
波斯婆婆纳的繁殖能力及其入侵原因探析
2016-08-10 09:15谢翠容汤林彬刘茗枫戴璨湖北大学资源环境学院湖北武汉430062湖北省生物资源绿色转化协同创新中心湖北武汉430062区域开发与环境响应湖北省重点实验室湖北武汉430062
生态环境学报 2016年5期
关键词:传粉
谢翠容,汤林彬,刘茗枫,戴璨*1. 湖北大学资源环境学院,湖北 武汉 430062;2. 湖北省生物资源绿色转化协同创新中心,湖北 武汉 430062;3. 区域开发与环境响应湖北省重点实验室,湖北 武汉 430062.
波斯婆婆纳的繁殖能力及其入侵原因探析
谢翠容1§①,汤林彬1§②,刘茗枫1③,戴璨1, 2, 3*
1. 湖北大学资源环境学院,湖北 武汉 430062;2. 湖北省生物资源绿色转化协同创新中心,湖北 武汉 430062;3. 区域开发与环境响应湖北省重点实验室,湖北 武汉 430062.
摘要:入侵植物是近年来生态关注的热点。成功入侵植物的有性和无性繁殖能力通常呈现特殊性和优势性。探究其繁殖特征,对于应对与管理植物入侵问题具有重要的生态与经济意义。以波斯婆婆纳(Veronica persica)为对象,多方面研究其繁殖能力,以期探究波斯婆婆纳在短时间内迅速扩张成为全球广泛入侵杂草的原因。进行了波斯婆婆纳开花与传粉机制的观察实验;对波斯婆婆纳自然种群的生长密度和植株构建进行调查;并通过自然传粉和人工授粉实验比较花、茎长、果实的差异,通过同质园移栽和生长实验对波斯婆婆纳的有性和无性繁殖能力进行量化和评价。结果表明,波斯婆婆纳的有性繁殖依赖于昆虫传粉,自交亲和;在自然环境、人工自花授粉和补充授粉3种方式下均有较高的座果率和结实率;种子萌发迅速,萌发率达70%,表明其有较强的有性繁殖能力。同时,波斯婆婆纳的营养生长密度大,匍匐茎发达,平均每个植株能产生8条左右的直立茎。波斯婆婆纳移栽实验的存活率达到83%,且茎长在40天平均增长6.5 cm,说明其具有较强的生长和无性繁殖能力。波斯婆婆纳的营养生长和有性繁殖之间有着极强的相关性。波斯婆婆纳繁殖能力强,环境限制因子少,为其成功地大范围扩张而成为全球广泛入侵杂草提供了重要条件。
关键词:无性繁殖;传粉;有性繁殖;波斯婆婆纳
引用格式:谢翠容, 汤林彬, 刘茗枫, 戴璨. 波斯婆婆纳的繁殖能力及其入侵原因探析[J]. 生态环境学报, 2016, 25(5): 795-800.
XIE Cuirong, TANG Linbing, LIU Mingfeng, DAI Can. The Reproductive Capacity of Veronica Persica and Implications for Its Worldwide Invasion [J]. Ecology and Environmental Sciences, 2016, 25(5): 795-800.
物种入侵被认为是全球变化的一个重要组成因素(Sakai et al.,2001)。入侵杂草是外来植物中具有侵略扩张性的植物物种,在全世界范围内已经造成了巨大的生态破坏和经济损失(桑卫国,2002)。探究杂草的入侵机制和繁殖特征是防止和应对入侵的关键,也是制定有效管理措施的重要依据(Forman et al.,2003;Ruaux et al.,2009)。