
羊踯躅和日本杜鹃的传粉生物学与杂交育种初探
2016-02-06 09:27杨灿娇郑硕理杨荣萍张敬丽
中国野生植物资源 2016年6期
杨灿娇,郑硕理,杨荣萍,田 伟,张敬丽*
(1.云南农业大学 园林园艺学院,云南 昆明 650201;2.鲁东大学 农学院,山东 烟台 264025)
羊踯躅和日本杜鹃的传粉生物学与杂交育种初探
杨灿娇1,郑硕理1,杨荣萍1,田 伟2,张敬丽1*
(1.云南农业大学 园林园艺学院,云南 昆明 650201;2.鲁东大学 农学院,山东 烟台 264025)
杜鹃属植物在自然界的杂交现象是普遍存在的,国内外已经通过杂交育种培育出许多杜鹃花新品种。以羊踯躅和日本杜鹃为试材,进行传粉生物学特性研究和种间杂交及自交试验,为杜鹃花属植物的杂交育种及新品种选育提供基础。
杜鹃花;传粉生物学;繁育系统;亲和性
羊踯躅[Rhododendronmolle(Bl.) G. Don]和日本杜鹃(Rhododendronmollesubsp.japonicum)同属于杜鹃科(Ericaceae)杜鹃属(RhododendronL.)羊踯躅亚属[Subgen.Pentanthera(G. Don) Pojarkova]。据《中国植物志》记载[1],该亚属分三组,主要分布于北美洲东部,欧洲和亚洲(自我国西南、华南、华中、华东)到日本,朝鲜有少数种类。全世界约24种,我国只有1组。羊踯躅和日本杜鹃均为落叶灌木,羊踯躅产自中国,叶纸质,新叶上面被微柔毛,下面密被灰白色柔毛,总状伞形花序顶生,花多达13朵,花冠阔漏斗形,黄色或金黄色,内有红色斑点(图1:A)。日本杜鹃叶纸质,新叶上面被微柔毛,下面密被灰白色柔毛,总状伞形花序顶生,花多达9朵,花冠阔漏斗形,橙色,内有黄色斑点(图1:B),为张长芹研究员引自日本。
国外杜鹃花杂交育种工作起步较早,通过引种我国的杜鹃花,经过近200年的研究,已经培育出了许多耐寒、大花、早花、晚花和花香的杜鹃花新品种,截至2004年,已登记的杜鹃花新品种达2.8万个[7],但国内杜鹃花新品种的培育起步晚,基础薄弱,仅见张长芹[2-5]、刘小青[6]等的报道。对杜鹃花的传粉生物学和繁育系统的研究,国内仅见张敬丽对迷人杜鹃等四种杜鹃花的研究[8],边才苗对云锦杜鹃的研究[9],田晓玲对大喇叭杜鹃的研究[10-12]和白天对锈叶杜鹃的研究[11]。
羊踯躅和日本杜鹃,前者产自中国,后者引自日本,长期的生殖隔离和地理隔离使得二者的在遗传基因方面存在着一定的差异。而杂交双亲的遗传差异越大,杂交后代出现变异的几率就越高,出现超亲优良性状的杂种的可能性也就越大[13]。本研究拟通过杜鹃传粉生物学和人工授粉研究,初步探讨羊踯躅和日本杜鹃的开花习性和杂交育种情况,可以为充分利用杜鹃花资源,培育具有自主产权的新品种作出初步研究,同时,能够为该属其他种的杂交育种和种质资源保存方面研究提供借鉴和理论依据。
1 研究方法
1.1 开花物候的观察
2014年3月至5月,对羊踯躅和日本杜鹃开花情况及单花的开花特性进行观察、记录。标记特定花序开花的时间,在自然条件下连续数天定位观察并记录花朵的开放过程,直至花瓣脱落。
1.2 传粉昆虫的观察
在两种杜鹃花开花期间,每种杜鹃花各选取5株已开放的植株进行标记,从7:00—18:00,连续观察1周,对每种昆虫的访花行为进行摄影、描述。记录在每花序的访花昆虫的种类。并观察不同天气情况下昆虫的传粉行为。
