳定的界面,针对整体林分变化的持续监测是人工林分抚育的必要手段,更是今后研究的方向。
参考文献:
[1]FINER L,OHASHI M,NOGUCHI K,etal.Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics[J].ForestEcologyandManagement,2011,262(11):2 008-2 023.
[2]FAHEY T J,HUGHES J W.Fine rootdynamics in a northern hardwood forest ecosystem,Hubbard Brook Experimental Forest,NH[J].JournalofEcology,1994:533-548.
[3]KING J S,ALBAUGH T J,Allen H L,etal.Below-ground carbon input to soil is controlled by nutrient availability and fine rootdynamics in loblolly pine[J].NewPhytologist,2002,154(2):389-398.
[4]HISHI T.Heterogeneity of individual roots within the fine root architecture:Causal links between physiological and ecosystem functions[J].JournalofForestResearch,2007,12(2):126-133.
[5]MAJDI H,TRUUS L,JOHANSSON U,etal.Effects of slash retention and wood ash addition on fine root biomass and production and fungal mycelium in a Norway spruce stand in SW Sweden[J].ForestEcologyandManagement,2008,255(7):2 109-2 117.
[6]HELMISAARI HS,MAKKONEN K,KELLOMAKI S,etal.Below- and above-ground biomass,production and nitrogen use in Scots pine stands in eastern Finland[J].ForestEcologyandManagement,2002,165(1):317-326.
[7]TATENO R,HISHI T,TAKEDA H.Above- and belowground biomass and net primary production in a cool-temperatedeciduous forest in relation to topographical changes in soil nitrogen[J].ForestEcologyandManagement,2004,193:297-306.
[8]HARPER J L,JONES M,HAMILTON N R.The evolution of roots and the problems of analyzing their behavior[M].Oxford:Blackwell Scientific Publications,1991:3-24.
[9]HENDRICK RL,PREGITZER KS.Thedemography of fine roots in a northern hardwood forest[J].Ecology,1992,73:1 094-1 104.
[10]NADELHOFFER KJ,RAICH JW.Fine root production estimates and belowground carbon allocation in forest ecosystems[J].Ecology,1992,73:1 139-1 147.
[11]FRANSEN B,KROON HD,BERENDSE F.Root morphological plasticity and nutrient acquisition of perennial grass species from habitats ofdifferent nutrient availability[J].Oecologia,1998,115(3):351-358.
[12]HODGE A.The plastic plant:root responses to heterogeneous supplies of nutrients[J].TheNewPhytologist,2004,162(1):9-24.
[13]CRAINE J M.Competition for nutrients and optimal root allocation[J].PlantandSoil,2006,285(1-2),171-185.
[14]MCGUIRE J P,MITCHELL R J,MOSER E B,etal.Gaps in a gappy forest:plant resources,longleaf pine regeneration,and understory response to tree removal in longleaf pine savannas[J].CanadianJournalofForestResearch,2001,31(5):765-778.
[15]DECHANTAL M,LEINONEN K,KUULUVAINEN T,etal.Early response ofPinussylvestrisandPiceaabiesseedlings to a experimental canopy gap in a boreal spruce forest[J].ForestEcologyandManagement,2003,176(1):321-336.
[16]XIAN J R(鲜骏仁),HU T X(胡庭兴),ZHANG Y B(张远彬),etal.Effects of forest canopy gap onAbiesfaxonianaseedling's biomass and its allocation in subalpine coniferous forest of West Sichuan[J].ChineseJournalofAppliedEcology(应用生态学报),2007,18(4):721-727(in Chinese).
[17]WU Q G(吴庆贵),WU F ZH(吴福忠),YANG W Q(杨万勤),etal.Characteristics of gaps anddisturbance regimes of the alpine fir forest in Western Sichuan[J].J.Appl.EnvironBiol.,2013,19(06):922-928.
[18]HUGHES M K,FUNKHOUSER G.Frequency-dependent Climate Signal in Upper and Lower Forest Border Tree Rings in the Mountains of the Great Basin[M]//Climate Variability and Change in High Elevation Regions:Past,Present & Future.Springer Netherlands,2003:233-244.
[19]XU P F(徐培富),CHEN J J(陈进军).The effects ofdifferent cultural practice on speciesdiversity of low-beneficial cypress forests in low hill of Mianyany City[J].JournalofSichuanForestryScienceandTechnology(四川林业科技),2009,30(5):88-94(in Chinese).
[20]XIAN J R(鲜骏仁),HU T X(胡庭兴),WANG K Y(王开运),etal.Studies on the characteristics of gap border trees in the subalpine coniferous forest in Western Sichuan [J].ForestResearch(林业科学研究),2004,5:636-640(in Chinese).
[21]FITTER A H,STICKLAND T R.Architectural analysis of plant root systems.Ⅲ.Studies on plans under field conditions[J].NewPhytologist,1992,121(2):243-248.
