
1982—2011年江苏大麦育成品种亲本分析
2016-06-14 23:47张英虎沈会权臧慧栾海业乔海龙陶
江苏农业科学 2016年4期
张英虎+沈会权+臧慧+栾海业+乔海龙+陶红+陈健+陈和
摘要:掌握育成品种的遗传基础,可揭示品种演变特点,总结亲本选配经验,并指导育种实践。本研究对1982—2011年江苏省审定的42个大麦品种的育种途径和系谱进行分析,结果表明,杂交方法是大麦育种的主要方法,有 71.42% 的品种用杂交方法选育。育成品种共来源于67个细胞核祖先亲本和34个细胞质祖先亲本,来源于江苏、日本两地品种的细胞核遗传贡献分别为36.90%、45.98%,表明江苏地区育成品种祖先亲本主要来源于这两地。来源于日本关东二条3号、新田二条1号累积细胞核贡献大、衍生品种多,为核心祖先亲本。分析了由这2个品种衍生育成品种的系谱关系,结果表明,江苏大麦育成品种遗传基础较为狭隘,需要加强新种质的研究利用,以扩大品种的遗传基础。
关键词:江苏;大麦;育成品种;亲本;系谱
中图分类号:S512.303
文献标志码: A
文章编号:1002-1302(2016)04-0141-04
育成品种是经过人工选择和系统鉴定的,具有优良基因型,是进一步进行品种改良的重要亲本来源[1]。掌握育成品种的遗传基础,可揭示品种演变特点,总结亲本选配经验,并指导进一步育种实践[2]。
在大豆上,熊冬金等对中国1923—2005年间的1 300个育成品种亲本进行分析,发现所有品种来源于670个细胞核和344个细胞质终端祖先亲本,并归纳出1986—2005年间最重要的54个祖先亲本[3]。在玉米上,孙琦等分析了2001—2012年国审普通玉米杂交种及其双亲自交系的来源,发现亲本自交系主要来自黄早四的衍生系、丹340及其衍生系、478衍生系、78599选系及其衍生系、Mo17改良系[4]。在水稻上,刘化龙等分析了黑龙江审定的260个水稻品种的骨干亲本以及20世纪90年代后育成品种的系谱,发现20世纪90年代前育成品种的骨干亲本是石狩白毛、农林11号、虾夷;20世纪90年代后育成品种的骨干亲本是藤系138、上育397、富士光[5]。在小麦上,杨春玲等对河南省近20年小麦品种系谱分析表明,河南省小麦品种选育中起骨干作用的亲本基本为三大系:分别为地方品种辉县红及其衍生系、意大利“St”系统及其衍生系、意大利阿夫及其衍生系[6]。
大麦是世界上最主要、最古老的栽培作物之一,播种面积和总产量仅次于玉米、小麦、水稻,居谷类作物第4位[7]。江苏是中国三大大麦主产区之一,也是种植面积波动最小、面积最大的地区[8]。梁长东等回顾了江苏大麦种植历史,介绍了从“七五”到“十五”末,江苏共育成的30个大麦品种,建议啤酒大麦新品种的选育应将酿造品质放在第一位[9]。赵理清等分析了1949—1987年育成的大麦品种的系谱,发现在长江中下游冬大麦区的核心亲本为早熟3号[10]。
本研究以江苏省1982—2011年育成的42个大麦品种为研究材料,追溯其系谱,分析祖先亲本的地理来源和遗传贡献,为有效利用大麦种质资源,拓展大麦品种遗传基础,合理选配亲本提供理论依据。
1 材料与方法
1.1 材料
追溯1982—2011年江苏育成的42个大麦品种的系谱,系谱资料主要来源于江苏种业信息网(http://www.jsseed.cn)、《江苏省稻麦品种志》[11]以及各育种单位有关专家及相关刊物上发表的选育报告。
