
匍匐茎克隆植物蛇莓的遗传多样性和遗传结构
2016-04-11 15:26刘春香李钧敏李红丽罗芳丽
江苏农业科学 2016年2期
关键词:遗传多样性
刘春香++李钧敏++李红丽++罗芳丽


摘要:采用8对简单重复序列(simple sequence repeat,SSR)引物对克隆植物蛇莓33个野生种群353个个体的遗传多样性及遗传结构进行分析。结果表明,参试的33个种群中的平均等位基因数(A)为2.841,平均多态性位点(PPL)为88.6%,平均期望杂合度(He)为0.494,平均Neis遗传多样性指数(h)为0.468。所有参试样本中共有223个基因型。该物种的克隆多样性和等位基因水平的变异均较高,各种群的平均基因型数(G)为6.758,平均克隆大小(N/G)为2.063,克隆多样性指数Simpson指数(D)为0.769,基因型均匀度(E)为0.769。基于分子的方差分析结果表明,蛇莓野生种群间和种群内的分化系数分别为0.55和0.45,这说明蛇莓种群间和种群内遗传分化均显著。STRUCTURE分析和基于Neis遗传一致度的UPGMA聚类结果表明,参试的33个野生种群可分为四大类群。Mantel相关性分析结果表明,种群间遗传距离与其地理距离不存在显著相关关系(r=0.052,P=0.150)。该研究结果说明蛇莓野生种群的遗传多样性高,所有种群均为多克隆种群,且基因型的局域性很强,其中82.6%的基因型为局部基因型,不存在广布基因型。物种水平的遗传差异主要表现在种群间,但在种群内也较高,这表明该物种在种群间和种群内在一定程度上能通过有性繁殖进行基因交流。蛇莓种群建群者的奠基者效应和所在位置生态因子对其遗传多样性和遗传结构的形成和进化具有重要的意义。
关键词:蛇莓;克隆生长;遗传多样性;简单重复序列;基因型多样性;遗传分化
中图分类号: Q945.79文献标志码: A文章编号:1002-1302(2016)02-0215-06
收稿日期:2015-01-16
基金项目:国家自然科学基金(编号:31200314、31470475);高等学校博士学科点专项科研基金新教师类资助(编号:20120014120001)。
作者简介:刘春香(1990—),女,湖南邵阳人,硕士研究生,主要从事克隆植物生态学研究。E-mail:chunxiangliu_1990@163.com。
通信作者:罗芳丽,女,四川都江堰人,博士,副教授,主要从事湿地生态学研究。E-mail:ecoluofangli@bjfu.edu.cn。遗传结构、遗传多样性和生活史特征等影响物种与其所在环境间的相互作用,也决定物种的进化潜势[1]。繁育系统是决定植物种群遗传结构的重要因素之一,而克隆植物一般兼具有性和无性2种繁殖方式,且这2种繁殖策略在一定程度上有权衡作用[2-3]。因此,研究克隆植物的遗传结构对深入了解植物种群遗传多样性的形成及进化机制具有重要意义[4-5]。
克隆生长是克隆植物所特有的生活史过程[6]。很多克隆植物通过形成根状茎、匍匐茎、鳞茎、珠芽、块茎等,能够迅速产生大量的无性后代个体即分株[7]。除少数分株可能存在概率极低的遗传突变以外,绝大多数分株的遗传组成与亲体分株(母株)完全相同[8]。由于母株可通过克隆整合向后代分株提供光合产物、水分和养分的支持[9-12],使得这些后代分株能够顺利地度过建立期,并迅速占据局部空间,进而可能形成由1个或几个基因型占优势的分株种群[13-15]。这些生态过程对克隆植物种群遗传结构和遗传多样性均可产生十分显著的影响[16]。
