
松墨天牛核糖体蛋白(RPS15A)cDNA的克隆及农药胁迫对其表达的影响
2016-06-15 03:14罗淋淋蔡紫玲
西北农林科技大学学报(自然科学版) 2016年2期
罗淋淋,蔡紫玲,林 同
(华南农业大学 林学与风景园林学院,广东 广州 510642)
松墨天牛核糖体蛋白(RPS15A)cDNA的克隆及农药胁迫对其表达的影响
罗淋淋,蔡紫玲,林同
(华南农业大学 林学与风景园林学院,广东 广州 510642)
[摘要]【目的】 研究不同类型农药胁迫下松墨天牛核糖体蛋白基因表达的变化,为靶标防治松树重要害虫松墨天牛提供理论依据和参考。【方法】 从构建的松墨天牛 cDNA 文库中获得表达序列标签(EST),用 Blast 程序从GenBank数据库中查找同源序列,克隆得到松墨天牛的一个核糖体蛋白基因;用实时荧光定量 PCR (RT-qPCR) 检测 11 种农药胁迫下该基因的表达。【结果】 从松墨天牛 cDNA 文库中获得1条 EST 序列与昆虫核糖体蛋白 S15A (RPS15A)同源的基因,命名为MaRPS15A。MaRPS15A cDNA长504 bp,含有 5′、 3′ 非编码区和长 393 bp 的开放阅读框,编码 130个氨基酸,预测的蛋白质分子质量为 14.75 ku,理论等电点为 9.95。MaRPS15A与赤拟谷盗(Tribolium castaneum)核糖体蛋白RPS15A的相似性为98%,与其他昆虫核糖体蛋白RPS15A的相似性大于88%。除溴氰菊酯外,其他农药胁迫导致MaRPS15A相对表达量下降。【结论】 成功克隆了松墨天牛核糖体蛋白基因MaRPS15A (GenBank登录号:KJ930032)。大多数农药抑制了MaRPS15A的表达,表明松墨天牛受到了不利影响。
[关键词]松墨天牛;核糖体蛋白;RPS15A;实时荧光定量 PCR (RT-qPCR);农药胁迫
核糖体(Ribosome)是细胞内负责蛋白质合成的重要细胞器。在真核生物中,核糖体由一个 60S 大亚基和一个 40S 小亚基组成。其中,大亚基约包含 49 种蛋白质,此外还有28S rRNA、5S rRNA和5.8S rRNA 3种rRNA;小亚基约包含 34 种蛋白质和一种 18S rRNA[1-2]。核糖体蛋白 (Ribosomal protein,RP) 和核糖体研究一直是遗传学研究的重要领域。RPS15A 是 40S 小亚基的组成成分之一,属于核糖体蛋白 S8P 家族,它位于细胞质中,是高度保守的核糖体蛋白。松墨天牛(MonochamusalternatusHope)又名松褐天牛、松天牛,是我国松树的重要蛀干害虫,也是毁灭性病害松材线虫病(Bursaphelenchusxylophilus) 的主要媒介昆虫。本研究从松墨天牛 cDNA 文库中克隆到了松墨天牛的一个 RP 基因,并通过实时荧光定量PCR(RT-qPCR)技术,分析农药对松墨天牛核糖体蛋白基因表达的影响,旨在为松墨天牛的防治提供理论依据。
1材料与方法
1.1材料
松墨天牛幼虫由广东省林业科学研究院惠赠,在人工气候箱设置温度为 25 ℃、相对湿度为 75%、黑暗条件下,用人工饲料饲喂至成虫[3]。
Total RNA KitⅡ总RNA 提取试剂盒,购自 OMEGAS 生物公司;反转录试剂盒PrimeScript®RT Reagent Kit with gDNA Eraser、SYBR染料法实时荧光定量试剂盒SYBR®Premix Ex TaqTM,均购自TaKaRa生物有限公司。
1.2基因克隆和 RT-qPCR引物设计