1.2 方法
根据42个大麦育成品种的系谱资料列出其祖先亲本,计算出每个育成品种的祖先亲本细胞核遗传贡献值。凡由祖先亲本通过自然变异选择法育成的品种,其祖先亲本的细胞核遗传贡献值为1;由杂交育成的品种,其双亲的核遗传贡献均为0.5,每个亲本再按均等分割方法上推其双亲,直至终极的祖先亲本,这样每一育成品种的各祖先亲本核遗传贡献值总和应等于1;凡通过诱变育成的品种,因突变成分相对较小,其祖先亲本核遗传贡献值的计算与自然变异选择育成品种的方法相同;凡由杂交与诱变相结合方法育成的,其祖先亲本核遗传贡献值的计算与杂交育种相同。亲本贡献值的计算参照盖钧镒等的方法[12]。
2 结果与分析
2.1 1982—2011年江苏省大麦育成品种
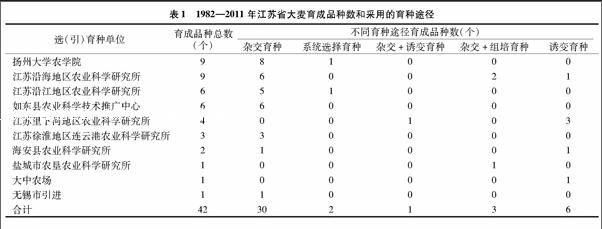
江苏省1982—2011年共育成42个大麦品种(表1),扬州大学农学院、江苏沿海地区农业科学研究所育成的品种数最多,均为9个;江苏沿江地区农业科学研究所、如东县农业科学技术推广中心、江苏里下河地区农业科学研究所、江苏徐淮地区连云港农业科学研究所、海安县农业科学研究所分别育成了6、6、4、3、2个大麦品种,盐城市农垦农业科学研究所、大中农场和无锡市选(引)大麦品种均为1个。江苏大麦育种途径以杂交育种为主,共育成30个品种,占71.42%;以诱变育种育成6个品种,占14.28%;杂交+组培育种育成3个品种,占7.14%;系统选择育种育成2个品种,占 4.76%;杂交+诱变育种育成1个品种,占2.38%。江苏里下河地区农业科学研究所大麦育种方法主要采用诱变育种,在该所育成的4个大麦品种中,3个采用诱变育种,1个采用杂交+诱变育种的方法育成。
2.2 1982—2011年江苏大麦育成品种祖先亲本的地理来源及其细胞核遗传贡献
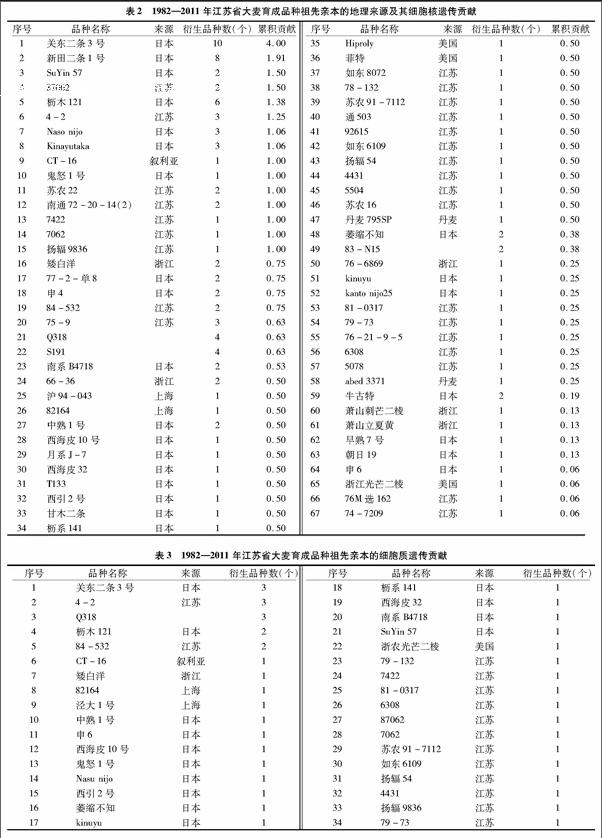
42个育成品种来源于67个祖先亲本,其中国内亲本33个,国外亲本31个,来源不清楚的有3个(表2)。国内亲本来源于江苏、浙江、上海3省(市),江苏为26个,占 38.24%;浙江为5个,占7.35%;上海为3个,占4.41%。细胞核遗传贡献率江苏为15.50,占36.90%;浙江为1.75,占4.17%;上海为1.50,占3.57%;国外亲本来源于日本、美国、丹麦、叙利亚等地,日本为25个,占36.76%;美国为3个,占4.41%;丹麦为2个,占2.94%;叙利亚为1个,占1.47%。细胞核遗传贡献率日本为19.31,占45.98%;美国为1.06,占2.52%;丹麦为0.75,占1.79%;叙利亚为1.00,占2.38%。表明国内祖先亲本主要来自江苏,国外祖先亲本主要来自日本。在祖先亲本中,细胞核贡献最大的2个祖先亲本分别是来自日本的关东二条3号(4.00)、新田二条1号(1.91),分别衍生了10个和8个育成品种。
2.3 1982—2011年江苏大麦育成品种祖先亲本细胞质遗传贡献
42个大麦育成品种细胞质祖先亲本有34个(表3),细胞质来源主要有关东二条3号,衍生品种3个;4-2,衍生品种3个;Q318,衍生品种3个;枥木121,衍生品种2个;84-532,衍生品种2个。同时发现,江苏大麦育成品种中,细胞质遗传贡献主要来源于江苏和日本的材料,来源于江苏的品种衍生了17个品种,占细胞质遗传贡献的40.48%,来源于日本的品种衍生了17个品种,占细胞质遗传贡献的40.48%。