在自然界中,大多数克隆植物除了通过克隆生长进行无性繁殖之外,还能够通过产生种子等进行有性繁殖[8]。在1个或几个基因型占优势的分株种群中,由于相同基因型分株的大量存在,使得相同基因型个体自交的概率极高,从而影响其遗传多样性水平[8-16]。由于克隆植物的这些特性,人们最初普遍认为克隆植物种群的遗传多样性水平很低,遗传结构简单[17]。有研究表明,一些克隆植物的遗传多样性的确很低[17-18]。例如,在全球范围内,凤眼莲(Eichhornia crassipes)在非原产地的个体中约80%属于同一个基因型[14,19-21]。中国南方地区的空心莲子草(Alternanthera philoxeroides)很可能只存在1个基因型[22]。然而,很多克隆物种具有与非克隆植物相当的遗传多样性如甜樱桃(Prunus avium)、红球姜(Zingiber zerumbet)、大苦草(Vallisneria americana)和羊柴(Hedysarum laeve)等[23-27]。这是因为与非克隆植物相比,克隆植物生命周期长,所以即使非常少量的实生苗(即种子萌发所产生的幼苗)更新也能使克隆种群维持较高的遗传多样性[28]。蛇莓(Duchesnea indica Focke)为蔷薇科(Rosaceae)蛇莓属的多年生草本植物,主要分布在我国辽宁以南各省(市、区),常生长在山坡、河岸、草地及潮湿的地方[29]。该物种通过产生较长的地上匍匐茎表现出旺盛的克隆生长习性。该物种也可通过有性生殖产生种子,并进一步萌发形成实生苗。蛇莓是一种典型的游击型克隆植物,能够快速形成单一基因型的大种群。该物种还具有较强的花展示,即能通过产生花蜜来吸引传粉者,以增强有性繁殖[30]。近年来,对蛇莓的研究主要集中在化学活性成分、药理,克隆构型对环境的可塑性反应和小尺度克隆结构等方面[31-33]。本研究采用简单重复序列(simple sequence repeat,SSR)分子标记技术对全国范围内的33个蛇莓野生种群的353株个体的遗传多样性水平和遗传结构进行分析,旨在了解兼性克隆植物蛇莓的种群遗传多样性与遗传分化,探讨其遗传结构的特点,具体回答以下科学问题:克隆植物蛇莓种群的遗传多样性水平;蛇莓种群的遗传多样性主要发生在种群内还是种群间;蛇莓种群间遗传距离与其地理距离的相关关系。
1材料与方法
1.1植物采集
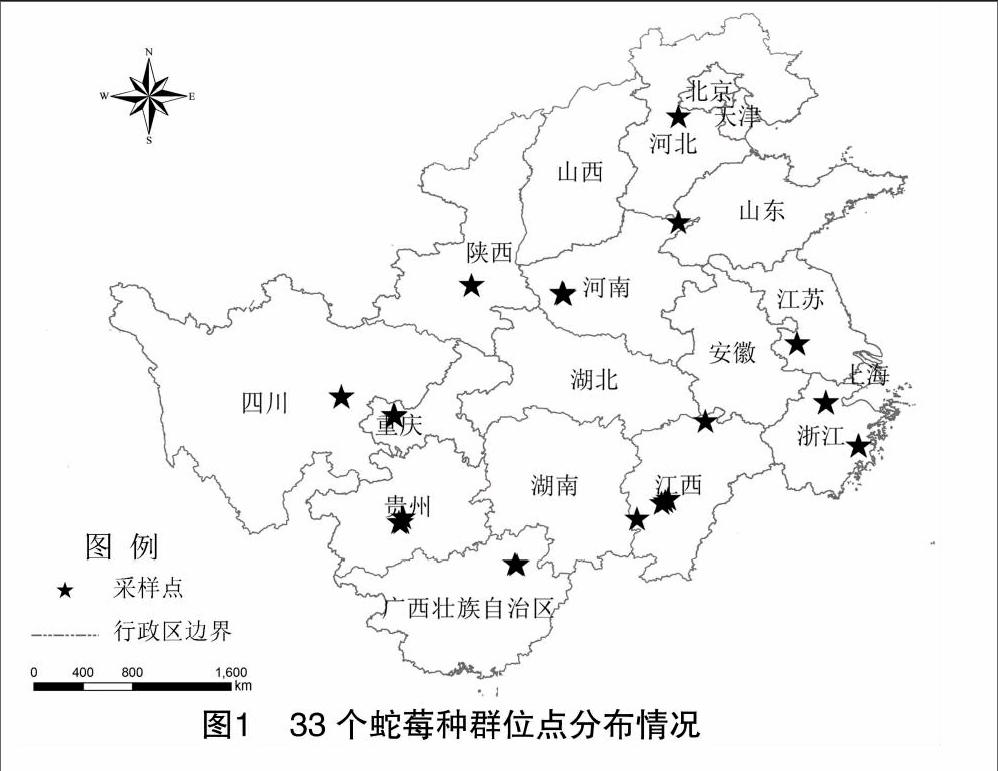
33个蛇莓野生种群分别采自广西、浙江、江西、湖南、贵州、河南、北京和陕西等10个省(市、区),具体信息见表1和图1。每个种群根据面积大小各采集5~17个样本,共353个,采集的植株在北京林业大学科技股份有限公司的温室中进行培养备用。
1.2DNA提取和PCR扩增
根据CTAB植物基因组DNA快速提取试剂盒(艾德莱生物,DN1401,北京)的说明方法提取幼嫩蛇莓叶片中的总DNA,采用NAS-99分光光度计(ACTGene,美国)确定总DNA的质量和浓度,并稀释至10 ng/μL备用。
蛇莓SSR引物的开发由北京阅微基因技术有限公司完成,从中筛选出8对清晰多态引物用于所有个体的PCR扩增,引物序列见表2。PCR反应体系为:8 μL 2.5 × Multiplex Buffer(MRP17,北京阅微基因技术有限公司,北京),0.4 μL 15 μmol/L TP-M13荧光接头(北京阅微基因技术有限公司),2 μL引物(5 μmol/L),0.2 μL 5 U/μL Fast Taq DNA聚合酶,2 μL DNA模板,7.6 μL无菌双蒸水。PCR扩增程序为:95 ℃ 预变性5 min;94 ℃变性30 s,56 ℃退火45 s,72 ℃延伸45 s,30个循环;94 ℃变性30 s,53 ℃退火45 s,72 ℃ 延伸 45 s,10个循环;72 ℃延伸12 min。PCR扩增在9 600 PCR扩增仪(美国应用生物系统公司)上完成,PCR扩增产物在3 730 xl DNA分析仪(美国应用生物系统公司)上进行基因分型分析。
