
黄土高原旱地不同种植系统对土壤水稳性团聚体及碳氮分布的影响
2016-03-24 08:00魏艳春马天娥魏孝荣王昌钊郝明德西北农林科技大学资源环境学院陕西杨凌7200西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室陕西杨凌7200陕西出入境检验检疫局技术中心西安70068
农业环境科学学报 2016年2期
魏艳春,马天娥,魏孝荣,2*,王昌钊,郝明德,张 萌(.西北农林科技大学资源环境学院,陕西杨凌7200;2.西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌7200;.陕西出入境检验检疫局技术中心,西安70068)
黄土高原旱地不同种植系统对土壤水稳性团聚体及碳氮分布的影响
魏艳春1,马天娥1,魏孝荣1,2*,王昌钊3,郝明德1,张萌1
(1.西北农林科技大学资源环境学院,陕西杨凌712100;2.西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌712100;3.陕西出入境检验检疫局技术中心,西安710068)
摘要:以黄土高原连续进行了27年的长期定位试验为对象,研究了粮-草长周期轮作、粮-豆短周期轮作、玉米连作和小麦连作系统土壤团聚体及其碳氮分布特征,并分析了土壤碳氮分布与土壤团聚体及其碳氮含量之间的关系。结果表明:黄土高原旱作农田土壤中<0.053 mm团聚体含量最高,占土壤质量的35%;长周期轮作系统0~20 cm和20~40 cm土层土壤0.25~2 mm团聚体含量高于玉米连作、小麦连作和短周期轮作系统,而<0.053 mm团聚体含量低于这3种轮作系统,且长周期轮作系统土壤团聚体的平均重量直径和几何平均直径也较高。种植系统对团聚体有机碳和全氮分布的影响主要体现在0~20 cm土层土壤,长周期轮作系统土壤中>2 mm和0.25~2 mm团聚体有机碳含量显著高于其他种植系统,<0.25 mm团聚体有机碳含量与其他种植系统差异不显著。长周期轮作系统团聚体全氮含量均显著高于其他种植系统,碳氮比则呈现出相反的趋势。土壤总有机碳、氮含量与团聚体有机碳、氮含量呈极显著正相关关系。土壤有机碳和全氮含量的变化主要取决于0.25~2 mm和0.053~0.25 mm团聚体有机碳和全氮的变化,而且有豆科植物苜蓿长期参与的长周期轮作系统可以有效改善土壤结构,提高土壤和团聚体的有机碳和全氮含量。
关键词:黄土高原;旱作农田;种植系统;水稳性团聚体;有机碳;全氮
魏艳春,马天娥,魏孝荣,等.黄土高原旱地不同种植系统对土壤水稳性团聚体及碳氮分布的影响[J].农业环境科学学报, 2016, 35(2):305-313.
团聚体是土壤结构的重要组分,在土壤肥力维持、质量改良和生态功能调节方面发挥着重要作用[1-2]。大量研究结果表明,不同粒级的团聚体在提供植物氮、磷素营养方面有着不同的功能[3-4],如微团聚体有较强的养分储备能力,而大团聚体供应养分的能力较强[5-6]。在土壤有机碳循环方面,团聚体不但能对有机碳起到物理保护作用,减少其损失[7-8],而且可以增强新输入的有机碳在团聚体中的富集和积累,从而有利于土壤碳汇功能的增强[9-10]。由于土壤团聚体是在有机物质、植物根系、微生物活动等共同作用下形成的[11-13],土壤团聚体的大小及其中有机碳和养分的分布、周转均受这些因素的影响。
目前研究者对农田生态系统土壤团聚体及其碳氮分布的研究主要集中在不同管理措施的影响方面,并且积累了大量的数据和可靠的结果。有研究表明,保护性耕作尤其是免耕措施能提高土壤大团聚体的形成,并增加大团聚体有机碳含量[14]。这些耕作措施结合秸秆还田还可以通过改变土壤的湿度和温度、根系生长状况、作物残茬的数量和质量,影响土壤微生物量及其活性,最终影响团聚体形成和团聚体有机碳的动态[15]。孙汉印等[13]发现陕西关中平原地区秸秆还田减少了土壤微团聚体含量,增加了大团聚体和中微团聚体的含量;高会议等[16]的研究表明,黄土塬区黑垆土长期施肥可增加5~2 mm和2~1 mm水稳性团聚体的有机碳含量。这些研究为深入认识农田生态系统团聚体碳氮的分布和稳定性提供了重要的基础和依据,但是很少有研究比较不同种植系统土壤团聚体及其碳氮的分布特征。对于不同的种植系统,作物的地上部和地下部生物量、根系分布以及根系活动特征不同,必将对土壤团聚体以及团聚体有机碳和全氮分布产生显著影响。因此,分析不同种植系统土壤团聚体碳氮分布特征对于认识土壤碳氮变化规律、分析土壤养分保持特征和有效性、评价生态系统碳汇功能具有重要的指导意义和理论价值。