1.3 花粉活力与柱头可授性测定
花粉活力用TTC(2,3,5-triphenyl tetrazolium chloride)法测定花粉的活力和寿命。具体做法是:标记开花时间不同的花朵,将散出时间不同的花粉撒在载玻片上,滴加含0.5%TTC的蔗糖溶液,迅速盖上盖片,置入内有湿润滤纸的培养皿中,连同培养皿放置在37 ℃黑暗条件下2 h。统计盖片中央部分3~5个视野中全部观察花粉中红色花粉所占比例。
柱头可授性用联苯胺-过氧化氢法测定。具体方法为:在盛花期,标记开花时间不同的花朵,每天中午采开花后不同天数的花朵,将其柱头浸入凹面载片中含有联苯胺-过氧化氢反应液[V(1%联苯胺)∶V(3%过氧化氢)∶V(水)=4∶11∶22]的凹陷处。若柱头具可授性,则柱头周围的反应液呈现蓝色并有大量气泡出现。
1.4 繁育系统研究
为了检测这两种杜鹃花的繁育系统,我们在研究地点设立了如下7个处理并统计结实率(各处理5~10个花序,每花序5~10朵花):(1)自然对照,不作任何处理,挂牌直至果实成熟;(2)不去雄,直接套袋,检验是否自花授粉;(3)去雄不套袋,判断是否虫媒异交,及酬物;(4)人工自花授粉后,去雄,套袋,鉴定是否自交亲和;(5)同种不同株人工异花授粉;(6)异种人工异花授粉;(7)去雄,套网,鉴定是否风媒传粉。
1.5 种子萌发实验
我们对人工授粉实验所得到的种子在人工气候培养箱中按照同一条件(每天光照时间为12 h,光照强度为2 000 lx, 温度为23 ℃),进行种子萌发实验,在播种第40天时统计其萌发率及萌发势,3次重复,结果取3次萌发的平均值。
2 结果与分析
2.1 开花物候的观察
羊踯躅开花较早,花期大约从2月下旬至4月底。日本杜鹃的开花时间稍晚, 花期从3月初至5月中旬。
羊踯躅和日本杜鹃的单花开花时间都在1周左右。在花冠展开的前1天柱头已具有极少量粘液; 在开花后2~4 天, 柱头粘液最多。花粉则成丝状散出, 具粘性; 在开花5天后, 柱头粘液渐少, 花粉已基本完全散出; 开花1周左右, 柱头开始枯萎,花冠脱落, 雄蕊也开始脱落, 完成单花开花周期。
经观察发现,羊踯躅和日本杜鹃的花期存在重叠的现象,这就给两种杜鹃花的杂交育种提供了条件,可以直接采收花药,进行杂交授粉。
2.2 传粉昆虫的观察
羊踯躅和日本杜鹃的访花昆虫都在单花开花后2~3 天内较多。访花昆虫活动高峰期, 随太阳照射的强度而改变。天气晴朗时访花昆虫较多,阴雨天气几乎没有见到昆虫访花。两种杜鹃花的访花昆虫只有中华蜜蜂(Apisceranacerana),它们访花所得的酬物为花蜜和花粉,是有效的传粉者。但中华蜜蜂访花频率不高且在每个花朵上的停留时间短。昆虫访花时段集中在8:00—15:00,15:00以后昆虫稀少,到17:00以后基本见不到访花昆虫。
对传粉昆虫的观察,发现两种杜鹃花共有传粉昆虫为中华蜜蜂(Apisceranacerana) (图1:C),这说明在自然的状态下,两种杜鹃花也有发生杂交的可能性。这为我们进行人工授粉,提供了一定的理论依据。
2.3 花粉活力与柱头可授性测定
羊踯躅和日本杜鹃在花瓣未开放前1~2天,花粉已从花粉囊散出。花粉活力在开花后24~72小时达到最高,在120 h之后逐渐下降,192小时之后,仍有一部分具有活力。在室温下,两种杜鹃花的花粉寿命均可达72小时以上。