1.3数据分析
人工判读PCR扩增产物的基因分型结果,记录目的条带的片段大小以构成SSR分子标记数据矩阵。使用Popgen 32软件[34]计算以下遗传多样性指数:每个位点的平均等位基因数(A)、预期杂合度(He)、Neis基因多样性指数(h)及Neis遗传一致度和遗传距离。计算实际基因型(即基株或克隆,所有位点基因型相同的植株为1个基株)总数(G)、平均克隆大小(N/G,基因型数/取样个体数)、局部基因型(只存在于1个群体中的基因型)比例(GL)、广布基因型(存在于75%以上群体中的基因型)比例(GW)。种群遗传多样性指数采用Simpson指数(D),其计算公式为:D=1-∑{[ni(ni-1)]/[N(N-1)]}。种群遗传均匀性指数采用Fager指数(E)来表示,计算公式为E=(D-Dmin)/(Dmax-Dmin),其中Dmin =(G-1)(2 N-G)/N(N-1),Dmax=(G-1)N/G(N-1)。式中N为取样个体总数,G为基因型(基株)总数,ni为第i基因型的个体数[27]。
使用GenAlEx 6.5软件[35]计算种群间遗传分化系数Fst,并进行分子方差分析(AMOVA,analysis of molecular variance analysis),计算种群内和种群间遗传方差分量。遗传方差分量和成对种群遗传分化系数的统计显著性均采用9 999次置换评价。
为了检测参试的蛇莓种群遗传结构,采用STRUCTURE[36]分析参试样本的遗传结构。将MCMC (markov chain monte carlo) 不作数迭代(length of burn-in period)设为1万次,不作数迭代后的MCMC也设为1万次,组群数(K)设定为2~33(参试种群数目),评价K值运行20次。依据每次测试过程中计算的后验概率lnP(D)值计算ΔK值,绘制ΔK曲线图。散点曲线最大值对应的K值视为最合理的组群数[37]。
使用NTSYS 2.1软件[38]对种群间Neis遗传多样性指数表2蛇莓SSR引物序列
引物序列(5′→3′)重复碱基片段大小
统计量距离进行非加权平均配组(unweighted pair group method with arithmetic-means,UPGMA)聚类分析,绘制树状聚类图。利用GenAlEx 6.5软件中的Distance计算33个蛇莓野生种群间地理距离,Mantel统计学分别检验分析种群间的地理距离和遗传距离相关性,并进行显著性检测(9 999次置换)。
2结果与分析
2.1种群遗传变异和克隆多样性
选用8对能扩增出稳定、清晰条带的SSR引物,对33个蛇莓种群353株个体进行PCR扩增,共扩增出94个等位基因,每个位点的等位基因数为7~23。蛇莓各种群的平均每个位点的等位基因数(A)为1.5~5,平均2.841;期待杂合度(He)为0.213~0.682,平均0.494;Neis基因多样性指数(h)为0.465~0.640,平均0.468(表3)。蛇莓种群平均遗传多样性较高(A=2.841,He=0.494,h=0.468),其中,北京小龙门种群(XLM)的遗传多样性最高(A=4.55,He=0.682,h=0.640),而江西九江种群(JJ)的遗传多样性最低(A=1.5,He=0.213,h=0.199,表3)。蛇莓物种水平的遗传多样性也较高(A=13.000,He=0.657,h=0.656)。