对于成功入侵的植物而言,诸多研究表明其繁殖能力(徐汝梅等,2003;黄乔乔等,2012)、抗干扰能力(Alpert et al.,2000)、较宽泛的生态幅(宫伟娜等,2005;冯玉龙等,2009;Zhang et al.,2012)、较强的自我防御机制(倪广艳等,2014)和竞争能力(余香琴等,2010)等在其扩张和归化的过程中具有十分重要的作用。Baker(1974)总结外来入侵杂草的12项主要生态学特征,如具有自交亲和性、花果期长、种子产量高、发芽的环境类型广泛、无性繁殖能力强等,一大部分都是和植物的有性和无性繁殖能力直接相关的特性,表明外来植物的成功入侵在一定程度上取决于其繁殖能力(郝建华等,2009;贺俊英等,2009)。
波斯婆婆纳(Veronica persica),又名阿拉伯婆婆纳,属双子叶植物玄参科婆婆纳属的一年生或两年生的全球性入侵杂草。原产于欧洲、亚洲西部至伊朗,现广泛分布于温带、亚热带地区。在我国分布范围极广,全国19个省和3个直辖市有分布(李扬汉,1998),主要生长于长江沿岸及西南部分地区的旱地,常形成优势居群。同大多数植物一样,波斯婆婆纳可以通过有性和无性两种繁殖方式繁衍后代,有性繁殖为主要方式(淮虎银等,2004)。有关研究推测其成功入侵是因强大的繁殖能力造成(李扬汉,1998)。
目前国内外对波斯婆婆纳的研究主要集中在其入侵与危害(吴海荣,2006)、化感作用(李美娜等,2001;吴海荣等,2006)、生态经济阈值(李建波,2003)、药用价值(田亮等,2004;张仁波等,2009)等方面。而对于它的繁殖能力,有少量研究指出波斯婆婆纳具有较强的不定根生长能力,可助其繁殖扩散(Harris et al.,1980a,1980b),无性繁殖对其有性繁殖有一定的贡献作用(淮虎银等,2004),此外,波斯婆婆纳的种子萌发条件和资源分配也是其繁殖扩散的重要条件(Harris et al.,1980a)。目前,对于波斯婆婆纳在自然条件下无性繁殖的水平、个体的扩张程度以及在不同生境中存活和生长的能力等方面的研究较为缺乏。在有性繁殖方面,国内的相关研究仅涉及到种子重量、单株结实率等(吴海荣,2006);国外仅有Tsuruuchi (1994)比较了波斯婆婆纳与常青藤婆婆纳的主动自花授粉(Autogamy)能力。目前,对波斯婆婆纳在不同传粉方式下的结实水平、不同生境下的有性繁殖能力等都缺乏了解。据此,本文拟通过对波斯婆婆纳自然居群的有性和无性繁殖能力进行研究,以期揭示其自然居群的繁殖策略,探讨波斯婆婆纳广泛入侵的原因。
1 材料与方法
1.1 材料与样方
波斯婆婆纳为矮小草本植物,在湖北武汉地区2月中旬发芽,3月进入花期,4月大部分个体进入果期,常大片生长在路边和绿化带。根据人为扰动强度、居群规模(面积和植株密度)等在湖北大学及周边地区选择了19个样地,样地信息见表1。
1.2 实验方法
自然居群实验,(1)花期观测:随机选取5个样地中的50个未开的花苞进行跟踪,在连续3天以上的晴好天气观测波斯婆婆纳花的开放和闭合时间,花凋谢则表示花期结束。(2)访花昆虫观测:于天气晴好的不同时间段(10:00—16:00)在11个样方地各随机选择10个植株(每个植株1朵花)进行 20分钟的连续观察,并记录在规定时间内访花昆虫的种类和数量以及每一个昆虫个体所接触的花朵数。(3)生长情况调查:用皮尺随机圈出0.5 m×0.5 m的范围作为单位面积,统计波斯婆婆纳植株覆盖率,计数和测量单位面积内的植株茎数、株数、地下茎的长度和个数。