A. 羊踯躅[Rhododendron molle (Bl.) G. Don]花朵; B. 日本杜鹃(R.molle subsp.japonicum)花朵; C. 蜜蜂传粉;D. 果实; E. 种子; F. 种子萌发; G. 羊踯躅幼苗;H. 杂交F1代幼苗;I. 日本杜鹃幼苗
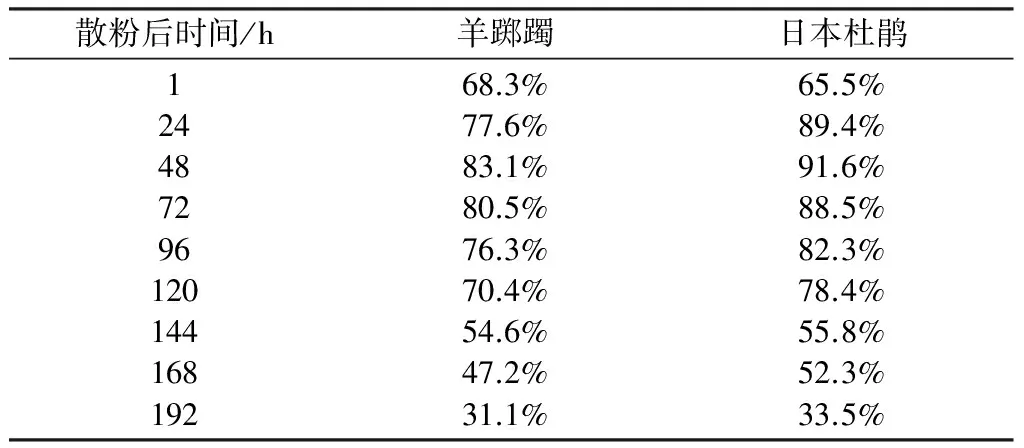
散粉后时间/h羊踯躅日本杜鹃12448729612014416819268.3%77.6%83.1%80.5%76.3%70.4%54.6%47.2%31.1%65.5%89.4%91.6%88.5%82.3%78.4%55.8%52.3%33.5%
羊踯躅和日本杜鹃的柱头均在开花前一天已具有过氧化物酶活性。羊踯躅的柱头在开花后2~6天逐渐膨大,第3天具强过氧化物酶活性,第6天花瓣及花丝开始脱落,柱头已没有弱过氧化物酶活性,第7天柱头枯萎;日本杜鹃在开花2~6天柱头逐渐膨大,2~3天具强过氧化物酶活性,第5天,花瓣开始脱落,柱头仍有过氧化物酶活性,6~7天柱头枯萎。

表2 两种杜鹃花柱头可授期的联苯胺-过氧化氢法检测结果
注:+ + + 表示具有最强的可授性,+ + 表示具有较强的可授性,+ 表示具有可授性,- 表示没有可授性。
通过花粉活力和柱头可授性的研究,为我们提供了最佳的采收花粉和进行人工授粉的时间,为了避免花粉及柱头的污染,我们在开花前采收花粉和进行人工授粉,因为此时花粉已经具有活力,且柱头具有可授性,花粉在柱头上也可存活72 h以上,能保证授粉的成功。
2.4 繁育系统研究
我们共得到7个组合的实验结果,结果如表3所示。繁育系统的研究结果表明,在自然状态下,两种花的结实率都较低,且羊踯躅的结实率低于日本杜鹃。将尚未开放的花蕾直接套袋,不作任何处理,羊踯躅和日本杜鹃的结实率均为0,说明羊踯躅和日本杜鹃非自花授粉。去雄,自花授粉,套袋,二者的结实率分别为33.3%和29.4%,表明羊踯躅和日本杜鹃自花授粉亲和但需要媒介。去雄不套袋,二者结实率分别为8.6%和15.7%,表明吸引昆虫授粉的可能是柱头上的黏液而非花粉。同株自花授粉和不同株异花授粉,日本杜鹃的结实率也略高于羊踯躅。异种异花授粉,羊踯躅为母本结实率为25.0%,日本杜鹃为母本的结实率为37.1%,说明这两种杜鹃能够杂交成功。羊踯躅和日本杜鹃种间正反交有一定差异,日本杜鹃为父本时低于其反交组合。去雄套网,羊踯躅和日本杜鹃的结实率皆为0,说明二者不是风媒花。

表3 六种不同人工授粉组合的结实率
2.5 种子萌发实验
2014年11—12月采收繁育系统研究所得果实。羊踯躅、日本杜鹃和杂交种果实均为圆柱状矩圆形蒴果(图1:D)。羊踯躅蒴果长2.1~3.5 cm,每个蒴果平均有258.5粒种子,千粒重0.37 g。日本杜鹃蒴果长1.4~3.0 cm,每个蒴果平均有107.2粒种子,千粒重0.25 g。杂交种(日本杜鹃(♀)×羊踯躅(♂))蒴果长2.5~4.2 cm,每个蒴果平均有548.6粒种子,千粒重0.29 g。三种果实均为5心室,有细柔毛和疏刚毛,种子扁平,黄褐色(图1:E)。将种子置于室温下保存,2015年4月进行种子萌发实验(图1:F)。

表 4 种子的发芽率