所有种群均为多克隆(基因型)种群,克隆(基因型)总数为223,各种群的基因型数为2~15,平均6.758;克隆大小为1.0~7.5,平均2.063;局部基因型比例为82.6%;广布基因型比例为0;平均基因型多样性指数Simpson指数为0.769(表3)。
2.2种群遗传结构
SRUCTURE分析结果表明,当K=4时,ΔK散点曲线呈稳定趋势,可推测出所有参试的353株个体最合理的组群数为4(图2)。图2中纵坐标Q值表示不同植株归属于不同组群的比例,黑色(组群Ⅰ)、深灰色(组群Ⅱ)、浅灰色(组群Ⅲ)和白色(组群Ⅳ)分别代表组群的趋向,每个个体在4种颜色中最长色条的颜色决定该个体所属的组群。组群Ⅰ包括FS、GZ2、HZ3、JJ和SCH1,组群Ⅱ包括LYW1、LYW2、LYW3、LYW4、SCH2、SCH3、TZ1、 TZ2、XA1、XA2和XLM,组群Ⅲ包括GZ1、HZ1、HZ2、JAX、JGS、JGSU、JS、NJ1、NJ2、NJ3、GL1、GL2和GL3,组群Ⅳ包括CQ1、CQ2、CQ3和GZ3。
基于种群间遗传分化系数Neis遗传一致度指数的UPGMA聚类结果(图3)表明,在遗传分化系数以0.62为阈值的情况下,33个种群可分为四大类,其中CQ1、CQ2和CQ3为第1个类群;GZ3为第2个类群;HZ3和JJ为第3个类群;其他27个种群为第4个类群。
AMOVA结果(表4)表明,蛇莓种群间遗传变异约占总变异的55%,种群内遗传变异占总变异的45%,种群间遗传分化显著(Fst=0.55,P=0.01)。成对种群间遗传分化系数在0.020~0.464之间,各种群间遗传分化达到显著水平(P< 0.05)。其中,GL2和JJ种群之间遗传分化系数最大(Fst=0.464),HZ1与NJ1种群之间遗传分化系数最小(Fst=0.020)。在地理距离水平上,北京房山(FS)与桂林3(GL3)种群之间距离最远,为1 660.158 km;南京种群1与南
京种群3距离最近,为0.57 km。Mantel相关性矩阵检验结果表明,种群间遗传距离Fst与地理距离呈正相关但不显著(r=0.052,P=0.150)。
3结论与讨论
3.1蛇莓的遗传变异和克隆多样性
很多研究结果表明,克隆植物具有一定的遗传变异水平,但不同克隆植物的遗传变异程度、基因型多样性有很大的差异[27,39]。本研究结果表明,蛇莓物种水平的遗传多样性(A=13.000,He=0.657,h=0.656,表3)比27种克隆植物的平均值(D=0.62)高,比100多种多年生木本植物的平均值(Na=1.76,He=0.148)高[40-41]。这可能是由于蛇莓为一种广布的草本植物,因此具有较高的物种水平遗传多样性。蛇莓种群水平的遗传多样性也较高(A=2.841,He=0.494,h=0.468),其中以北京小龙门种群(XLM)的遗传多样性最高(A=4.55,He=0.682,h=0.640),而江西九江种群(JJ)的遗传多样性最低(A=1.5,He=0.213,h=0.199)。这可能与不同生境中蛇莓种群的有性繁殖与无性繁殖的相对贡献度有关。
蛇莓种群的克隆多样性较高(G=6.758,N/G=2.063,D=0.769,E=0.769)。虽然一些克隆植物的克隆多样性很低,如水葫芦和空心莲子草等。其他一些研究也发现,有些克隆植物的克隆多样性非常高,如越橘(Vaccunium vitisidaea,D=0.84)和仙人掌科入侵植物Stenocereus eruca(N/G=125)等[42-43]。理论研究结果表明,因为克隆植物的生命期长,所以即使非常少量的幼苗更新也能使克隆种群维持较高的克隆多样性[28]。Bengtsso认为,克隆植物1个世代只要有3个不同的基因型就可以保持克隆植物的遗传变异水平[44]。