本研究以黄土高原旱作农田生态系统为对象,针对小麦连作、玉米连作、粮-豆3年短周期轮作和粮-草8年长周期轮作4种典型种植系统,研究了土壤团聚体分布特征以及团聚体碳氮含量的变化,分析了土壤碳氮分布与土壤团聚体及其碳氮含量之间的关系,以期为认识不同种植系统土壤团聚体碳氮分布及其对土壤总有机碳氮的贡献,为评价不同种植系统土壤碳汇功能及变化趋势,提供基础资料和科学依据。
1 材料与方法
1.1试验区概况
试验在中国科学院长武黄土高原农业生态试验站进行。试验站建于1984年,位于陕西省长武县,海拔1200 m,为典型的黄土高原沟壑区,土壤类型为黄盖黑垆土,成土母质为马兰黄土。气候类型为暖温带半湿润大陆性季风气候,年日照时数2 226.5 h,日照百分率51%,年总辐射483.7 kJ·cm-3,年均气温9.2℃,塬面全年>0℃的积温为3029℃,多年平均无霜期171 d;年降雨量为578 mm,年内降雨主要集中在7—9月,占全年降雨量的53%。试验开始时土壤有机碳含量为6.09 g·kg-1,全氮含量为0.80 g·kg-1,碱解氮含量为37.0 mg·kg-1,全磷含量为0.70 g·kg-1,速效磷含量为3.0 mg·kg-1,速效钾含量为129 mg·kg-1。
1.2试验设计
长武黄土高原农业生态试验站长期定位试验布设于1984年,包括不同作物(表1)种植系统和施肥处理,设置36个处理,3个重复,共108个小区,每个小区长10.3 m,宽6.5 m。本研究选取小麦连作、玉米连作、粮-草8年长周期轮作和粮-豆3年短周期轮作4种种植系统进行研究,其中小麦连作和玉米连作系统只种植冬小麦或玉米,粮-草8年轮作系统作物轮作方式为洋芋-小麦-小麦-小麦/苜蓿-苜蓿-苜蓿-苜蓿-苜蓿;粮-豆3年轮作系统作物轮作方式为豌豆-小麦-小麦+糜子。所有轮作系统一个轮作周期结束后重复相同的轮作顺序,进行下一个周期的轮作。选取各种植系统中相同的施肥处理(N为120 kg·hm-2、P2O5为26.2 kg·hm-2),所用氮肥为尿素,磷肥为过磷酸钙,于作物播种前撒施地表,翻入土壤。在每种作物收获季节将小区内作物全部收获,称量鲜重,同时取部分样品带回实验室用烘干法测定其含水量,利用小区作物的总鲜重和含水量估算小区地上部生物量。本研究选取的小麦或玉米连作系统各有3个重复,采样时种植的作物为小麦或者玉米;8年长周期轮作系统有24个重复(有8种不同的种植序列处理,这样在每年均会有该种植系统不同的作物,如第一年苜蓿、第二年苜蓿等),采样时种植的作物为洋芋、小麦或者苜蓿;3年短周期轮作系统有9个重复(有3种不同的种植序列处理,这样在每年均会有该种植系统不同的作物),采样时种植的作物为豌豆或者小麦。

表1 供试作物品种及生育期Table 1 Varieties and seeding and harvesting periods of crops studied
1.3土壤样品采集、分析和数据处理
于2011年9月播前采集所选处理0~20 cm和20~40 cm土层土壤样品,每个小区内“S”型布设5个采样点,采集原状土组成该小区的混合土壤样品,带回实验室风干。土壤水稳性团聚体分级采用改进的湿筛法进行[17-19],将水稳性团聚体分为>2、0.25~2、0.053~0.25 mm和<0.053 mm 4个级别。
原状土壤样品和团聚体样品研磨通过0.25 mm筛孔以供有机碳和全氮测定,有机碳用重铬酸钾外加热法测定,全氮用凯氏定氮法测定。
为了分析团聚体分布特征,计算土壤团聚体的平均质量直径(MWD)和几何平均直径(GMD):
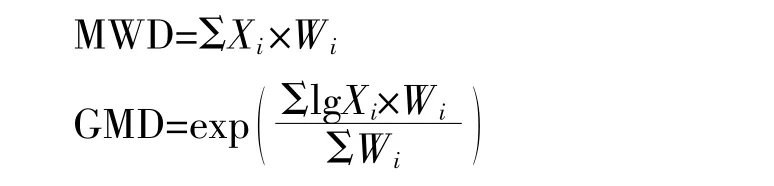
式中:Wi为某级别团聚体的质量百分数,%;Xi为该级别团聚体的平均直径,mm。
为了确定不同种植系统对土壤团聚体分布、土壤总有机碳和全氮、团聚体有机碳和全氮含量影响的差异,采用一元方差分析方法分析这些变量在不同种植系统之间差异的显著性,用简单线性回归方法分析土壤总有机碳、全氮与团聚体含量以及团聚体有机碳、全氮含量之间的关系。方差分析和线性回归分析采用JMP软件(v10.0)进行。
2 结果与分析
2.1团聚体分布特征
本研究中黄土高原旱作农田生态系统土壤不同级别团聚体含量之间差异显著(P<0.000 1)。所有处理0~20 cm和20~40 cm土层土壤中<0.053 mm团聚体占土壤质量的35%,而>2、0.25~2 mm和0.053~0.25 mm团聚体分别占土壤质量的20%、18%和27%,且这种含量顺序在不同种植系统呈现出相似的趋势(图1)。此外,土壤团聚体的含量受种植系统的影响,并与团聚体级别和土层有关(表2、图1)。0~20 cm和20~40 cm土层土壤中>2 mm团聚体含量在长周期和短周期轮作系统较高,小麦和玉米连作系统较低;0.25~2 mm团聚体也在长周期轮作系统最高,玉米连作系统最低;2个土层土壤0.053~0.25 mm团聚体均在玉米连作系统最低,而<0.053 mm团聚体均在连作系统较高,轮作系统较低(图1)。
