
氨基酸新功能及其对脂质代谢和能量平衡的调控
2016-07-25 06:13印遇龙院士中国科学院亚热带农业生态研究所
饲料与畜牧(规模养猪) 2016年6期
印遇龙 院士 中国科学院亚热带农业生态研究所
氨基酸新功能及其对脂质代谢和能量平衡的调控
印遇龙 院士 中国科学院亚热带农业生态研究所

湖南师范大学生命科学学院院长、特聘教授,中国科学院亚热带农业生态研究所畜牧健康养殖中心主任,学术委员会副主任,湖南省政协常委,湖南省海外联谊会常务理事,湖南省九三学社省委委员,动物营养学国家重点实验室、生物饲料安全与污染防控国家工程实验室和中国科学院亚热带农业生态系统过程重点实验室学术委员会委员。
氨基酸是构成动物机体蛋白质结构功能的基本单位。过去一直认为氨基酸只是提供蛋白质合成的原料。25%~54%氨基酸在肠道发生分解代谢,机体蛋白质的沉积实质上是氨基酸的沉积。研究表明,氨基酸对多种生物活性物质的合成也是必需的,并且对动物的生长、发育和肉品质均具有重要作用。下面我们将重点探讨一下氨基酸的新功能及其对脂质代谢和能量平衡的调控。
1 精氨酸对脂质代谢和能量分配的作用
研究表明,日粮添加1%精氨酸,可提高育肥猪胴体瘦肉率,降低脂肪率,改善猪肉质性状,提高肌肉组织4E-BP1、rpS6和eIF4E的磷酸化水平,降低总eIF4G的含量,促进机体蛋白质的合成,减少脂肪的沉积。
精氨酸可差异调控育肥猪肌肉和脂肪组织的脂肪代谢相关酶活性和基因表达,从而降低机体脂肪率、提高肌内脂肪含量,改善肉品质。
研究发现,添加精氨酸能够降低血清中肠道菌群的代谢产物浓度,通过改变脂质信号转导分子浓度来增强机体蛋白质合成和减少脂肪的沉积,提高育肥猪背最长肌粗蛋白含量和脂肪含量(图1),提高育肥猪背最长肌肌糖原含量,从而降低乳酸含量,显著提高屠宰后45min的肌肉pH(图2),提高育肥猪肌肉组织中油酸(C18∶1n-9)含量,而降低了硬脂酸(C18∶0)和亚油酸(C18∶2n-6)含量(图3)。

图1 背最长肌化学组成(g/100g)

图2 背最长肌糖原、乳糖含量及屠宰后45min肌肉pH
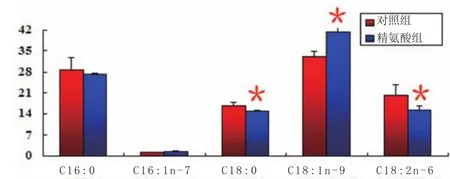
图3 肌肉组织脂肪酸组成(g/100g)
2 谷氨酸对生长猪胴体性状和脂质代谢的调控
常脂日粮添加谷氨酸钠,生长猪的眼肌面积显著降低,脂肪率和肌内脂肪含量显著升高;高脂日粮添加谷氨酸钠,生长猪肌内脂肪含量显著降低,可显著降低背最长肌中单不饱和脂肪酸(MUFA)含量、增加多不饱和脂肪酸(PUFA)含量;高脂日粮添加谷氨酸钠,可增加股二头肌中SFA的含量,降低MUFA含量。表明谷氨酸钠能正向调节肌肉脂肪酸组成,从而改善猪肉品质。
日粮中添加谷氨酸钠和脂肪都能促进肝脏中脂肪酸的装载。谷氨酸钠可增加背部脂肪组织而降低肾周脂中游离脂肪酸的供给,日粮脂肪则使得包含肾周脂在内的脂肪组织的脂肪酸供给加强。
常脂日粮中添加谷氨酸钠可上调背脂中与脂肪合成相关基因的表达,高脂日粮中添加谷氨酸钠则可上调背脂中与脂肪分解相关基因的表达。常脂或高脂日粮中添加谷氨酸钠并不影响生长猪脂肪和肝脏组织中与脂肪酸β-氧化相关的CPT-1基因的表达。
3 支链氨基酸对脂质代谢和能量分配的调控
日粮氮营养素的高效利用主要体现在肌肉组织的增长等方面。支链氨基酸(BCAA)在促进肌肉蛋白质合成方面有着明显优势,而在脂质代谢方面的研究却较少。
Ashley Knight(2013)报道,支链氨基酸成员中的亮氨酸、异亮氨酸和缬氨酸在机体代谢中竞争相同的关键酶(BCAT和BCKDC),说明三者的比例平衡具有重要意义。
以BCAA的平衡为理论基础,初步开展了低蛋白日粮补充不同比例的BCAA对生长猪脂质代谢的影响。
试验表明,生长猪10~30kg阶段,与20%CP相比,17%CP补充BCAAs比例为1∶0.25∶0.25 和1∶0.75∶0.75显著增加股二头肌的肌内脂肪含量,而比例大于1∶0.25∶0.25则提高胴体的脂肪率。与20%的日粮蛋白水平相比,17%的蛋白水平结合平衡的BCAAs可改善不同部位肌肉组织n-3 PUFA含量以及n-6/n-3 PUFA,其中以1∶0.75∶0.75组最佳(见表1)。该结果进一步提示,氨基酸代谢与脂肪酸代谢之间存在互作。
不同的支链氨基酸比例特异性调控不同部位肌肉组织的脂肪代谢,主要通过调节脂肪合成与分解的限速酶以及脂肪酸结合蛋白与转运蛋白相关的基因表达。
试验进一步表明,前期关于脂质代谢的研究主要集中于肌肉组织和脂肪组织,其能量平衡还与两种组织之间的相互作用有关,这种作用关系直接影响到肌肉生长、脂肪沉积和肌内脂肪的形成。肌肉组织和脂肪组织通过肌细胞因子Myokine和脂肪细胞因子Adipokine进行“对话”,为氨基酸的营养调控提供了新的效应靶点。前期研究发现,肌细胞因子Myostain通过激活ERK1/2通路抑制3T3-L1细胞分化,脂肪细胞因子Chemerin通过激活ERK1/2通路抑制C2C12细胞分化。
氨基酸是否调控肌细胞因子或脂肪细胞因子的分泌?并通过这些因子调节肌肉组织和脂肪组织的脂质代谢与能量分配?这些问题都有待进一步深入研究。