计算机模拟结果显示,匍枝毛茛(Rannuulus repens)只要每一代有1个有性幼苗建立(约占总基株的0.5%),就足以维持15个基因型的稳定结构[28]。此外,不同蛇莓种群的基因型数量差异较大,如CQ1、GZ2和GL2等3个种群只有2个基因型组成,但其等位基因水平上的多样性却很高,3个种群的期望杂合度(He)分别是0.404、0.399、0.392,而期望杂合度(He)相对低的JJ种群(He=0.213)却有多个基因型(G=5,N/G=1600),这说明蛇莓能很好地维持种群的遗传多样性。对蛇莓有性繁殖特性的研究结果表明,蛇莓虽然能产生大量的种子,但种子的萌发率并不高,仅10%[31,45]。而从该研究的结果来看,较低的种子萌发率并没有影响蛇莓的遗传多样性。与其他克隆植物相比,有性繁殖对这些植物遗传多样性的维持起非常重要的作用[42,46-47]。所以,尽管蛇莓的种子萌发率低,但只要有很少的幼苗更新就能维持其种群的遗传多样性。
然而,在基因型较少的种群中,可能存在克隆繁殖占主导地位的现象。因为克隆生长产生的分株个体由母体的供养能够比较顺利地度过建立期,有利于基株对局部空间的快速占据,而克隆整合、克隆可塑性和克隆器官的存储能力等克隆生长习性还可以缓解资源的小尺度空间和时间异质性对基株适合度的影响[1]。克隆繁殖占优势意味着蛇莓对有性繁殖的资源分配较少[2],从而可以分配更多的能量进行克隆生长,从环境中获取更多资源,进而在群落中占领优势地位。此外,较少的有性繁殖意味着近交衰退的减少,而蛇莓本身具有较高的等位基因多样性,所以在某种程度上克隆繁殖有利于维持蛇莓种群的遗传多样性。蛇莓野外种群的有性繁殖和无性繁殖的相对格局还须要进一步研究。
3.2蛇莓种群的遗传结构
一般认为,当Fst<0.05时,种群间遗传分化较小;当 0.05
UPGMA聚类结果表明,地理距离较近的种群则归属于同一类群,同一省(市、区)的种群基本属于同一类群,但也有例外,一些地理距离较近的种群没有归为同一类群,如北京房山(FS)和北京小龙门(XLM),四川简阳市1(SCH1)与四川简阳市2(SCH3)和四川简阳市3(SCH2),以及杭州植物园2(HZ3)与杭州其他2个种群(HZ1和HZ2),另外贵州3个种群分别属于不同的类群。STRUCTURE分析结果也基本类似,大部分同省(市、区)种群都率先聚集在一起,然后再依地理距离由近到远依次聚在一起,但是仍存在一些同省(市、区)没有优先聚在一起的种群,其中GZ3、HZ3和JJ即为特殊的类群。比较STRUCTURE和UPGMA结果可知,33个蛇莓野生种群的聚类大体一致,均可以分为4个组群。说明蛇莓的遗传距离与地理距离相关,但也有其特殊的复杂性。
Mantel分析发现,蛇莓的遗传距离与地理距离的相关性不显著。这些均与蛇莓没有广布基因型,且局部基因型占比(GL=82.6%)很高有关。葛颂等在兼性克隆植物羊柴(Hedysarum laeve)中也发现了类似结果,其局部基因型比例为60%,广布基因型比例为3.2%。这与Ellstrand等的总结一致,即克隆植物的大部分基因型都仅存在于1个或很少的种群内[40]。这是因为克隆植物的遗传基础来源于建群者,并且在种群的发展过程中不断变化。克隆植物在不同的环境中受到不同的选择压力,环境异质性可以促进不同基因型的固定[53],从而占据不同的生境。尽管蛇莓在种群间存在一定的基因交流,但是该物种一般进行无性繁殖,所以基因型的局域性很强,而蛇莓在中国是一个广布种,所以其物种水平上的遗传多样性很高。
本研究结果表明,蛇莓在物种水平上的遗传多样性很高,基因型存在很强的局域性,且其遗传距离与地理距离不存在显著相关性。这些发现对研究蛇莓乃至克隆植物的进化潜能及生态特征具有重要意义。对于克隆植物野生种群的繁殖格局,遗传多样性对物种繁殖格局的影响,以及克隆植物中有性繁殖与无性繁殖之间权衡的科学问题仍须要开展相关的研究。