从松墨天牛幼虫 cDNA 文库[4]中随机挑选阳性克隆测序,获得表达序列标签(Expressed sequence tags,ESTs),通过 NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi) 的 Blast程序搜索获取核糖体蛋白基因。根据获得的目的基因序列使用 Premier 5.0 软件设计引物,上游引物是:5′-GGGAAGATCGTTGTGAATTTG-3′,下游引物是: 5′-GGATTTTGCCTCCCAGATGT-3′,产物长度202 bp;内参基因上游引物是:5′-CTCTGCTATGTAGCCCTTGACTT-3′,下游引物是:5′-GGAGTTGTAGGTGGTTTCGTG-3′。引物均由上海生物工程技术服务有限公司合成。
1.3农药处理
用7种化学农药和4种生物农药,分别作用于松墨天牛成虫,农药规格、稀释倍数和处理方法见表1。
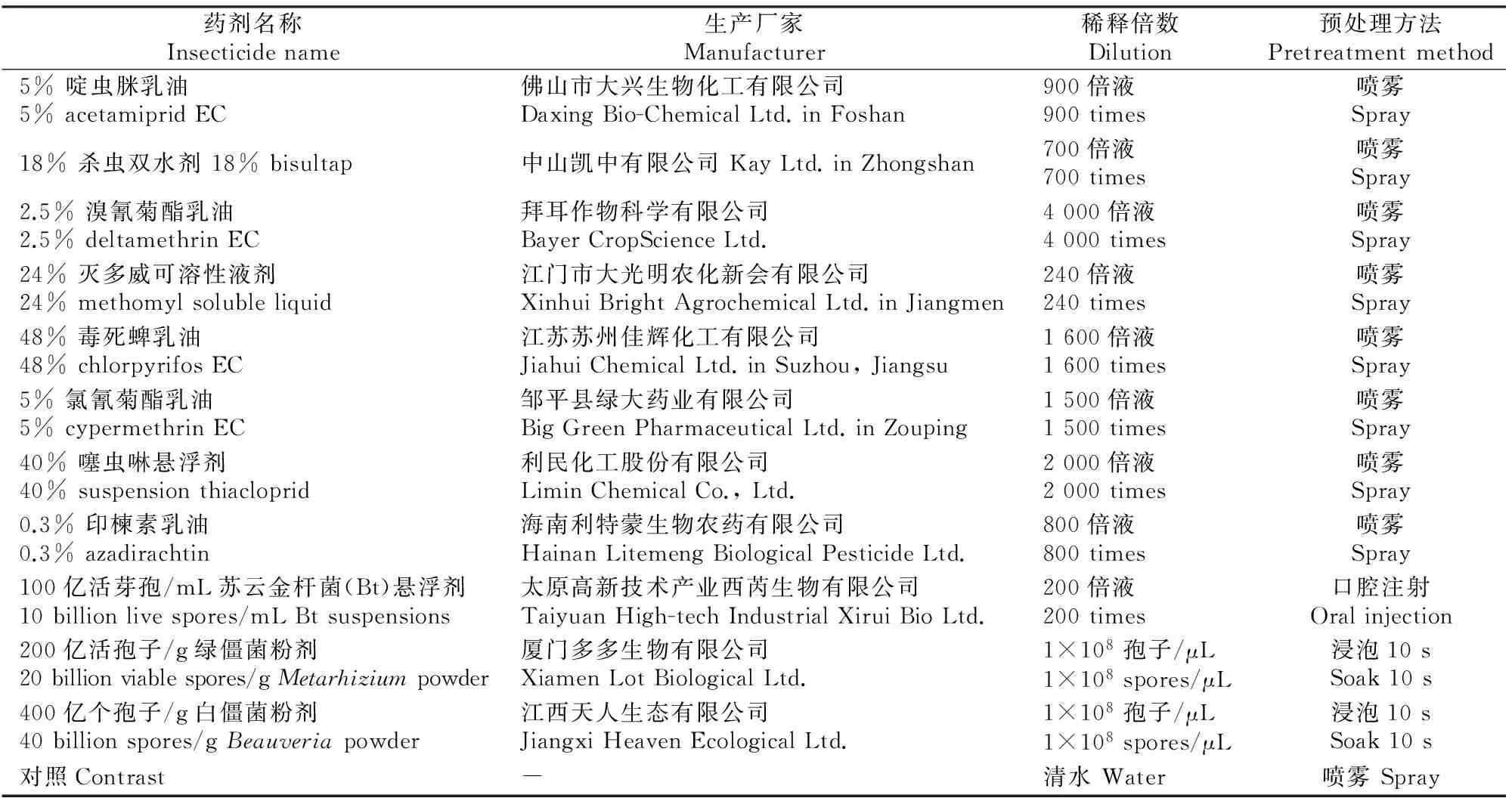
表 1 供试农药规格及稀释倍数
每种农药处理设3个重复,每个重复5头试虫,每个处理试虫受药12 h后,用液氮速冻,于-80 ℃ 保存备用。
1.4RT-qPCR检测
以目标基因在对照组 (未做处理的试虫) 的表达量为标准参量,以微管蛋白 (β-actin) 基因(GenBank登录号:JZ143740) 为内参基因,无菌超纯水为阴性对照,用RT-qPCR技术检测目标基因在11种农药胁迫后的相对表达量。RT-qPCR反应程序为:95 ℃预变性5 min;95 ℃ 10 s,60 ℃ 20 s,进行40个循环,cDNA样品设置3个重复,反应结束后采集目标基因和内参基因的CT平均值,利用 2-ΔΔCT相对定量法计算相对表达量[5]。
1.5克隆基因序列分析及其编码蛋白性质预测
用NCBI在线工具ORF finder (http://www.ncbi.nlm.nib.gov/gorf/grof.html)进行DNA序列开放阅读框(ORF)查找;用ExPASyProtParam工具 (http://www.expasy.org/) 进行蛋白质等电点、分子质量、磷酸化位点、信号肽预测等分析;用DNAMAN软件对氨基酸序列进行相似性比对;用Clustal X和MEGA 4.0软件构建系统发育树 (NJ 法)。
1.6数据处理与分析
采用Excel 2003和DPS-6.5软件的Duncan’s单因素方差分析法对RT-qPCR数据进行统计分析[6]。
2结果与分析
2.1松墨天牛MaRPS15A基因及序列分析
对松墨天牛幼虫cDNA 文库中的阳性克隆随机测序获得 EST序列,通过 NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi)中的 Blast 程序比对,发现在这些 EST 序列中,有1 条长度为 504 bp 的序列,与GenBank数据库中的昆虫 40S 核糖体蛋白 S15A 相似,将该基因命名为MaRPS15A,GenBank登录号为KJ930032。MaRPS15A基因从第 81-473 位核苷酸为开放阅读框(ORF)(图1),长393 bp,编码 130 个氨基酸,5′非编码区(5′-UTR) 长度为 80 bp,3′非编码区 (3′-UTR) 长度为31 bp,在3′-UTR有典型的加尾信号aataaa,这是 mRNA 前体在翻译后剪切所需要的。

图 1松墨天牛MaRPS15A基因核苷酸及其推导的氨基酸序列起始密码子atg用波浪线表示,丝氨酸和酪氨酸磷酸化修饰位点分别以圆形和下划线标注,N-糖基化位点用三角形表示,终止密码子用星号标注,加尾信号aataaa加框标注