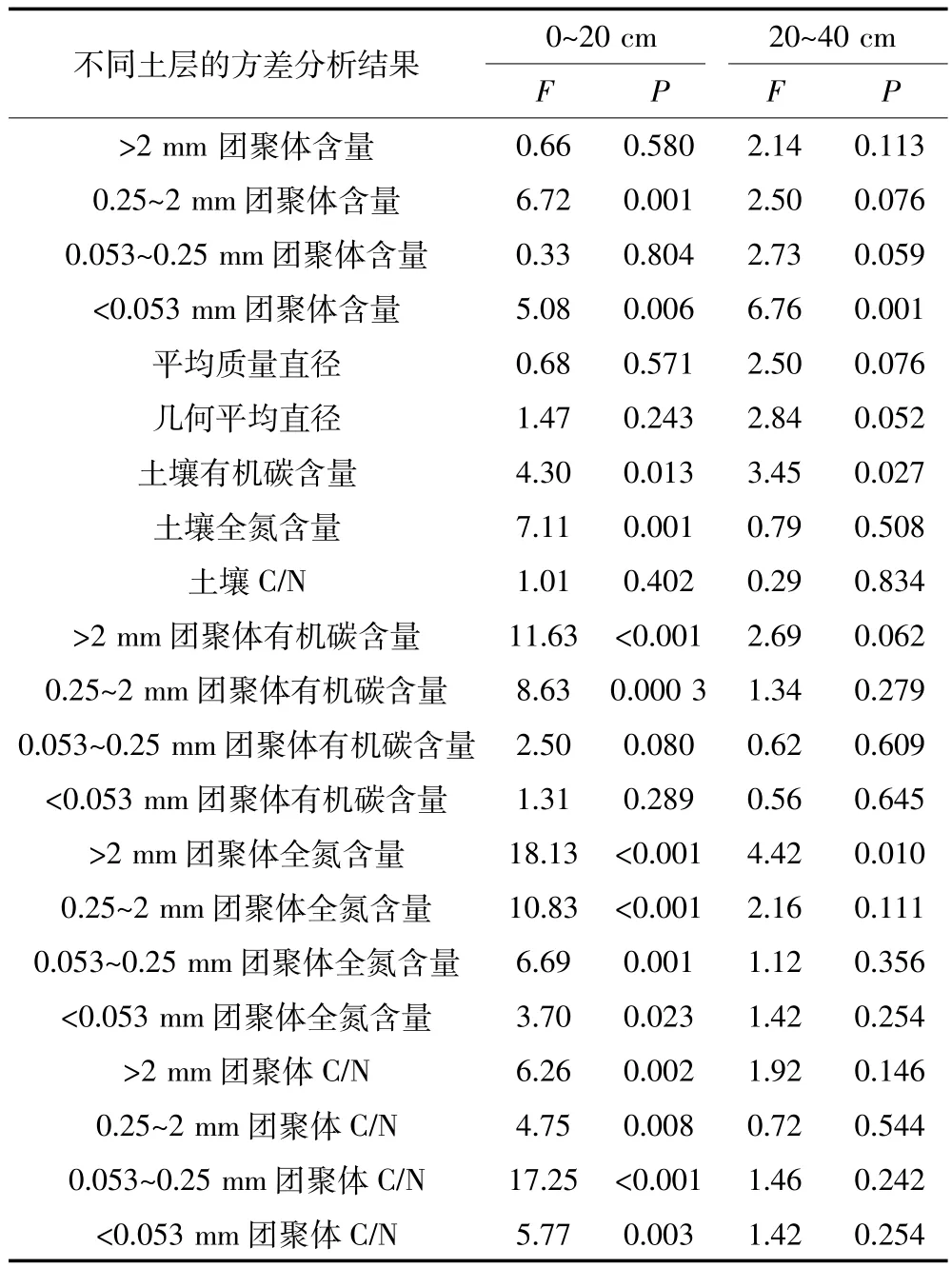
表2 不同种植系统对土壤团聚体、团聚体有机碳和全氮影响的方差分析结果Table 2 ANOVA of effects of cropping systems on aggregates and their OC and N content
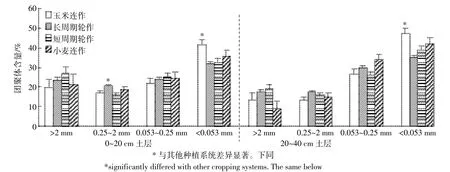
图1 不同种植系统团聚体含量分布特征Figure 1 Size distribution of aggregates as affected by cropping systems
土壤团聚体的平均质量直径和几何平均直径是表征土壤团聚体分布和土壤结构特征的重要指标,其值越高,表明土壤结构越好,越有利于植物生长[20]。本研究中,长周期轮作系统土壤团聚体平均质量直径在0~20 cm土层分别比玉米连作和小麦连作系统高19%和9%,在20~40 cm土层分别比这2种种植系统高27%和79%,但是在2个土层与短周期轮作相近。与此相似,长周期轮作系统的土壤团聚体的几何平均直径在0~20 cm土层分别比玉米连作和小麦连作系统高29%和24%,在20~40 cm分别高57%和69%,在2个土层与短周期轮作系统相近(图2),表明作物轮作可以显著改善土壤结构。
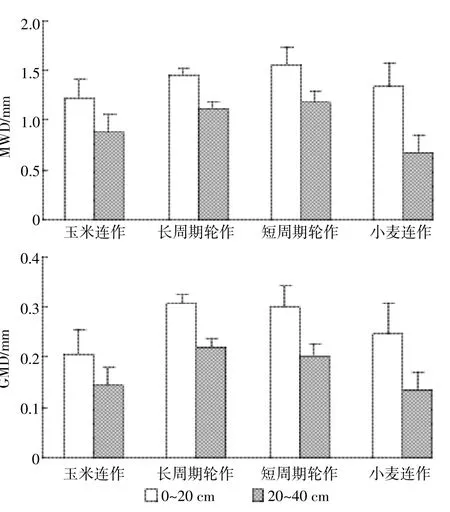
图2 不同种植系统团聚体平均质量直径(MWD)和几何平均直径(GMD)Figure 2 MWD and GMD of aggregates as affected by cropping systems
2.2不同种植系统土壤总有机碳和全氮分布特征
不同种植系统,由于作物的地上部和地下部生物量、根系分布以及根系活动的影响,其土壤有机碳和全氮分布特征存在显著差异。本研究中土壤C/N不受种植系统的影响,土壤有机碳和全氮含量则受其影响,而且种植系统的影响与土层深度有关(表2、图3)。玉米连作、小麦连作、短周期轮作和长周期轮作系统中土壤C/N在0~20 cm土层分别为11.3、11.3、11.1 和10.8(P=0.402),在20~40 cm土层分别为10.9、11.0、11.0和11.4(P=0.834)。土壤有机碳含量在0~20 cm和20~40 cm土层均以长周期轮作系统最高,玉米连作系统最低。长周期轮作系统土壤有机碳含量在0~20 cm分别比玉米连作、小麦连作和短周期轮作系统高13%、7%和13%(P=0.013),在20~40 cm土层分别比这3种种植系统高17%、5%和7%(P=0.027)。同样,长周期轮作系统土壤全氮含量在0~20 cm土层分别比这3种种植系统高18%、11%和15%(P<0.001),在20~40 cm土层分别高13%、3%和4%(P= 0.508),表明长周期轮作显著增加了土壤有机碳和全氮含量(图3)。
2.3土壤团聚体碳氮分布特征
不同级别团聚体有机碳、全氮含量以及C/N差异显著。土壤有机碳和全氮含量均以0.25~2 mm团聚体最高,<0.053 mm团聚体最低;土壤C/N以0.053~0.25 mm团聚体最高,<0.053 mm团聚体最低(图4)。0~40 cm土层土壤有机碳平均含量在>2、0.25~2、0.053~0.25 mm和<0.053 mm团聚体分别为7.0、8.0、6.9、6.1 g·kg-1,土壤全氮含量在这四个级别团聚体分别为0.86、0.96、0.82和0.79 g·kg-1,土壤C/N则分别为8.2、8.3、8.4和7.8(P=0.001)。
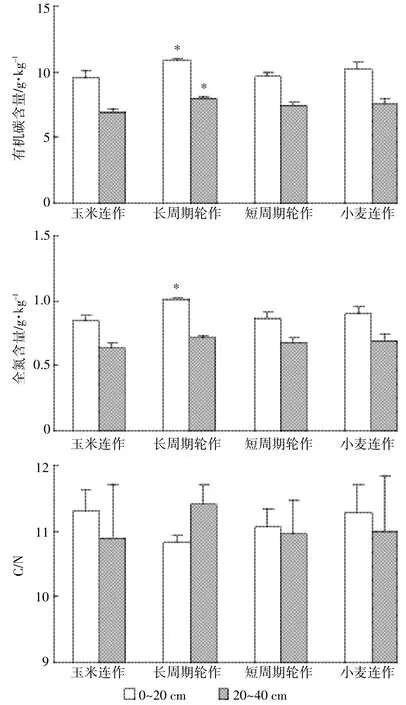
图3 不同种植系统土壤有机碳和全氮含量及C/NFigure 3 Soil OC, N and C/N as affected by cropping systems
本研究中不同种植系统显著影响0~20 cm土层土壤团聚体有机碳、全氮和C/N的分布,但是这种影响在20~40 cm土层不显著(在>2 mm团聚体全氮含量受到显著影响),表明种植系统对团聚体有机碳和全氮分布的影响主要体现在0~20 cm的表层土壤,20~40 cm土层土壤团聚体有机碳和全氮的分布不受种植系统的影响(表2、图4)。在0~20 cm土层,长周期轮作系统土壤>2 mm和0.25~2 mm团聚体有机碳含量分别比玉米连作、小麦连作和短周期轮作系统土壤高18%、9%、9%(P<0.000 1)和20%、13%、13%(P= 0.000 3),但是0.053~0.25 mm和<0.053 mm团聚体有机碳含量与其他种植系统差异不显著(P分别为0.079 5和0.288 8)。