将3种种子用培养皿萌发法进行种子萌发实验,播种后第9天,日本杜鹃开始发芽,另外两种种子均无萌发现象,直至播种后第11天,羊踯躅和杂交种才开始发芽。第15~19天,三种种子都进入萌发高峰期。到第22天,羊踯躅和日本杜鹃不再萌发,第24天,杂交种子也停止萌发。整个萌发过程持续了约15天。最终,羊踯躅的平均萌发率为71.7%,日本杜鹃的平均萌发率较羊踯躅高,达到83.7%,杂交种子的平均萌发率为78%,说明杂交种子具有良好的萌发能力。
种子萌发实验结束后,将培养皿中的小苗转移至腐叶土中栽种,置于温室大棚内培养。五个月后,在同样环境条件下,羊踯躅幼苗大量死亡(图1,G),日本杜鹃幼苗部分出现叶片发黄(图1:I),而杂交后代幼苗生长状况较为良好(图1:H),经统计,羊踯躅幼苗的成活率为2.8%,日本杜鹃幼苗的成活率为10.4%,而杂交后代的成活率达到20.3%,杂交后代明显表现出优于两亲本的环境适应性。杜鹃属于木本植物,生长缓慢,杂交后代的其他表现性状仍需进一步研究。
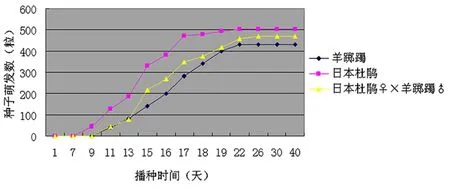
图 2 三种杜鹃种子的萌发情况
3 讨 论
3.1 花部特征与昆虫的传粉行为
我们发现羊踯躅和日本杜鹃的有效传粉昆虫为中华蜜蜂,其他昆虫虽有访花活动,但仅停留在花瓣上,并未出现有效的传粉行为。羊踯躅的花朵为黄色,日本杜鹃的花朵为橙色,这与其他学者讨论的蜂类喜欢黄色的结论相吻合。羊踯躅和日本杜鹃的花冠都为5裂,且其中一个裂片的基部都有红色、黄色斑点,这些斑点是引导昆虫采蜜进而完成传粉行为的蜜导[14]。
羊踯躅和日本杜鹃的花药在开花前1~2天就已散出,而花柱在开花前1天只分泌出极少的黏液,花药散粉初期花柱尚未完全成熟,即雌雄异熟。开花后,二者的花柱均比花药长,呈现出空间上的分离,即雌雄异位。雌雄异熟和雌雄异位是促进异交,避免单花自交的重要原因[15]。单花去雄套袋的实验结果也印证了这一点。
3.2 花粉活力与结实率
羊踯躅和日本杜鹃的花粉活力在开花第2天达到最高,随后逐渐降低,但直至花末期,仍具有活力。羊踯躅的的花粉活力最高时达到83.1%,日本杜鹃的花粉活力最高时达到91.6%,而二者的自然结实率分别只有13.3%和25%。这与边才苗对云锦杜鹃[9],田晓玲对大喇叭杜鹃[10,12],白天对锈叶杜鹃[11]的研究中出现的高花粉活力与低自然结实率的结果一致,说明部分杜鹃的花粉活力与自然结实率关联性较小。
同时,耿兴敏等的研究结果显示,花粉活力与杂交亲和性没有相关性,杜鹃花杂交结实率与母本的选择密切相关[16]。
3.3 繁育系统研究与杜鹃花的保护
羊踯躅和日本杜鹃在自花授粉套袋时都结实,表明二者自花授粉亲和,这与Escaravage对R.ferrugineum,Stout对R.ponticum, Wheelwright对R.arboreum和Ono对R.semibarbatum的研究结果一致,杜鹃属的大部分杜鹃是自交亲和的,但授粉方式主要是以传粉媒介辅助的同株异花授粉[17-21]。羊踯躅和日本杜鹃的有效传粉昆虫只有中华蜜蜂。
羊踯躅和日本杜鹃的花粉粒具有粘性,花药呈丝状散出,很难通过风媒扩散,而去雄套网实验结果也表明二者并非风媒花。
综上所述,羊踯躅和日本杜鹃的传粉昆虫少,访花频率低且缺失风媒,这种花粉限制是造成二者自然结实率低的主要原因。在被子植物中,花粉限制对结实率的影响已被证明是普遍存在的。Burd检测了258种被子植物,发现约有62%的物种受到不同程度的花粉限制。对濒危植物来说,生境的破坏和破碎化、种群规模的缩减以及由此而引起的传粉者的缺乏将进一步加剧花粉限制的程度[23]。