Fig.1Nucleotides and deduced amino acid sequence ofMaRPS15AfromMonochamusalternatusThe translation start codon is indicated with wavy line,the serine sites are marked with circles and the tyrosine phosphorylation site is underlined,the glycosylation site is marked with triangle,the stop codon is marked with asterisk,and the polyadenylation signal is in rectangular
由ORF 核苷酸序列推测得到的 MaRPS15A 蛋白的分子式为C662H1 070N190O177S7,分子质量为 14.75 ku,理论等电点为9.95,为碱性蛋白。疏水性分析显示,平均亲水性结果为-0.090,为两性蛋白,MaRPS15A在29-48位氨基酸含有最强的疏水性区域;跨膜位点分析显示,MaRPS15A 在29-48 位氨基酸之间存在 1 个从内向外的跨膜位点,与该蛋白质的疏水性区域分析结果一致;MaRPS15A蛋白性质分析显示,不稳定指数为 35.22,属于稳定蛋白;MaRPS15A包含 2 个丝氨酸磷酸化位点、1 个酪氨酸磷酸化位点和1 个 N-糖基化位点。
2.2松墨天牛MaRPS15A相似性比对及系统发育树构建
用DNAMAN 软件对MaRPS15A氨基酸序列的相似性比对的结果见图2。

图 2松墨天牛MaRPS15与其他19种昆虫RPS15A氨基酸序列的比对
图中不同颜色阴影部分强调序列的相似性,黑色.相似性=100%;灰色.相似性 75%;白色.相似性 50%
Fig.2Amino acid sequence alignment of MaRPS15 and RPS15A fromMonochamusalternatusand other 19 of insects Different colors in the shadow highlight homology level,Black.Homology level=100%;Gray.Homology level=75%;White.Homology level=50%
图2表明,MaRPS15A 与来源于GenBank数据库的19 种昆虫核糖体蛋白 RPS15A 相似性达到88%~98%,其中,与赤拟谷盗(Triboliumcastaneum)、栎实象鼻虫 (Curculioglandium) 和白杨叶甲(Chrysomelatremulae) 等的相似性为98%,与黑腹果蝇(Drosophilamelanogaster) 和家蚕 (Bombyxmori) 相似性分别为 95% 和 94%。用以上相同的基因序列构建的系统发育树显示,所有的昆虫 RPS15A 起源于一个共同的祖先蛋白,首先进化出 2 个原始分支,其中节肢动物门甲壳纲无甲目卤虫(Artemiafranciscana) 单独一个进化分支;其他都属于昆虫纲,聚合成另一个独立的进化分支。其中松墨天牛与栎实象鼻虫在同一分支下,与白杨叶甲和赤拟谷盗亲缘关系比较近,与蚁类(佛罗里达弓背蚁(Camponotusfloridanus)、切叶蚁 (Acromyrmexechinatior)、印度跳蚁 (Harpegnathossaltator)) 和蜂类(丽蝇蛹集金小蜂(Nasoniavitripennis)、茶足柄瘤蚜茧蜂 (Lysiphlebustestaceipes)) 亲缘关系比较远(图3)。