长周期轮作系统土壤团聚体全氮含量显著高于其他种植系统,在>2、0.25~2、0.053~0.25 mm和<0.053 mm团聚体分别比其他种植系统高14%~23%(P<0.000 1)、17%~28%(P<0.000 1)、13%~20%(P=0.001 4)和5%~10%(P=0.022 8)。不同种植系统土壤团聚体C/N却呈现出与有机碳和全氮相反的趋势,即长周期轮作系统土壤团聚体C/N显著低于其他种植系统,在四个级别团聚体依次比其他种植系统低0.33~0.54(P=0.002 1)、0.27~0.52(P=0.008 2)、0.54~0.68(P<0.000 1)和0.37~0.43(P=0.003 2)。
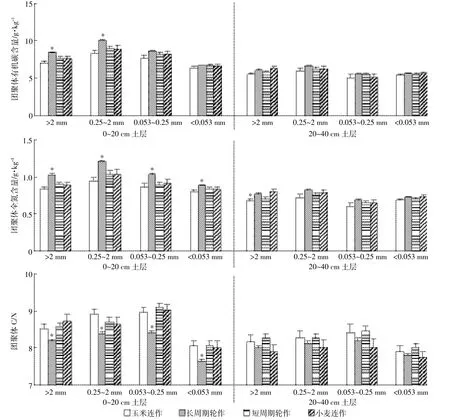
图4 不同种植系统团聚体有机碳和全氮含量以及C/N分布特征Figure 4 Distribution of organic carbon,total nitrogen and C/N in aggregates as affected by cropping systems
2.4土壤有机碳氮分布与团聚体分布和团聚体有机碳氮的关系
土壤总有机碳和全氮含量均随土壤中>2 mm和0.25~2 mm团聚体含量的增加显著增加(P<0.000 1),而随0.053~0.25 mm和<0.053 mm团聚体含量增加显著降低(P<0.000 1)。土壤总有机碳含量与团聚体有机碳含量呈极显著增加的线性关系(P<0.000 1),且0.25~2 mm和0.053~0.25 mm团聚体的斜率较大;土壤全氮与团聚体全氮含量呈现出相似的线性关系,也表现为0.25~2 mm和0.053~0.25 mm团聚体的斜率较大(表3)。
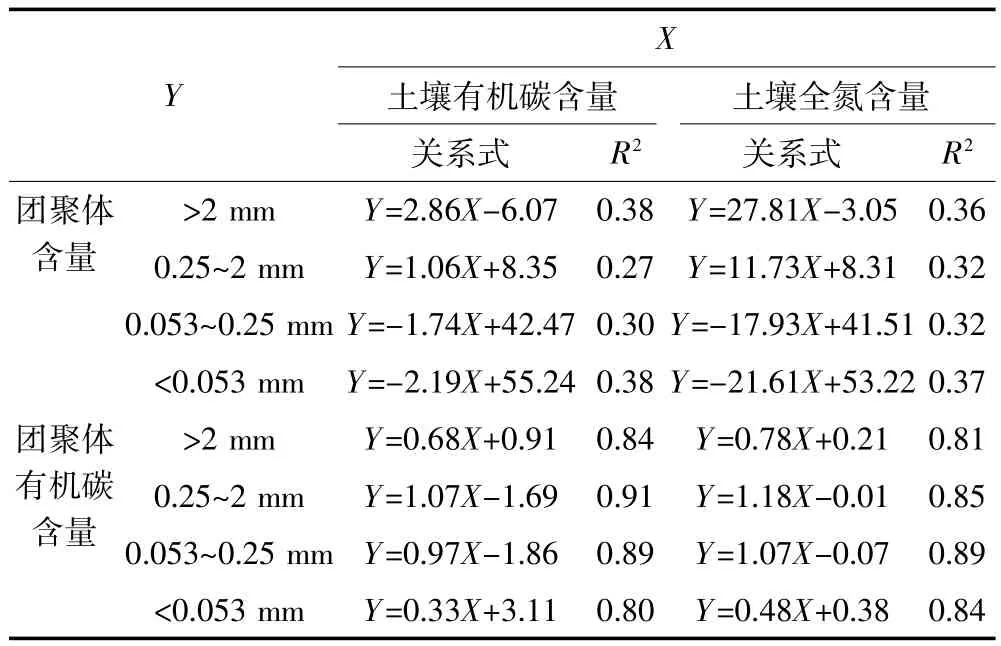
表3 土壤有机碳和全氮含量与土壤团聚体含量和团聚体有机碳含量之间的关系(n=78,P<0.001)Table 3 Linear relationships of soil OC and N content with aggregate content and OC content in aggregates(n=78,P<0.001)
3 讨论
各种植系统由于作物和轮作周期不同,植物根系分布、根系分泌物种类和数量、根系周转特征等有所差异,从而对土壤微环境,特别是土壤微生物性质产生影响,并影响土壤团聚体的分布特征[11,13]。本研究结果表明,长周期轮作系统有利于提高土壤中0.25~2 mm级别团聚体含量,而这部分团聚体在土壤肥力维持、物理性状调节、生物学性质改良以及对有机碳固持方面起着重要作用[18,21]。因此,有苜蓿参与的长周期轮作系统对土壤团聚体分布特征的影响可能是其改善土壤性质和功能的重要环节。与单一作物连作系统(小麦连作和玉米连作系统)相比,长周期轮作系统有3种植物参与,土壤中根系分泌物种类增加,而且不同植物根系的替换有利于土壤中微生物多样性的维持[22],这些因素均有利于土壤大团聚体的形成和土壤结构的改良[23]。另一方面,苜蓿具有较为庞大的根系,其根系在土壤中的穿插和分割作用增加了土壤孔隙度,可形成细小开放通路,使水分保留在土壤浅层,逐渐改善土壤水分和养分的贮藏,从而改善土壤结构和物理性状[24-25]。虽然短周期轮作系统中也有豆科植物(豌豆)参与,但是豌豆的生物量远远小于苜蓿,其对土壤物理性状的改善作用有限[26]。此外,在长周期轮作系统,苜蓿参与轮作的时间(5年/8年)远远高于短周期轮作系统中豆科植物豌豆参与轮作的时间(1年/ 3年),因此有苜蓿长期参与的长周期轮作系统土壤团聚体的平均质量直径和几何平均直径也均高于连作系统,并和短周期轮作系统相近。这也表明,对于黄土高原旱作农田生态系统,轮作系统对土壤结构能起到一定的改善作用,尤其是有苜蓿参与的长周期轮作系统,其改善作用更为明显。
长周期轮作系统可以显著增加土壤有机碳和全氮含量,可能与该系统较高的地上部生物量有关。本研究中长周期轮作系统平均地上部生物量分别比短周期轮作、小麦连作和玉米连作高91%、76%和50%,其通过光合作用固定并向土壤输送的碳也多,而且通过残留物归还到土壤中的有机物质也较多,有利于土壤有机碳和氮的提高。此外,长周期轮作系统土壤有机碳和全氮的增加还与苜蓿的长期参与有关。该种植系统中,轮作周期里有5年/8年为苜蓿,而苜蓿的生物量远远高于其他种植系统的植物,尽管研究区农田生态系统地上部生物量均被收获,但是苜蓿收获后残留的根茬以及掉落在土壤中的残余物也高于其他种植系统,其对土壤有机物质的补充量较大。同时,研究区苜蓿的根系生物量较大,通过根系归还到土壤中的有机物也较多[27],因而其参与的种植系统的土壤有机碳和全氮也较高。另一方面,苜蓿为豆科植物,具有较强的固氮功能,其根瘤从大气中固定的氮素储存在土壤中,不但有利于土壤氮素的积累,而且这种氮素的积累还能促进该种植系统其他植物的生长和对碳的吸收固定,从而增加土壤有机碳含量[28-29]。因此,有苜蓿参与的种植系统可以显著提高土壤的碳汇功能,这与Janzen等[30]在加拿大草原和Franzluebbers等[31]在美国南山麓地区的研究结果一致。