参考文献:
[1]董鸣,于飞海,陈玉福,等. 克隆植物生态学[M]. 北京:科学出版社,2011:67,71-74.
[2]张玉芬,张大勇. 克隆植物的无性与有性繁殖对策[J]. 植物生态学报,2006,30(1):174-183.
[3]王洪新,胡志昂. 植物的繁育系统、遗传结构和遗传多样性保护[J]. 生物多样性,1996,4(2):32-36.
[4]Escaravage N,Questiau S,Pornon A,et al. Clonal diversity in a Rhododendron ferrugineum L.(Ericaceae) population inferred from AFLP markers[J]. Molecular Ecology,1998,7(8):975-982.
[5]Setsuko S,Ishida K,Tomaru N. Size distribution and genetic structure in relation to clonal growth within a population of Magnolia tomentosa Thunb. (Magnoliaceae)[J]. Molecular Ecology,2004,13(9):2645-2653.
[6]Dong M. Clonal growth in plants in relation to resource heterogeneity:foraging behavior[J]. Acta Botanica Sinica,1996,38(10):828-835.
[7]于飞海. 克隆植物对异质性环境的生态适应对策[D]. 北京:中国科学院研究生院/植物研究所,2002:2-4.
[8]夏立群,李建强,李伟. 论克隆植物的遗传多样性[J]. 植物学通报,2002,19(4):425-431,418.
[9]Yu F H,Dong M,Krusi B. Clonal integration helps Psammochloa villosa survive sand burial in an inland dune[J]. New Phytologist,2004,162(3):697-704.
[10]Yu F H,Wang N,He W M,et al. Adaptation of rhizome connections in drylands:increasing tolerance of clones to wind erosion[J]. Annals of Botany,2008,102(4):571-577.
[11]Yu F H,Wang N,He W M,et al. Effects of clonal integration on species composition and biomass of sand dune communities[J]. Journal of Arid Environments,2010,74(6):632-637.
[12]Luo F L,Chen Y,Huang L,et al. Shifting effects of physiological integration on performance of a clonal plant during submergence and de-submergence[J]. Annals of Botany,2014,113(7):1265-1274.