2.3农药胁迫下松墨天牛MaRPS15A的表达
利用RT-qPCR分析了松墨天牛成虫MaRPS15A基因在11种农药胁迫下的相对表达量,结果见图4。由图4可见,以未处理组成虫MaRPS15A基因的表达量为对照,杀虫剂处理组的结果为:溴氰菊酯处理组MaRPS15A表达上调,相对表达量为对照组的1.101倍,其余10个处理组MaRPS15A表达均下调,杀虫双、毒死蜱、氯氰菊酯、啶虫脒、灭多威、噻虫啉、印楝素、绿僵菌、白僵菌、Bt胁迫下MaRPS15A的相对表达量分别为对照组的30.39%,40.68%,66.65%,19.92%,14.50%,92.90%,54.79%,16.85%,52.00% 和42.19%。各处理间的相对表达量差异显著(P<0.5)。

图 3 基于RPS15A氨基酸序列构建的松墨天牛与其他19种昆虫的系统发育树

图 4 农药胁迫下松墨天牛成虫MaRPS15A基因的相对表达量
3讨论
真核生物核糖体由 4 种 RNA 和大约 80 种核糖体蛋白组成。除了在蛋白质合成和调控中发挥基本功能外,核糖体蛋白还参与核糖体以外的生命活动,包括细胞增殖的调节、DNA 修复、转录和 RNA加工[7]。
RPS15A 基因编码一种核糖体蛋白,该蛋白是 40S 核糖体亚基(真核生物核糖体小亚基)的组分。从GenBank数据库相关基因的比对结果看,本研究获得的MaRPS15A基因推测的蛋白属于40S 核糖体亚基。真核生物的核糖体蛋白RPS15A与原核生物的 S8 蛋白同源[7-8]。原核生物的 S8 结合 30S 亚基中的 16S rRNA,在核糖体组装中起关键作用[9-11]。真核生物中,RPS15A在转录起始时发挥功能[12],它可以在翻译早期启动 mRNA 和核糖体的相互作用,并且通过与真核起始因子 4F (eIF-4F) 作用而增强翻译进程。RPS15A 也可能促进细胞生长。斑马鱼(Daniorerio) 中RPS15A基因敲除后严重影响其大脑的发育和生长,循环系统出现问题,尾部发育延迟[13]。由此获得启发:从防治的角度考虑,可以通过 RNA 干扰方法以抑制松墨天牛RPS15A基因的表达,为基因控制松墨天牛及阻断松材线虫的传播提供参考。
核糖体蛋白 RPS15A 在原生动物、细菌和真核生物间都高度保守[14]。从本研究中松墨天牛MaRPS15A与其他昆虫同源序列比对结果来看,MaRPS15A与其他昆虫 RPS15A 蛋白在进化上也高度保守,相似性程度很高,达到88%~98%。系统发育树分析表明,松墨天牛与栎实象鼻虫聚为一支,与白杨叶甲和赤拟谷盗遗传距离也较近,与形态学上的分类相一致,即这些昆虫同属于鞘翅目。
核糖体蛋白基因表达异常将影响核糖体的功能,引发各种疾病,如肿瘤发生、免疫性疾病、代谢性疾病等。有研究表明,个体核糖体蛋白数量的减少会导致非致死性的畸形,这可能是转录效率下降所致[13]。本研究发现,多数农药胁迫下MaRPS15A下调表达,但未发现松墨天牛出现明显的个体畸形现象,这可能是由于观察时间不足:农药处理 12 h 后,松墨天牛就被液氮冷冻用于MaRPS15A表达量分析;更为重要的是,如果在幼虫期就进行农药胁迫,保持亚致死状态饲养,直到成虫,就有可能观测到松墨天牛个体出现畸形的现象。