通过自花和同种不同株异花人工授粉发现,两种杜鹃结实率都明显提高,因此我们可以通过人工授粉提高其结实率。
3.4 杜鹃花的杂交育种
杜鹃花品种(种)间的杂交结实率高低与花色无关,与株型和瓣型也无明显关系,而与品种(种)的亲缘关系密切相关[24]。羊踯躅和日本杜鹃同属于羊踯躅亚属,二者能够能过人工授粉得到能育的后代,跟二者之间亲缘关系较近有关。
杂交育种可以增加植物群体内的遗传多样性,形成合理稳定的遗传结构,新产生的杂种具有潜在的、更为全面的、在各种生境下的适应能力[25]。杂交育种一直都是培育植物新品种的一个重要手段,尤其是在草本花卉上。而木本花卉常规杂交育种的年份较长,所以常规杂交育种在木本花卉中开展并不多[26]。杜鹃属植物的人工杂交育种在国内还没有得到发展和普及,杜鹃花的育种工作与国外相比差距较大[27]。制定系统而持续的育种计划,对我国杜鹃属植物杂交进程的加快,育种水平的提高和新品种的开发意义重大。
4 小 结
本文对羊踯躅和日本杜鹃的传粉生物学和二者之间的杂交育种进行了初步的研究,虽然得到了一些结果,但还有很多问题有待解决。杜鹃花为木本花卉,通过人工杂交育种,得到杂交种子,从种子萌发到开花大约经历5~10年的时间。本文的研究,仅是得到的杂交种的小苗,研究初步显示,杂交种小苗适应能力高于其双亲,其它性状须经过数年以后生长开花,才能继续进行鉴定。
[1] 方瑞征.中国植物志:第57卷[M].北京:科学出版社,1996.
[2] 冯国楣.中国杜鹃花[M].北京:科学出版社,1988.
[3] 张长芹,冯宝钧,吕元林.杜鹃花属的杂交育种研究[J].云南植物研究,1998,20(1):94-96.
[4] 张长芹,罗吉凤,冯宝钧.杜鹃花新品种“朝晖”和“红晕”[J].园艺学报,2002,29(3):296.
[5] 张长芹,罗吉凤.杜鹃花新品种“金踯躅”和“紫艳”[J].园艺学报,2002,29(5):502.
[6] 刘小青,苏家乐,李畅,等.高山杜鹃新品种“富丽金陵”[J].园艺学报,2011,38(11):2237-2238.
[7] LESLIE A C.The internationalRhododendronregister & checklist[M]. London:Royal Horticultural Society,2004.
[8] 张敬丽,张长芹,吴之坤,等. 探讨种间传粉在杜鹃花属自然杂交物种形成中的作用[J].生物多样性,2007,15(6):658-665.
[9] 边才苗,金则新. 云锦杜鹃的开花动态与繁育系统研究[J].广西植物,2005,2:169-173.
[10] 田晓玲,马永鹏,张长芹,等. 杜鹃花繁殖生物学研究进展[J].南京林业大学学报,2011,35(3):124-128.
[11] 白天,关文灵,宋杰,等. 锈叶杜娟的开花特性及繁育系统研究[J].西部林业科学,2014,43(1):47-53.
[12] 田晓玲. 大喇叭杜鹃繁殖生物学研究[D].南京:南京林业大学,2011.
[13] 王明麻. 林木遗传育种学[M]. 北京:中国林业出版社,2001.
[14] 张长芹.杜鹃花[M].北京:中国建筑工业出版社,2003.