[13]Zhang Y F,Zhang D Y. Asexual and sexual reproductive strategies in clonal plants[J]. Frontiers of Biology in China,2007,2(3):256-262.
[14]Li J,Ye W H. Genetic diversity of alligator weed ecotypes is not the reason for their different responses to biological control[J]. Aquatic Botany,2006,85(2):155-158.
[15]Zhang Y Y,Zhang D Y,Barrett S C. Genetic uniformity characterizes the invasive spread of water hyacinth (Eichhornia crassipes),a clonal aquatic plant[J]. Molecular Ecology,2010,19(9):1774-1786.
[16]Wilcock C C,Jennings S B. Partner limitation and restoration of sexual reproduction in the clonal dwarf shrub Linnea borealis L.(Caprifoliaceae)[J]. Protoplasma,1999,208:76-86.
[17]Dietz H,Steinlein T. Ecological aspects of clonal growth in plants[J]. Ecology,2001,62:511-529.
[18]项卫东,张亚梅. 外来入侵种空心莲子草的RAPD遗传多样性分析[J]. 南京林业大学学报:自然科学版,2004,28(6):35-38.
[19]Gitzendanner M A,Weekley C W,Germain-Aubrey C C,et al. Microsatellite evidence for high clonality and limited genetic diversity in Ziziphus celata (Rhamnaceae),an endangered,self-incompatible shrub endemic to the Lake Wales Ridge,Florida,USA[J]. Conservation Genetics,2012,13(1):223-234.
[20]Ren M X,Zhang Q G,Zhang D Y. Random amplified polymorphic DNA markers reveal low genetic variation and a single dominant genotype in Eichhornia crassipes populations throughout China[J]. Weed Research,2005,45(3):236-244.
[21]Li W G,Wang B R,Wang J B. Lack of genetic variation of an invasive clonal plant Eichhornia crassipes in China revealed by RAPD and ISSR markers[J]. Aquatic Botany,2006,84(2):176-180.
[22]Ye W,Li J,Cao H,et al. Genetic uniformity of Alternanthera philoxeroidesin South China[J]. Weed Research,2003,43(4):297-302.
[23]Kavitha P G,Thomas G. Population genetic structure of the clonal plant Zingiber zerumbet (L.) Smith (Zingiberaceae),a wild relative of cultivated ginger,and its response to Pythium aphanidermatum[J]. Euphytica,2008,160(1):89-100.
[24]Lloyd M W,Burnett R,Engelhardt K M,et al. The structure of population genetic diversity in Vallisneria americana in the Chesapeake Bay:implications for restoration[J]. Conservation Genetics,2011,12(5):1269-1285.
[25]Vaughan S P,Cottrell J E,Moodley D J,et al. Clonal structure and recruitment in British wild cherry (Prunus avium L.)[J]. Forest Ecology and Management,2007,242(2/3):419-430.
[26]de Rogatis A,Ferrazzini D,Ducci F,et al. Genetic variation in Italian wild cherry (Prunus avium L.) as characterized by nSSR markers[J]. Forestry,2013,86(3):391-400.
[27]葛颂,王可青,董鸣. 毛乌素沙地根茎灌木羊柴的遗传多样性和克隆结构[J]. 植物学报,1999,41(3):75-80.
[28]Watkinson A R,Powell J C. Seedling recruitment and the maintenance of clonal diversity in plant-populations-a computer-simulation of Ranunculus-repens[J]. Journal of Ecology,1993,81(4):707-717.
[29]Wu Z Y,Raven P H,Hong D Y. Flora of China[M]. St. Louis:Science Press(Beijing) & Missouri Botanical Garden,2004.
[30]Naruhashi N,Sugimoto M. The floral biology of Duchesnea(Rosaceae)[J]. Plant Species Biology,1996,11(2/3):173-184.
[31]李钧敏,金则新. 匍匐茎草本植物蛇莓小尺度克隆结构[J]. 生态学报,2009,29(7):3540-3548.
[32]罗学刚,董鸣. 匍匐茎草本蛇莓克隆构型对土壤养分的可塑性反应[J]. 生态学报,2001,21(12):1957-1963.