孟飞等[15]研究了中华蜜蜂(Apisceranacerana)核糖体蛋白 L11基因AccRPLI1在不同环境胁迫下的表达,结果表明,紫外线、高温能在一定时间内使AccRPLI1基因的表达量上升,不同种类的农药如菊酯、蚊蝇醚、辛硫磷、杀螨剂也对AccRPLI1基因 mRNA 的表达量产生不同的影响。本试验中所使用的7种化学农药分属于拟除虫菊酯类(溴氰菊酯、氯氰菊酯),烟碱类(噻虫啉,啶虫脒),有机氮类(杀虫双),氨基甲酸酯类 (灭多威),有机磷类农药 (毒死蜱),4种生物农药分属于真菌 (绿僵菌、白僵菌)、细菌 (Bt) 和植物源农药(印楝素)。这些农药代表了目前常用的主要农药类型,虽然大多数杀虫机理已经明确,但由于机理各异[16-21],研究其对核糖体蛋白基因的影响就具有广泛的代表性。从另一角度看,RPS15A 并不是这些农药传统的直接作用靶标,但农药对昆虫的毒力作用不仅影响主要靶标,还涉及其他代谢途径和生理过程,因而可能有更多的生物分子受到影响。因此,本研究为探究非传统靶标的农药毒理学提供了分子学依据。
本研究中,灭多威胁迫使松墨天牛MaRPS15A下调表达,这是因为许多氨基甲酸盐类农药具有基因毒性,灭多威属于基因毒性制剂,在亚致死浓度下可导致DNA损害和细胞凋亡[22]。灭多威可引起人类周围血液染色体畸变、姊妹染色单体互换、DNA单链断裂[23]。
包括真菌在内的昆虫病原物的杀虫效能不仅依赖于病原物合成的不同毒力因子作用,还依赖于昆虫抑制感染进程或减弱中毒效应的机理。从这个角度而言,昆虫自身的免疫系统调节可以影响药物对其杀害作用[24]。但本研究中绿僵菌作用下松墨天牛MaRPS15A表达下调,认为绿僵菌素可能发挥重要作用。绿僵菌素是从绿僵菌胞外培养基中分离到的真菌毒素,其通过昆虫与真菌之间的进化竞争来特异性抑制昆虫的免疫反应成分[25]。
Tan 等[26]的研究认为,尖音库蚊淡色亚种(Culexpipienspallens) 抗溴氰菊酯品系中的核糖体蛋白 RPL39 的转录水平是敏感品系的 23.4 倍,因而 RPL39 与抗药性的产生有关。改变基因表达可以产生抗性,即基因拷贝数没有发生变化,而是基因调节发生了改变,从而抗药品系比敏感品系的转录水平更高,表达量更多[27]。本研究中,溴氰菊酯胁迫下松墨天牛MaRPS15A表达上调,松墨天牛是否因此会产生抗药性,这需要以后的试验验证。田海生等[28]研究发现,某些核糖体蛋白基因(40S 核糖体蛋白S4 基因等) 在抗性蚊中高表达,而40S 核糖体蛋白S29 基因则在敏感蚊品系中高表达。核糖体蛋白表达量与杀虫剂抗性的关系究竟如何,是通过核糖体蛋白与杀虫剂结合起作用,还是通过核糖体蛋白质间的作用而发挥抗性作用,抑或为其他作用机制值还需进一步探究。
[参考文献]
[1]Alberts B,Johnson A,Lewis J,et al.The molecular biology of the cell [M].New York and London:Garland Publishing,2002:342.
[2]Doudna J A,Rath V L.Structure and function of the eukaryotic ribosome:The next frontier [J].Cell,2002,109(2):153-156.
[3]徐金华,黄秀凤,徐华潮,等.松墨天牛室内人工饲养及其生物学特性观察 [J].浙江林业科技,2009,29(4):86-88.
Xu J H,Huang X F,Xu H C,et al.Rearing and biological properties ofMonochamusalternatus[J].Journal of Zhejiang Forestry Science and Technology,2009,29(4):86-88.(in Chinese)
[4]韦春梅,罗淋淋,吴华俊,等.松墨天牛幼虫cDNA文库的构建及EST分析 [J].基因组学与应用生物学,2014,33(1):113-120.