[22]HODGE A.The plastic plant:root responses to heterogeneous supplies of nutrients[J].NewPhytologist,2004,162(1):9-24.
[23]PREGITZIER K S,DEFOREST J L,BURTON A J,etal.Fine root architecture of nine north American trees[J].EcologicalMonographs,2002,72(2):293-309.
[24]SHI W(师伟),WANG ZH Q(王政权),LIU J Q(刘金梁),etal.Fine root morphology of twenty hardwood species in Maoer shan natural secondary forest in northeastern China[J].JournalofPlantEcology(植物生态学报),2008,32(6):1 217-1 226(in Chinese).
[25]LIU J(刘佳),XIANG W H(项文化),XU X(徐晓),etal.Analysis of architecture and functions of fine roots of five subtropical tree species in Huitong,Hunan Province,China[J].JournalofPlantEcology(植物生态学报),2010,34(8):938-945(in Chinese).
[26]HENDRICK R L,PREGITZER K S.Thedynamics of fine root length,biomass,and nitrogen content in two northern hardwood ecosystems[J].CanadianJournalofForestResearch,1993,23(12):2 507-2 520.
[27]YU S Q,WANG Z Q,SHI J W,QUAN X K,etal.Estimating fine-root longevity of and using mini-rhizotrons[J].JournalofPlantEcology,2007,31(1):102-109.
[28]NOGUCHI K,HAN Q,ARAKI M G,etal.Fine-rootdynamics in a young hinoki cypress (Chamaecyparisobtusa) stand for 3 years following thinning[J].JournalofForestResearch,2011,16(4):284-291.
[29]LIU J L(刘金梁),MEI L(梅莉),GU C C(谷加存),etal.Effects of nitrogen fertilization on fine root biomass and morphology ofFraxinusmandshuricaandLarixgmelinii:A study with in-growth core approach[J].ChineseJournalofEcology(生态学杂志),2009,28(1):1-6(in Chinese).
[30]PERSSON H,MAJDI H,CLEMENSSON-LINDELL A.Effect of aciddeposition on tree roots[J].EcolBull,1995,44:158-167.
[31]PERSSON H,AHLSTROM K.Fine root response to nitrogen supply in nitrogen manipulated Norway spruce catchment areas[J].ForestEcologyandManagement,2002,168(1),29-41.
[32]BAYALA J,TEKLEHAIMANNOT Z,OUEDRAOGO S J.Fine rootdistribution of pruned trees and associated crops in a parkland system in Burkina Faso[J].AgroforestrySystems,2004,60(1):13-26.
[33]EISSENSTATd M,YANAI Rd.The ecology of root lifespan[J].AdvancesinEcologicalResearch,1997,27:1-60.
[34]CANHAM C A,FROEND R H,STOCK Wd,etal.Dynamics of phreatophyte root growth relative to a seasonally fluctuating water table in a Mediterranean-type environment[J].Oecologia,2012,170(4):909-916.
[35]MAKITA N,ISHII H,KANAZAWA Y,etal.Very fine roots respond to soildepth biomass allocation,morphology,and physiology in a broad leaved temperate forest[J].EcologicalResearch,2011,(1),26:95-104.
[36]WEI P(魏鹏),LI X W(李贤伟),FAN CH(范川),etal.Fine root biomass and carbon storage in surface soil ofCinnamomumcamphoraplantation in Rainy Area of West China[J].ChineseJournalofAppliedEcology(应用生态学报),2013,24(10):2 755-2 762(in Chinese).
[37]LI R X(李瑞霞),PENG T T(彭婷婷),LIU N(刘娜),etal.Response of carbon storage,fine root biomass and morphology to thinning inPlatycladusorientalisplantation[J].JournalofNortheastForestryUniversity(东北林业大学学报),2013,41(5):33-37(in Chinese).
[38]GUOd L,XIA M X,WEI X,etal.Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J].NewPhytol.,2008,180(3):673-683.
[39]GUOd L,MITCHELL R J,HENDRICKS J J.Fine root branch orders responddifferentially to carbon source-sink manipulations in a longleaf pine forest[J].Oecologia,2004,140(3):450-457.
[40]WANG ZH Q(王政权),ZHANG Yd(张彦东).Study on the root intreactions betweenFraxinusmandshuricandLarixgmelinii[J].ActaPhytoecologicaSinica,2000,24(3):346-350(in Chinese).
[41]WANG Z H(王祖华),LI R X(李瑞霞),HAO J P(郝俊鹏),etal.Effects of thinning on fine root morphology in Chinese Fir plantations[J].JournalofNortheastForestryUniversity(东北林业大学学报),2011,39(6):13-15(in Chinese).
(编辑:潘新社)
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
中国绿色画报(2016年7期)2016-12-26
文艺生活·下旬刊(2016年11期)2016-12-12
商情(2016年40期)2016-11-28
艺术科技(2016年9期)2016-11-18
人间(2016年27期)2016-11-11
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