表1 试验分组

图4 胰岛素抵抗的形成过程中,脂肪细胞周围的血管生成减少,巨噬细胞的渗透增强
4 亮氨酸对肌细胞因子和脂肪细胞因子分泌的调节
利用Transwell小室将分化后的C2C12肌管细胞与3T3-L1成熟的脂肪细胞进行共培养,考察亮氨酸对脂质代谢调控相关细胞因子表达的调节。研究表明,肌细胞与脂肪细胞共培养条件下,亮氨酸上调mTOR而下调AMPKβ1磷酸化蛋白的表达水平,细胞事件趋于消耗能量的蛋白合成。经分泌蛋白质组学分析,筛选得到4个倍数大于1.2的上调蛋白和6个倍数小于0.75的下调蛋白,其具体功能有待进一步挖掘。脂肪细胞与肌细胞共培养条件下,亮氨酸促进脂肪细胞因子Chemerin、肌细胞因子IL-6和生长因子家族IGFBP-3的分泌表达,而降低IGFBP-2、IGFBP-5、IGFBP-6的分泌表达。这些因子将继续发挥细胞之间的能量调节作用。共培养条件下,亮氨酸上调脂肪细胞PPARγ 和FAS的基因表达,促进脂肪的合成。同时,我们发现mTOR信号通路抑制剂雷帕霉素显著抑制与脂肪生成相关的基因,促进脂肪的分解。
5 谷氨酰胺调节脂肪细胞和巨噬细胞的互作
胰岛素抵抗的形成过程中,脂肪细胞周围的血管生成减少,巨噬细胞的渗透增强(图4)。谷氨酸依赖谷氨酰胺合成酶(GS)生成谷氨酰胺,GS在分化成熟的脂肪细胞中高表达,而在巨噬细胞中的表达量非常低。因此,推测脂肪细胞为巨噬细胞提供谷氨酰胺作为能量来源。前脂肪细胞分化过程中的分子c/ EBPβ、PPARγ以及c/EBPα的表达都需要谷氨酰胺。脂肪细胞与巨噬细胞共培养增加了促炎性因子(IL-6、MCP-1、RANTES)的分泌表达。

6 N-氨甲酰谷氨酸(NCG)对育肥猪生长发育
N-氨甲酰谷氨酸(NCG)不会影响赖氨酸、组氨酸和色氨酸的吸收,还可以持续激活精氨酸和瓜氨酸的合成,同时具有高效稳定、使用成本低及在肠道黏膜中半衰期长(8~10h)等优点。吴琛等(2011)为了研究添加NCG对环江香猪胴体性状的影响,在28~33kg环江香猪日粮中添加800g NCG,发现可显著降低脂肪率和均膘厚,显著提高屠宰率。
7 半胱胺改善猪肉品质
半胱胺的巯基(-SH)自身具有很高的还原性。并且,半胱胺是谷胱甘肽的组成成分之一,能够提高血液中GSH的浓度,提高动物对抗高温应激氧化损伤的能力。
添加包膜半胱胺能够提高育肥猪的生长性能、胴体性能;改善猪肉保水性及风味;保持长时间贮存条件下肉色状态。研究表明,添加量为70mg/kg,对改善生长性能、胴体性能及肉质的效果相对较好。
8 研究展望
①精氨酸通过营养感受体调控棕色脂肪以及白色脂肪棕色化相关方面的研究。
②亮氨酸调控猪肌肉组织和脂肪组织能量代谢平衡方面的研究。
③亮氨酸中间代谢产物HMB 和α-KIC对猪脂质代谢的调节作用。
④日粮蛋氨酸限制不仅调控脂肪代谢,且改善机体的整体代谢,相关机制都有待进一步揭示。
⑤以脂肪细胞为中心,研究其与肌细胞、肝细胞、乳腺上皮细胞和血管内皮细胞之间的互作,为改善猪的脂质代谢与能量分配提供参考。
(编辑张瑞霜整理)
猜你喜欢
科学大众(2022年10期)2022-05-26
肉类研究(2020年9期)2020-12-14
科技资讯(2020年3期)2020-04-07
特别文摘(2019年5期)2019-02-28
分析化学(2018年4期)2018-11-02
新疆农垦科技(2018年11期)2018-02-13
肉类研究(2017年8期)2017-11-16
中成药(2017年6期)2017-06-13
动物营养学报(2015年10期)2015-12-01
江苏农业科学(2014年8期)2014-10-23