Wei C M,Luo L L,Wu H J,et al.Construction and analysis ofMonochamusalternatuslarvae EST cDNA library [J].Genomics and Applied Biology,2014,33(1):113-120.(in Chinese)
[5]Livak K J,Schmittgen T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod [J].Methods,2001,25(4):402-408.
[6]罗梅,林进添,宾淑英.扶桑绵蚡蚧环指蛋白基因的克隆及其序列特征与表达谱分析 [J].西北农林科技大学学报:自然科学版, 2014,42(4):125-131.
Luo M,Lin J T,Bin S Y.Molecular cloning, sequence characteristics and expression of ring finger protein gene ofPhenacoccussolenopsisTinsley [J].Journal of Northwest A&F University: Nat Sci Ed,2014,42(4):125-131.(in Chinese)
[7]Wool I G,Chan Y L,Glück A.Structure and evolution of mammalian ribosomal proteins [J].Biochemistry and Cell Biology,1995,73:933-947.
[8]Tishchenko S V,Vassilieva J M,Platonova O B,et al.Isolation,crystallization,and investigation of ribosomal protein S8 complexed with specific fragments of rRNA of bacterial or archaeal origin [J].Biochemistry,2001,66(9):948-953.
[9]Held W A,Ballou B,Mizushima S,et al.Assembly mapping of 30S ribosomal proteins fromEscherichiacolifurther studies [J].Journal of Biological Chemistry,1974,249(10):3103-3111.
[10]Svensson P,Changchien L M,Craven G R,et al.Interaction of ribosomal proteins,S6,S8,S15 and S18 with the central domain of 16S ribosomal RNA [J].Journal of Molecular Biology,1988,200(2):301-308.
[11]Brodersen D E,Clemons W M,Carter A P,et al.Crystal stru-cture of the 30S ribosomal subunit fromThermusthermophilus:Structure of the proteins and their interactions with 16S RNA [J].Journal of Biological Chemistry,2002,316(3):725-768.