不同种植系统显著影响0~20 cm土层土壤团聚体有机碳和全氮以及C/N的分布,可能与农田生态系统不同植物的根系活动范围主要集中在表层土壤有关。如张春霞等[27]发现甘肃陇东地区苜蓿在0~20 cm的根系生物量是20~40 cm土层的2~4倍,成向荣等[32]在陕北农牧交错带地区也观测到了相似的结果。此外,虽然作物根系在20~40 cm土层也有分布,但是深层土壤根系周转较慢[33],其对土壤团聚体有机碳和全氮分布的影响较小。本研究中不同级别团聚体有机碳和全氮的分布特征也与其他研究得到的结果相似。如郭菊花等[34]对江西红壤性水稻土团聚体有机碳的研究表明,有机碳主要分配在2~0.25 mm的大团聚体上;Du等[14]对中国北部平原的研究表明,土壤有机碳积累量与2~0.25 mm团聚体有机碳含量密切相关,同时发现粒径较大的团聚体比粒径较小的团聚体含有更多的碳、氮;苗淑杰等[12]的研究表明<0.053 mm团聚体中有机碳含量最低。团聚体中土壤有机碳和全氮的这种分布模式可能与团聚体形成过程中对有机碳和全氮的富集以及形成后对有机碳和全氮的保护作用有关[10,18,21,35]。此外,有苜蓿长期参与的种植系统可以显著促进表层(0~20 cm)土壤有机碳和全氮的积累,而且有机碳和全氮的积累主要体现在大团聚体上。这与Hajabbasi等[36]的研究结果一致,他们发现轮作系统中加入豆科植物有利于土壤碳氮的积累和团聚体颗粒的聚集。Wei等[19]和Qiu等[37]也发现,黄土高原地区植被恢复后0~20 cm土层土壤有机碳的积累主要体现在大团聚体上。本研究中土壤C/N不受种植系统的影响,但是长周期轮作系统团聚体C/N低于其他种植系统,主要是苜蓿固氮造成的土壤全氮增加幅度大于有机碳积累幅度引起的,这也可以从长周期轮作系统团聚体全氮与其他种植系统的差异大于有机碳与其他种植系统的差异得到验证。这些结果也表明,团聚体碳氮特征能更详细和敏感地反映种植系统对土壤碳氮分布和相关特征的影响。
相关分析结果表明,土壤中有机碳和全氮的变化主要取决于0.25~2 mm和0.053~0.25 mm团聚体有机碳和全氮的变化。在土壤中,新输入的有机碳和全氮一般先与土壤微团聚体结合[9,38],然后通过团聚作用被大团聚体结合而固持起来[10,39]。本研究中长周期轮作系统增加的有机碳和全氮将先进入到微团聚体中,之后逐渐向大团聚体转移并被储存起来。土壤团聚体对有机碳和全氮有着一定的物理保护作用[34],长周期轮作系统通过提高土壤大团聚体含量,增强了对土壤中原有的和新输入的有机碳和全氮的保护作用,促进了土壤中有机碳和全氮的积累。本研究结果与其他研究者在黄土高原地区关于植被恢复后土壤有机碳和团聚体分布之间关系的研究结果一致[37],可能这也是长周期轮作系统土壤有机碳和全氮积累的一个重要途径。
4 结论
(1)黄土高原旱作农田长周期轮作能显著提高土壤大团聚体数量和团聚体的平均质量直径和几何平均直径,可以显著改善土壤结构。
(2)种植系统对土壤和团聚体有机碳、全氮分布的影响主要体现在0~20 cm的表层土壤。与连作系统和短周期轮作系统相比,长周期轮作系统显著提高了0~20 cm土层土壤>2 mm和0.25~2 mm团聚体有机碳含量,并且提高了所有级别团聚体全氮含量,降低了土壤C/N。
(3)土壤总有机碳、全氮含量与团聚体有机碳、全氮含量极显著相关,而且土壤有机碳和全氮对种植系统的响应主要取决于0.25~2 mm和0.053~0.25 mm团聚体有机碳和全氮的响应。
参考文献:
[1] Lal R. Physical management of soils of the tropics:Priorities for the 21 century[J]. Soil Science, 2000, 165(3):191-207.
[2] Six J, Bossuyt H, Degryze S, et al. A history of research on the link between(micro)aggregates, soil biota, and soil organic matter dynamics [J]. Soil and Tillage Research, 2004, 79(1):7-31.