波斯婆婆纳可在地表或土下浅层生长匍匐茎,由匍匐茎相连的各个地上直立茎构成一个独立的植株(基因型),茎数是地上部分独立茎的数量,株数是辨清地下关系后的植株数量,地下茎的长度和个数由每个直立茎延生的地下茎结构得出,选取地下茎最长的3个。(4)胚珠数量统计:摘取完全开放的花,剥离出子房,置于宽视场体视显微镜下用解剖针和镊子进行解剖,记录胚珠数。(5)自然座果率和结实率统计:于2013 和 2014年进行两次观察和统计,随机标记开放的花100朵,待果实成熟后收回并统计种子数。(6)人工授粉实验:于晴天的下午对100朵自然授粉的花朵进行人工补充授粉,用小排笔蘸取其他花朵上的花粉,均匀涂抹到柱头上,待结实后统计种子数量。(7)花、茎长、果实比较实验:于植株成熟期,随机选取22个植株,统计每株的花和果实的数量,并测量茎长。(8)种子重量测量实验:于果实成熟期,随机选取150个饱满预裂开的果实,剥离出种子若干,计数后使用电子天平称量并换算为种子千粒重。

表1 波斯婆婆纳的样地信息Table 1 The information on sampling sites of Veronica persica
同质园实验:于2014年3月初在19个样方内挖取根长为1 cm左右,初始直立茎数量为3的植株100株,摘去多余的叶和花,只留取茎基部的2~3片叶子,分盆编号栽种于湖北大学资源环境学院的二楼平台(人为干扰较少、半阴生),记录每株波斯婆婆纳的初始茎长。生长40 d后,统计成活率,并量取存活植株生长后的茎长和直立茎数量。同时,于花期对同质园内随机选取的100朵花进行自花授粉实验。由于同质园光照有限,少有昆虫访问,所有花在凋谢时柱头无花粉落置,花粉囊饱满,故可在无需套袋的情况下进行人工授粉:用放大镜观察确定柱头上无花粉,再用小排笔将花粉涂抹到同一朵花的柱头上,而后摘去花瓣,避免昆虫访花,待果实成熟后摘取记录种子数。
萌发实验:于2013年4—5月在上述19个样方中随机采集波斯婆婆纳果实75个,共计767粒种子,将所有种子混合并装入封口袋,4 ℃冰箱保存待用。于2014年12月将所有种子随机均匀的放入50个铺有浸湿滤纸的培养皿(每个培养皿有种子 15颗左右),将培养皿放入人工气候箱,萌发条件为15 ℃、每日光照时间14 h、定期补水。实验周期为10 d,每隔24 h统计1次发芽种子数,累计统计萌发率。
1.3 统计方法
用卡方分析进行不同授粉方式的座果率的比较和同质园内不同环境的成活率比较;用Student's t检测比较两年的自然结实差异;用方差分析比较不同授粉方式的结实差异;用Pearson相关系数检验花、茎长与果实的相关性;用成对Student's t检测判断同质园内植株的营养生长状况。
2 结果与分析
2.1 花期与传粉
波斯婆婆纳的花大约在上午8点开放,下午17点闭合,单天开花时长约9 h。在天气晴好的情况下,波斯婆婆纳的单花花期为(2.13±0.07)d。多数波斯婆婆纳植株在光照充足、温度适宜时开花,而在阴雨天或者光照不足时其花闭合,待天气状况好时再度开放,花的开放过程可逆。
波斯婆婆纳花的主要访问昆虫为小型蜂类和蝇类,访花频率为(0.54±0.12)次/小时/朵。在20个时间段的观察中,有8个时间段没有观察到昆虫访问。由于观察选取的时间段(11:00—13:00)是昆虫最为活跃的时间,表明昆虫的访花频率不高。
2.2 胚珠数和自然结实