[33]罗学刚,董鸣. 匍匐茎草本蛇莓克隆构型对不同海拔的可塑性反应[J]. 应用生态学报,2002,13(4):399-402.
[34]Yeh F,Yang R,Boyle T J,et al. Popgen 32,microsoftware windows based freeware for population genetic analysis[M]. Edmonton:Molecular Biology and Biotechnology Center,2002.
[35]Peakall R,Smouse P E. GenAlEx 6.5:genetic analysis in Excel. Population genetic software for teaching and research:an update[J]. Bioinformatics,2012,28(19):2537-2539.
[36]Pritchard J K,Stephens M,Donnelly P. Inference of population structure using multilocus genotype data[J]. Genetics,2000,155(2):945-959.
[37]Evanno G,Regnaut S,Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study[J]. Molecular Ecology,2005,14(8):2611-2620.
[38]Rohlf F J. NTSYS-pc 2.1:numerical taxonomy and multivariate analysis system[M]. Exeter Software,Setauket,NY,1997.
[39]Hamrick J L,Godt M J W,Brown A H D,et al. Allozyme diversity in plant species[C]. Plant population genetics,breeding,and genetic resources,1990:43-63.
[40]Ellstrand N C,Roose M L. Patterns of genotypic diversity in clonal plant species[J]. American Journal of Botany,1987,74(1):123-131.
[41]Hamrick J L,Godt M J,Sherman-Broyles S L. Factors influencing levels of genetic diversity in woody plant species[J]. New Forests,1992,6(1/2/3/4):95-124.
[42]Kreher S A,Foré S A,Collins B S. Genetic variation within and among patches of the clonal species,Vaccinium stamineum L.[J]. Molecular Ecology,2000,9(9):1247-1252.
[43]Clark-Tapia R,Alfonso-Corrado C,Eguiarte L E,et al. Clonal diversity and distribution in Stenocereus eruca (Cactaceae),a narrow endemic cactus of the Sonoran Desert[J]. American Journal of Botany,2005,92(2):272-278.
[44]Bengtsson B O. Genetic variation in organisms with sexual and asexual reproduction[J]. Journal of Evolutionary Biology,2003,16(2):189-199.
[45]陈明. 蛇莓观赏性与繁殖习性研究[J]. 林业实用技术,2008(2):42-43.
[46]Clark-Tapia R,Mandujano M C,Valverde T,et al. How important is clonal recruitment for population maintenance in rare plant species?The case of the narrow endemic cactus,Stenocereus eruca,in Baja California,México[J]. Biological Conservation,2005,124(1):123-132.
[47]Li J,Dong M. Fine-scale clonal structure and diversity of invasive plant Mikania micrantha HBK and its plant parasite Cuscuta campestris Yunker[J]. Biological Invasions,2009,11(3):687-695.
[48]Wright S. The interpretation of population-structure by F-statistics with special regard to systems of mating[J]. Evolution,1965,19(3):395-420.
[49]Nybom H,Bartish I V. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants[J]. Perspectives in Plant Ecology,Evolution and Systematics,2000,3(2):93-114.
[50]Nybom H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants[J]. Molecular Ecology,2004,13(5):1143-1155.
[51]Glémin S,Bazin E,Charlesworth D. Impact of mating systems on patterns of sequence polymorphism in flowering plants[J]. Proceedings of Biological Sciences,2006,273(1604):3011-3019.
[52]Heleno R H,Ross G,Everard A,et al. The role of avian ‘seed predators as seed dispersers[J]. IBIS,2011,153(1):199-203.
[53]沈栋伟,李媛媛,陈小勇. 植物克隆多样性与生态系统功能[J]. 植物生态学报,2007,31(4):552-560.彭燕琼,吉根宝,李国平,等. 观光果园的建设与管理[J]. 江苏农业科学,2016,44(2):221-223,251.
猜你喜欢
长江蔬菜·学术版(2016年12期)2017-01-12
农家科技下旬刊(2016年9期)2016-12-15
河北渔业(2016年9期)2016-11-05
现代园艺(2016年3期)2016-03-24