[3]邱莉萍,张兴昌,张晋爱.黄土高原长期培肥土壤团聚体中养分和酶的分布[J].生态学报, 2006, 26(2):364-372. QIU Li-ping, ZHANG Xing-chang, ZHANG Jin-ai. Distribution of nutrients and enzymes in Loess Plateau soil aggregates after long-term fertilization[J]. Acta Ecologica Sinica, 2006, 26(2):364-372.
[4]孙天聪,李世清,邵明安.长期施肥对褐土有机碳与氮素在团聚体中分布的影响[J].中国农业科学, 2005, 38(9):1841-1848. SUN Tian-cong, LI Shi-qing, SHAO Ming-an. Effects of long-term fertilization on distribution of organic matters and nitrogen in cinnamon soil aggregates[J]. ScientiaAgricultura Sinica, 2005, 38(9):1841-1848.
[5] Kong A Y Y, Six J, Bryant D C, et al. The relationship between carbon input, aggregation, and soil organic carbon stabilization insustainable cropping systems[J]. Soil Science Society of America Journal, 2005, 69 (4):1078-1085.
[6]王丽,李军,李娟,等.轮耕与施肥对渭北旱作玉米田土壤团聚体和有机碳含量的影响[J].应用生态学报, 2014, 25(3):759-768. WANG Li, LI Jun, LI Juan, et al. Effects of tillage rotation and fertilization on soil aggregates and organic carbon content in corn field in Weibei Highland[J]. Chinese Journal of Applied Ecology, 2014, 25(3):759-768.
[7] Blanco-Canqui H, Lal R. Mechanisms of carbon sequestration in soil aggregates[J]. Critical Reviews in Plant Sciences, 2004, 23(6):481-504.
[8] César P, Denis C M, José M F, et al. Physical, chemical, and biochemical mechanisms of soil organic matter stabilization under conservation tillage systems:A central role for microbes and microbial by-products in C sequestration[J]. Soil Biology and Biochemistry, 2013, 57:124-134.
[9]韩志卿,韩志才,张电学,等.不同施肥制度下褐土微团聚体碳氮分布变化及其对肥力的影响[J].华北农学报, 2008, 23(4):190-195. HAN Zhi-qing, HAN Zhi-cai, ZHANG Dian-xue, et al. Distributions of organic carbon and nitrogen in cinnamon soil micro-aggregates and effect on soil fertility under the different fertilization regimes[J]. Acta A-griculturae Boreali-Sinica, 2008, 23(4):190-195.
[10] Six J, Paustian K, Elliott E T, et al. Soil structure and organic matter:I. distribution of aggregate-size classes and aggregate-associated carbon [J]. Soil Science Society of America Journal, 2000, 64(2):681-689.