2.4 江苏大麦主要祖先亲本关东二条3号、新田二条1号衍生大麦育成品种系谱情况
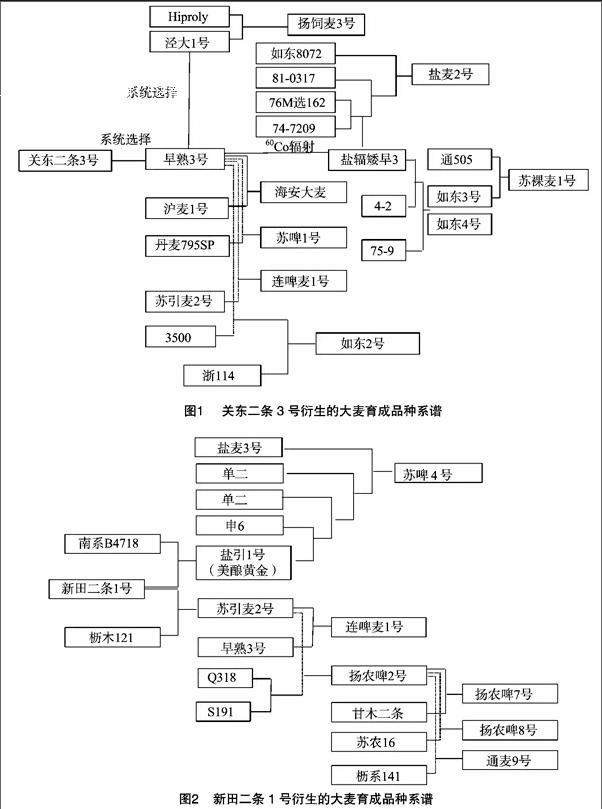
通过对42份江苏大麦育成品种祖先亲本细胞核遗传贡献分析,发现关东二条3号、新田二条1号细胞核遗传贡献最高。利用关东二条3号、新田二条1号在1982—2011年江苏地区衍生的大麦育成品种情况见图1、图2。
从图1可以看出,关东二条3号通过系统选择培育出了早熟3号,利用早熟3号作为亲本分别培育出了盐辐矮早3、海安大麦、苏啤1号、连啤麦1号和如东2号,利用盐辐矮早3为亲本培育出了盐麦2号、如东3号和如东4号,利用如东3号为亲本培育出了苏裸麦1号。利用早熟3号衍生的泾大1号为亲本培育出了扬饲麦3号。
从图2可以看出,利用新田二条1号作为亲本培育出盐引1号、苏引麦2号,利用盐引1号作为亲本培育出苏啤4号,利用苏引麦2号作为亲本培育出连啤麦1号和扬农啤2号,利用扬农啤2号作为亲本培育出扬农啤7号、扬农啤8号和通麦9号。
3 讨论
1982—2011年江苏大麦育成品种中71.42%是通过杂交育种育成,表明杂交育种仍是江苏大麦育种的主要方式,42个育成品种中还没有利用分子标记辅助选择培育出来的大麦品种。分子育种技术被认为是21世纪作物改良的基础技术[13],正逐渐成为作物育种的主流[14]。目前江苏还没有通过分子育种技术培育的品种,但江苏沿海地区农业科学研究所已收集整理报道的大麦耐盐QTL[15],为分子育种研究提供了基础信息。
江苏大麦育成品种的细胞核遗传贡献和细胞质遗传贡献主要来源于江苏地区和日本,表明江苏大麦育成品种的遗传基础较为狭隘。作物遗传基础的局限性会导致遗传脆弱性,其潜在的危害已引起各国育种间的关注[16],美国大豆育种的成功经验就是较好地利用了国外异生态区的种质[17]。起源中心的种质多样性较高,研究表明,大麦基因组有两个起源地,分别是伊拉克、叙利亚、黎巴嫩和以色列等国所处的中东地区,中国西藏及周边地区[18],但现阶段江苏地区育成品种对这两个起源地的种质利用较少,下一步将加强对起源地种质的引入、研究和利用。
赵理清等分析了1949—1987年育成的大麦品种的系谱,发现在长江中下游冬大麦区的核心亲本为早熟3号,早熟3号是由关东二条3号系统选育而来[10]。本研究对1982—2011年江苏育成品种的核心亲本分析结果与赵理清等的研究结果一致,同时进一步发现新田二条1号也是江苏育成品种的核心亲本之一。在其他作物上也发现少数核心品种遗传贡献和衍生品种数较高[19]。在今后的研究中拟进一步研究关东二条3号、新田二条1号的遗传结构,同时分析2个核心品种衍生品种的基因型和表型的变化,从表型和分子水平了解大麦品种的发展趋势,为大麦育种提供理论支持。
参考文献:
[1]Gai J Y,Chen L,Zhang Y H,et al. Genome-wide genetic dissection of germplasm resources and implications for breeding by design in soybean[J]. Breeding Science,2012,61(5):495-510.