[12]Lavoie C,Tam R,Clark M,et al.Suppression of a temperature-sensitive cdc33 mutation of yeast by a multicopy plasmid expressing aDrosophilaribosomalprotein [J].Journal of Biological Chemistry,1994,269(20):14625-14630.
[13]Uechi T,Nakajima Y,Nakao A,et al.Ribosomal protein gene knockdown causes developmental defects in zebrafish [J].PLoS ONE,2006,1:37.
[14]Hulm J L,Kerri B M,Peta C B.Variation in transcript abundance among the four members of theArabidopsisthalianaribosomal proteinS15agene family [J].Plant Science,2005,169:267-278.
[15]孟飞,苍保华,部兴启.中华蜜蜂核糖体蛋白L11基因的克隆及在不同环境胁迫下的表达 [C].华东六省一市生物化学与分子生物学会 2010 年学术交流会.南昌:江西省生物化学与分子生物学学会,2011.
Meng F,Cang B H,Bu X Q.CloningApisceranaceranaribosomal protein L11 gene and its expression in different environmental stress [C].East China Provinces and One City of Biochemistry and Molecular Biology 2010 Symposium.Nanchang:Jiangxi Institute of Biochemistry and Molecular Biology,2011.(in Chinese)
[16]李瑾,李凤良,焦东旭,等.溴氰菊酯胁迫下小菜蛾幼虫体内差异表达蛋白的分析 [J].昆虫学报,2014,57(1):36-44.
Li J,Li F L,Jiao D X,et al.Proteomic analysis of differentially expressed proteins in larvae of the diamondback moth,Plutellaxylostella(Lepidoptera:Plutellidae),under deltamethrin stress [J].Acta Entomologica Sinica,2014,57(1):36-44.(in Chinese)
[17]Zhang Z Y,Zhang C Z,Liu X J,et al.Dynamics of pesticide residues in the autumn Chinese cabbage(BrassicachinensisL.)grown in open fields [J].Pest Management Science,2006,62:350-355.
[18]Villar D,Balvin D,Giraldo C,et al.Plasma and brain cholinesterase in methomyl-intoxicated free-ranging pigeons (Columbaliviaf.domestica) [J].Journal of Veterinary Diagnostic Investigation,2010,22(2):313-315.
[19]Padilla S,Marshall R S,Hunter D L,et al.Time course of cholinesterase inhibition in adult rats treated acutely with carbaryl,carbofuran,formetanate,methomyl,methiocarb,oxamyl or propoxur [J].Toxicology and Applied Pharmacology,2007,219(2/3):202-209.
[20]樊会丹,张从海,严胜骄,等.印楝素的合成、结构修饰及生物活性研究进展 [J].有机化学,2009,29(1):20-33.
Fan H D,Zhang C H,Yan S J,et al.Advances of synthesis and structure modification and bioactivity of azadirachtin [J].Chinese Journal of Organic Chemistry,2009,29(1):20-33.(in Chinese)
[21]赵淑英,盛永丽,姜润田.印楝粗提物的毒性研究 [J].江苏农业科学,2009,6(1):398-399.
Zhao S Y,Sheng Y L,Jiang R T.Studies on toxicity of extracts of neem [J].Jiangsu Agricultural Sciences,2009,6(1):398-399.(in Chinese)
[22]Xiang G G,Li D Q,Yuan J Z,et al.Carbamate insecticide methomyl confers cytotoxicity through DNA damage induction [J].Food and Chemical Toxicology,2013,53:352-358.
[23]Bonatti S,Bolognesi C,Degan P,et al.Genotoxic effects of the carbamate insecticide methomyl:Ⅰ.Invitrostudies with pure compound and the technical formulation “Lannate 25” [J].Environmental and Molecular Mutagenesis,1994,23:306-311.
[24]Beckage N E.Insect immunology [M].Boston:Academic Pre-ss Elsevier,2008:152-160.
[25]Pal S S,Leger R J,Wu L P.Fungal peptide destruxin A plays a specific role in suppressing the innate immune response inDrosophilamelanogaster[J].Journal of Biological Chemistry,2007,282(12):8969-8977.