[11] Du Z L, Ren T S, Hu C S, et al. Transition from intensive tillage to notill enhances carbon sequestration in microaggregates of surface soil in the North China Plain[J]. Soil and Tillage Research, 2015, 146(Part A):26-31.
[12]苗淑杰,周连仁,乔云发,等.长期施肥对黑土有机碳矿化和团聚体碳分布的影响[J].土壤学报, 2009, 46(6):1068-1075. MIAO Shu-jie, ZHOU Lian-ren, QIAO Yun-fa, et al. Organic carbon mineralization and carbon contribution in aggregates as affected by long-term fertilization[J]. Acta Pedologica Sinica, 2009, 46(6):1068-1075.
[13]孙汉印,姬强,王勇,等.不同秸秆还田模式下水稳性团聚体有机碳的分布及其氧化稳定性研究[J].农业环境科学学报, 2012, 31 (2):369-376. SUN Han-yin, JI Qiang, WANG Yong, et al. The distribution of waterstable aggregate-associated organic carbon and its oxidation stability under different straw returning modes[J]. Journal of Agro-Environment Science, 2012, 31(2):369-376.
[14] Du Z L, Ren T S, Hu C S, et al. Soil aggregate stability and aggregateassociated carbon under different tillage systems in the North China Plain[J]. Journal of Integrative Agriculture, 2013, 12(11):2114-2123.
[15] Garcia-Franco N, Martinez-Mena M, Goberna M, et al. Changes in soil aggregation and microbial community structure control carbon sequestration after afforestation of semiarid shrublands[J]. Soil Biology and Biochemistry, 2015, 87:110-121.
[16]高会议,郭胜利,刘文兆,等.不同施肥处理对黑垆土各粒级团聚体中有机碳含量分布的影响[J].土壤学报, 2010, 47(5):931-938. GAO Hui-yi, GUO Sheng-li, LIU Wen-zhao, et al. Effect of fertilization on organic carbon distribution in various fractions of aggregates in cliche soil[J]. Acta Pedologica Sinica, 2010, 47(5):931-938.
[17] Cambardella C A, Elliott E T. Carbon and nitrogen distributions in aggregates from cultivated and grassland soils[J]. Soil Science Society of America Journal, 1993, 57(4):1071-1076.
[18] Qiu L P, Wei X R, Zhang X C, et al. Soil organic carbon losses due to land use change in a semiarid grassland[J]. Plant and Soil, 2012, 355 (1/2):299-309.
[19] Wei X R, Li X Z, Jia X X, et al. Accumulation of soil organic carbon in aggregates after afforestation on abandoned farmland[J]. Biology and Fertility of Soils, 2013, 49(6):637-646.
[20]周虎,吕贻忠,杨志臣,等.保护性耕地对华北平原土壤团聚体特征的影响[J].中国农业科学, 2007, 40(9):1973-1979. ZHOU Hu, LÜ Yi-zhong, YANG Zhi-chen, et al. Effects of conservation tillage on soil aggregates in Huabei Plain, China[J]. Scientia Agricultura Sinica, 2007, 40(9):1973-1979.
[21] Wei X R, Shao M A, Gale W J, et al. Dynamics of aggregate-associated organic carbon following conversion of forest to cropland[J]. Soil Biology and Biochemistry, 2013, 57:876-883.
[22] Freixo A A, Machado P L, Santos H P, et al. Soil organic carbon and fractions of a Rhodic Fcerralsol under the influence of tillage and crop rotation systems in southern Brazil[J]. Soil and Tillage Research, 2002, 64(3/4):221-230.
[23] Gale J, Cambardella C A, Bailey T B. Root-derived carbon and the formation and stabilization of aggregates[J]. Soil Science Society of America Journal, 2000, 64(1):201-207.
[24]胡发成.种植苜蓿改良培肥地力的研究初报[J].草业科学, 2005, 22 (8):47-49.HU Fa-cheng. Initial research report on soil fertility improvement by planting Medicago sativa[J]. Pratacultural Science, 2005, 22(8):47-49.
[25]李裕元,邵明安.黄土高原北部紫花苜蓿草地退化过程与植物多样性研究[J].应用生态学报, 2005, 16(12):2321-2327. LI Yu-yuan, SHAO Ming-an. Degradation process and plant diversity of alfalfa grassland in North Loess Plateau of China[J]. Chinese Journal of Applied Ecology, 2005, 16(12):2321-2327.
[26]郭胜利,吴金水,党廷辉.轮作和施肥对半干旱区作物地上部生物量与土壤有机碳的影响[J].中国农业科学, 2008, 41(3):744-751. GUO Sheng-li, WU Jin-shui, DANG Ting-hui. Effects of crop rotation and fertilization on aboveground biomass and soil organic C in semiarid region[J]. Scientia Agricultura Sinica, 2008, 41(3):744-751.
[27]张春霞,郝明德,王旭刚,等.黄土高原地区紫花苜蓿生长过程中土壤养分的变化规律[J].西北植物学报, 2004, 24(6):1107-1111. ZHANG Chun-xia, HAO Ming-de, WANG Xu-gang, et al. Study on soil nitrogen and fertility distribution characteristics in alfalfa field in gully region of the Loess Plateau[J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(6):1107-1111.
[28] Liu J X, Price D T, Chen J M. Nitrogen controls on ecosystem carbon sequestration:A model implementation and application to Saskatchewan, Canada[J]. Ecological Modelling, 2005, 186(2):178-195.
[29] Thomas R Q, Canham C D, Weathers K C, et al. Increased tree carbon storage in response to nitrogen deposition in the US[J]. Nature Geoscience, 2010, 3(1):13-17.
[30] Janzen H H, Campbell C A, Izaurralde R C, et al. Management effects on soil C storage on the Canadian prairies[J]. Soil and Tillage Research, 1998, 47(3):181-195.
[31] Franzluebbers A J, Stuedemann J A. Soil-profile organic carbon and total nitrogen during 12 years of pasture management in the Southern Piedmont, USA[J]. Agriculture, Ecosystems and Environment, 2009, 129(1/2/3):28-36.