[2]刘章雄,李卫东,孙 石,等. 1983—2010年北京大豆育成品种的亲本地理来源及其遗传贡献[J]. 大豆科学,2013,32(1):1-7.
[3]熊冬金,赵团结,盖钧镒. 中国大豆育成品种亲本分析[J]. 中国农业科学,2008,41(9):2589-2598.
[4]孙 琦,李文才,张发军,等. 2001—2012年国审玉米品种数量及骨干自交系的应用分析[J]. 玉米科学,2014,22(4):39-43.
[5]刘化龙,王敬国,赵宏伟,等. 黑龙江水稻育种骨干亲本及系谱分析[J]. 东北农业大学学报,2011,42(4):18-21.
[6]杨春玲,侯军红,宋志均,等. 河南省主要小麦品种系谱研究及核心种质利用[J]. 山东农业科学,2009(1):27-31.
[7]栾海业,陈 和,陈 健,等. 大麦耐湿性研究进展[J]. 江苏农业科学,2012,40(12):116-117.
[8]陈 和,许如根,王龙俊,等. 江苏啤酒大麦品种发展及其品质定位[J]. 江苏农业科学,2011,39(4):1-4.
[9]梁长东,徐大勇,李荣花. 江苏大麦育种工作的回顾与建议[J]. 大麦科学,2005(3):1-3.
[10]赵理清,陆美琴. 我国大麦品种的选育[J]. 浙江农业科学,1992(6):253-258.
[11]王才林. 江苏省稻麦品种志[M]. 北京:中国农业科学技术出版社,2009.
[12]崔章林,盖钧镒,Carter T J,等. 中国大豆育成品种及其系谱分析(1923—1995) [M]. 北京:中国农业出版社,1998.
[13]Moose S P,Mumm R H. Molecular plant breeding as the foundation for 21st century crop improvement[J]. Plant Physiology,2008,147(3):969-977.
[14]黎 裕,王建康,邱丽娟,等. 中国作物分子育种现状与发展前景[J]. 作物学报,2010,36(9):1425-1430.
[15]沈会权,张英虎,乔海龙,等. 基于BinMap的大麦耐盐QTL整合[J]. 江苏农业学报,2014,30(4):918-920.
[16]Singh R J,Hymowitz T. Soybean genetic resources and crop improvement[J]. Genome,1999,42(4):605-616.
[17]Sneller C H. Impact of transgenic genotypes and subdivision on diversity within elite North American soybean germplasm[J]. Crop Science,2003,43(1):409-414.
[18]Dai F,Chen Z H,Wang X L,et al. Transcriptome profiling reveals mosaic genomic origins of modern cultivated barley[J]. Proceedings of the National Academy of Sciences of the United States of America,2014,111(37):13403-13408.
[19]熊冬金,王吴彬,赵团结,等. 中国大豆育成品种10个重要家族的遗传相似性和特异性[J]. 作物学报,2014,40(6):951-964.吕艳东,孙立明,周 健,等. 膜下滴灌旱种对寒地水稻产量构成因素及产量的影响[J]. 江苏农业科学,2016,44(4):145-148.
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
环境卫生工程(2021年4期)2021-10-13
华北电力大学学报(社会科学版)(2021年2期)2021-07-21
长江蔬菜(2021年12期)2021-04-04
疯狂英语·新读写(2021年2期)2021-02-25
中国果业信息(2019年11期)2019-01-05
图书馆理论与实践(2018年2期)2018-01-28
中国奶牛(2017年2期)2017-03-22
求学·理科版(2017年1期)2017-03-02
西南农业学报(2016年5期)2016-05-17