[26]Tan W B,Sun L X,Zhang D H,et al.Cloning and overexpression of ribosomal protein L39 gene from deltamethrin-resistantCulexpipienspallens[J].Experimental Parasitology,2007,115:369-378.
[27]胡文涛.家蝇对高效氯氰菊酯抗药性机制的探索 [J].中华卫生杀虫药械,2008,14(2):87-90.
Hu W T.Resistance mechanisms ofMuscadomesticaagainst beta-cypermethrin [J].Chin J Hyg Insect & Equip,2008,14(2):87-90.(in Chinese)
[28]田海生,朱昌亮,高晓红,等.淡色库蚊对溴氰菊酯抗药性和敏感性相关基因克隆和序列分析 [J].中国寄生虫学与寄生虫病杂志,2001,19(4):193-197.
Tian H S,Zhu C L,Gao X H,et al.Cloning and identification of deltamethrin-resistance or susceptibility associated genes ofCulexpipienspallens[J].Chin J Parasitol Parasit Dis,2001,19(4):193-197.(in Chinese)
Cloning cDNA of ribosomal protein (RPS15A) fromMonochamusalternatusand effects of insecticides on its expression
LUO Lin-lin,CAI Zi-ling,LIN Tong
(CollegeofForestryandLandscapeArchitecture,SouthChinaAgriculturalUniversity,Guangzhou,Guangdong510642,China)
Abstract:【Objective】 This study explored the expression of ribosomal protein gene under different insecticide stresses to provide theoretical information for control of Monochamus alternatus as an important of pines.【Method】 Expressed sequence tags (ESTs) screened from the formerly constructed cDNA library of M.alternatus were selected randomly for sequencing.The homogenous sequences were obtained from GenBank by Blast.Real time quantitative RT-PCR (RT-qPCR) was used to determine transcript abundance patterns after treatment by 11 insecticides.【Result】 A EST from GenBank relating to insect ribosomal protein S15A (RPS15A) was obtained and named MaRPS15A.The isolated cDNA of MaRPS15A containing both 5′ and 3′ untranslated regions was 504 nucleotide long with an open reading frame of 393 bp and 130 amino acids.The calculated molecular weight was 14.75 ku and the pI was 9.95.The deduced protein sequence of MaRPS15A showed 98% amino acid identity to Triboliumc astaneum RPS15A and >88% identity to RPS15A ribosomal proteins of other insects.All insecticides resulted in decreased transcript abundances except deltamethrin.【Conclusion】 The ribosomal protein gene MaRPS15A was successfully cloned from M. alternatus (GenBank No.KJ930032).Most of the tested insecticides suppressed MaRPS15A expression,indicating that adverse impact on M.alternatus.
Key words:Monochamus alternatus;ribosomal protein;RPS15A;RT-qPCR;treatment with insecticides
DOI:网络出版时间:2016-01-0810:2210.13207/j.cnki.jnwafu.2016.02.023 网络出版时间:2016-01-0810:2210.13207/j.cnki.jnwafu.2016.02.024
[收稿日期]2014-06-20
[基金项目]国家自然科学基金项目(31170612)
[作者简介]罗淋淋(1988-),女,河南许昌人,硕士,主要从事昆虫分子生物学研究。[通信作者]林同(1969-),男,黑龙江通河人,副教授,主要从事昆虫分子生物学研究。E-mail:lintong@scau.edu.cn
[中图分类号]S763.38;Q78
[文献标志码]A
[文章编号]1671-9387(2016)02-0165-07
猜你喜欢
农业灾害研究(2022年11期)2023-01-09
内蒙古民族大学学报(自然科学版)(2022年2期)2022-11-22
小哥白尼(野生动物)(2021年1期)2021-07-16
科技创新与应用(2021年7期)2021-02-04
中国森林病虫(2020年6期)2020-11-28
亚热带植物科学(2020年4期)2020-09-28
肿瘤防治研究(2020年5期)2020-07-09
生物学教学(2019年9期)2019-09-23
小学生必读(低年级版)(2018年10期)2019-01-04
故事作文·低年级(2018年10期)2018-10-25