[32]成向荣,黄明斌,邵明安,等.紫花苜蓿和短花针茅根系分布与土壤水分研究[J].草地学报, 2008, 16(2):170-175. CHENG Xiang-rong, HUANG Ming-bin, SHAO Ming-an, et al. Root distribution and soil water dynamics of Medicago sativa L. and Stipabreviflora Griseb[J]. Acta Agrestia Sinica, 2008, 16(2):170-175.
[33]周梦华,程积民,万惠娥,等.云雾山本氏针茅群落根系分布特征[J].草地学报, 2008, 16(3):267-271. ZHOU Meng-hua, CHENG Ji-min, WAN Hui-e, et al. Distribution characteristics of the root system of the Stipabungeana Trin. community in Yunwu Mountain[J]. Acta Agrestia Sinica, 2008, 16(3):267-271.
[34]郭菊花,陈小云,刘满强,等.不同施肥处理对红壤性水稻土团聚体的分布及有机碳、氮含量的影响[J].土壤, 2007, 39(5):787-793. GUO Ju-hua, CHEN Xiao-yun, LIU Man-qiang, et al. Effects of fertilizer management practice on distribution of aggregates and content of organic carbon and nitrogen in red paddy soil[J]. Soils, 2007, 39(5):787-793.
[35] Garcia-Franco N, Albaladejo J, Almagro M, et al. Beneficial effects of reduced tillage and green manure on soil aggregation and atabilization of organic carbon in a Mediterranean agroeco system[J]. Soil and Tillage Research, 2015, 153:66-75.
[36] Hajabbasi M A, Fallahzade J. Aggregation, carbohydrate, total and particulate organic carbon changes by cultivation of an arid soil in Central Iran[R]. Brisbane: 19th World Congress of Soil Science, 2010.
[37] Qiu L P, Wei X R, Gao J L, et al. Dynamics of soil aggregate-associ -ated organic carbon along an afforestation chronosequence[J]. Plant and Soil, 2015, 391:237-251.
[38] Marriott E E, Wander M. Qualitative and quantitative differences in particulate organic matter fractions in organic and conventional farming systems[J]. Soil Biology and Biochemistry, 2006, 38(7):1527-1536.
[39] Six J, Elliott E T, Paustian K. Soil macroaggregate turnover and microaggregate formation:A mechanism for C sequestration under notillage agriculture[J]. Soil Biology and Biochemistry, 2000, 32(14):2099-2103.
WEI Yan-chun, MA Tian-e, WEI Xiao-rong, et al. Effects of cropping systems on distribution of water-stable aggregates and organic carbon and nitrogen in soils in semiarid farmland of the Loess Plateau[J]. Journal of Agro-Environment Science, 2016, 35(2):305-313.
Effects of cropping systems on distribution of water-stable aggregates and organic carbon and nitrogen in soils in semiarid farmland of the Loess Plateau
WEI Yan-chun1, MA Tian-e1, WEI Xiao-rong1,2*, WANG Chang-zhao3, HAO Ming-de1, ZHANG Meng1
(1.College of Resources and Environment, Northwest A&F University, Yangling 712100, China; 2.State Key Laboratory of Soil Erosion and Dry Land Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China; 3.Shaanxi Entry-Exit Inspection and Quarantine Bureau, Xi'an 710068, China)
Abstract:In this paper, we examined the distribution of water stable aggregates and organic carbon(OC)and nitrogen(N)in soils under different cropping systems, using a 27-year field experiment in the Loess Plateau. The relationships between OC and N contents in bulk soil and aggregates were also analyzed. Results showed that<0.053 mm aggregates, accounting for 35%, dominated the soil mass in 0~20 and 20~40 cm soil layers in the semiarid farmland of the Loess Plateau. The percentage of 0.25~2 mm aggregates was higher, but that of<0.053 mm aggregates was lower in the grass-grain rotation system than the other cropping systems. In the grass-grain rotation system, mean weight diameter and geometrical mean diameter of soil aggregates were also greater. The significant effects of cropping system on OC and N associated with aggregates mainly occurred in 0~20 cm layer. The OC content in>2 mm and 0.25~2 mm aggregates was significantly higher in grass-grain rotation system than in the other systems, while OC content in 0.053~0.25 mm and<0.053 mm aggregates did not differ amongbook=306,ebook=107different cropping systems. The grass-grain system showed higher N content and lower C/N ratio in each aggregate compared with other systems. The OC and N content in bulk soils was positively correlated with OC and N content associated with aggregates. These results suggest that the responses of OC and N in the bulk soil to cropping systems mainly depend on OC and N associated with 0.25~2 mm and 0.053~0.25 mm aggregates. Additionally, grass-grain rotation system has greater potential to improve soil structure and increase the content of OC and N associated with aggregates.
Keywords:Loess Plateau; semiarid farmland; cropping system; water-stable aggregates; organic carbon; total nitrogen
*通信作者:魏孝荣E-mail:xrwei78@163.com
作者简介:魏艳春(1991—),女,山东菏泽人,硕士研究生,从事土壤物质循环研究。E-mail:ycwei91@163.com
基金项目:国家自然科学基金项目(41271315,41571296);教育部新世纪优秀人才支持计划(NCET-13-0487);西北农林科技大学优秀青年人才科研专项(2014YQ007)
收稿日期:2015-09-11
中图分类号:S153.6
文献标志码:A
文章编号:1672-2043(2016)02-0305-09
doi:10.11654/jaes.2016.02.014
猜你喜欢
淄博师专论丛(2019年1期)2019-04-04
现代园艺(2017年13期)2018-01-19
植物资源与环境学报(2017年3期)2017-10-12
辽宁化工(2017年2期)2017-03-10
公民与法治(2016年3期)2016-05-17
公民与法治(2016年1期)2016-05-17
现代农业(2016年3期)2016-04-14
土壤与作物(2015年3期)2015-12-08
中国火炬(2015年7期)2015-07-31
郑州大学学报(理学版)(2012年4期)2012-03-25