[15] 候勤正,段元文,司庆文,等.青藏高原晚期开花植物线叶龙胆的传份生物学[J].植物生态学报,2009(6):1156-1164.
[16] 耿兴敏,张超仪,罗凤霞,等. 中国野生杜鹃杂交结实性研究[J].江苏农业科学,2013,41(2):159-161.
[17] ESCARVAGE N, WAGNER J. Pollination effectiveness and pollen dispersal in aRhododendronferrugineum(Ericaceae) population [J].Plant Biology,2004,6:606-615.
[18] STOUT J C. Reproductive biology of the invasive exotic shrub,RhododdendronponticumL.(Ericaceae)[J]. Botanical Journal of the Linnean Society,2007,155:373-381.
[19] STOUT J C. Pollination of invasiveRhododdendronponticum(Ericaceae) in Ireland [J].Apidologie,2007,38:198-206.
[20] WHEELWRIGHT N T.DUKESHIRE E E,FONTAINE J B, et al. Pollinator limitation,autogamy and minimal inbreeding depression in insect-pollinated plants on a boreal island {J}.American Midland Naturalist,2006,155:19-38.
[21] Ono A, Dohzono I,Sugawara T.Bumblebee pollination and reproductive biology ofRhododendronsemibarbatum(Ericaceae) [J].Journal of Plant Research,2008,121:319-327.
[22] 季荣,谢宝瑜,杨冠煌,等.从有意引入到外来入侵:以意大利蜂ApismelliferaL.为例[J].生态学杂志,2003,22:70-73.
[23] BURD M. Bateman’s principle and plant reproduction: The role of pollen limitation in fruit and seed set[J]. Botanical Review,1994,60:83-139.
[24] 刘晓青,苏家乐,李畅,等. 杜鹃花杂交、自交及开放授粉结实性研究[J].上海农业学报,2010,26(4):145-148
[25] BURKE, J M,ARNOLD M L. Genetic and the fitness of hybrids [J]. Annual Review of Genetics,2001(35):31-52.
[26] 董佳丽,于晓英,盛桢桢. 木本花卉杂交育种研究进展[J].安徽农业科学,2015,43(17):246-248,254.
[27] 王定跃,刘永金,白羽清,等. 杜鹃属植物育种研究进展[J].安徽农业科学,2012,40(32):15622-15625,15627.
Pollination Biology and Crossbreeding Between Rhododendron molle (Bl.) G. Don and Rhododendron molle subsp. japonicum
Yang Canjiao1, Zheng Shuoli1, Yang Rongping1, Tian Wei2, Zhang Jingli1*
(1. College of Landscape and Horticulture, Yunnan Agricultural University,Kunming 650201, China; 2. School of Agriculture, Ludong University, Yantai 264025, China)
Natural hybrid is common inRhododendronspecies. Many new varieties ofRhododendronwere cultivated by breeding in China and abroad.R.molle(Bl.) G. Don andR.mollesubsp.japonicumwere used as experimental materials in this reseach,and the pollination biological characteristics, the interspecific crosses and selfing were carried out in order to study the compatibility and provide basics for cross breeding and new variety breeding ofRhododendronL.
RhododendronL.; pollination biology; breeding systems; compatibility
10.3969/j.issn.1006-9690.2016.06.012
2016-04-18
国家自然科学基金—云南野生常绿杜鹃的二倍体杂交物种形成及种群进化研究(31260079);国家自然科学基金—大白杜鹃与两种真菌的菌根合成在引种驯化中的作用(31200248)。
杨灿娇(1987—),女,在读硕士,研究方向为园林植物资源应用与创新。
*通讯作者: 张敬丽(1975—),女,博士,副教授,主要从事野生植物资源起源与应用研究。E-mail:jl200812@yeah.net
S685
A
1006-9690(2016)06-0047-06
猜你喜欢
中国科学探险(2022年8期)2022-12-31
心声歌刊(2020年4期)2020-09-07
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
小学生作文(低年级适用)(2019年4期)2019-04-29
山东农业科学(2018年10期)2018-11-05
铜仁学院学报(2018年7期)2018-09-08
小学生作文(中高年级适用)(2018年5期)2018-06-11
大科技·百科新说(2018年3期)2018-04-11
种子(2018年3期)2018-03-31
作物研究(2015年4期)2015-03-24