在随机抽取的 91个波斯婆婆纳的子房中,最少胚珠数为9,最多胚珠数为19,平均胚珠数量为(13.7±0.2)。2013年的自然结实平均每果产生(11.9±0.3)颗种子,结实率为86.3%。自然成熟的种子千粒重为0.464 g。
2.3 不同授粉方式下的座果和结实
2014年波斯婆婆纳自然授粉、自花授粉和补充授粉的座果率均处于中高水平(图1),且自花授粉与补充授粉的座果率存在显著差异(χ2=6.57,P=0.01)。
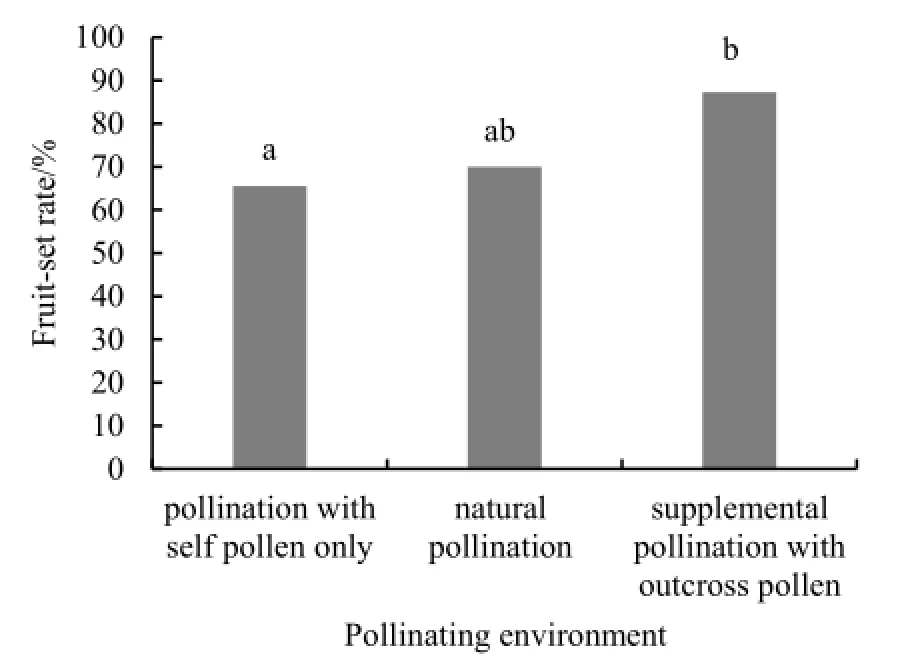
图1 2014年各种传粉环境下波斯婆婆纳的座果率Fig. 1 The fruit-set of Veronica persica under different pollinating environments in 2014
2014年自然结实平均每果有(10.14±0.51)颗种子,显著低于2013年自然结实的水平(t=2.63,P=0.0096;图2)。
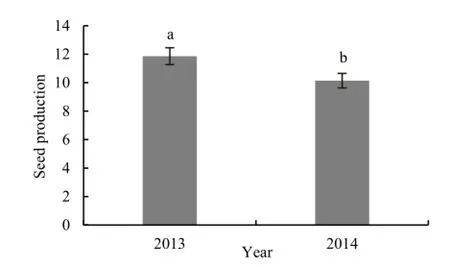
图2 2013和2014年波斯婆婆纳的自然结实Fig. 2 The seed production of Veronica persica under natural pollination in 2013 and 2014
2014年各种授粉方式结实后的种子数量相当,除了上述的自然结实,自花授粉后平均每果有(10.08±0.52)颗种子,补充授粉后平均每果有(10.58±0.46)颗种子,三者没有显著差异(F(2,110)=0.31,P=0.73)。
2.4 花、茎长、果实相关实验
波斯婆婆纳的花、茎长和果实数量两两间的相关性达到极显著水平(花与茎长:r=0.84,P<0.00001;果实与茎长:r=0.76,P<0.0001;花与果实:r=0.95,P<0.00001),从图3中可知,花与果实的数量随茎长的增减而同步上升或下降。
2.5 各种无性繁殖指标
单位面积的波斯婆婆纳株数与茎数变异较大(表2)。波斯婆婆纳的每段匍匐茎长度多在5~6 cm,也就是平均每5~6 cm可伸出一条直立茎。波斯婆婆纳个体间存在着明显的大小差异,地上直立茎数量在1~32条·株-1之间,平均每个植株产生8条左右的直立茎。波斯婆婆纳可产生3~6条一级匍匐茎,此后一级匍匐茎进一步伸长后于节处形成次一级匍匐茎或直立茎。
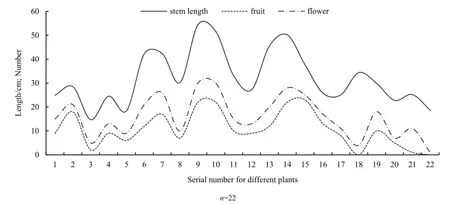
图3 同株波斯婆婆纳的茎长、花朵数和果实数Fig. 3 The stem length, number of flowers and fruits within individual plants of Veronica persica
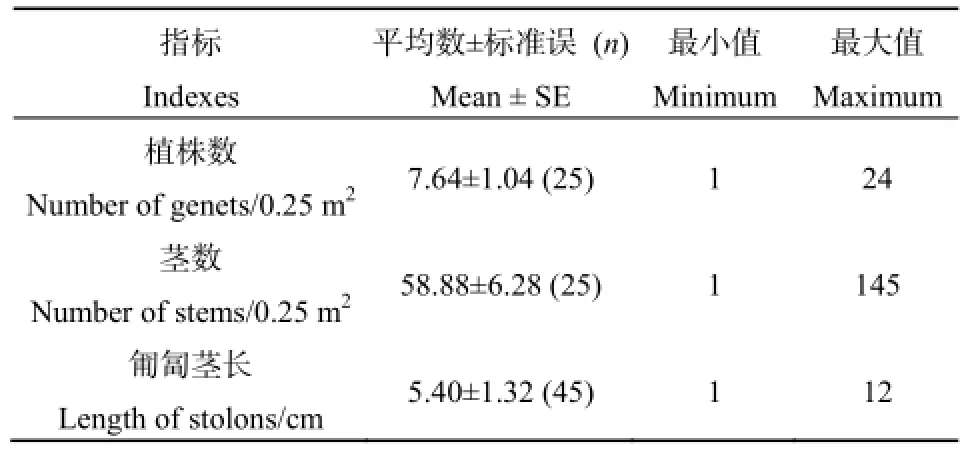
表2 波斯婆婆纳的无性繁殖各种指标Table 2 The indexes of asexual reproduction in Veronica persica
2.6 同质园的存活和营养生长
在人工构建的同质园中,波斯婆婆纳移栽后的存活率为83%。推测水分与光照条件影响了波斯婆婆纳的成活。编号前一半的盆栽植株被放置于易受阳光与风雨影响的平台边缘区,而编号后一半的在有遮蔽物的平台内部区。由于2014年3—4月多为阴雨天气(40 d中18 d为雨天),边缘区的盆栽土壤明显比内部区的潮湿。边缘区的植株成活率为76%,而内部区为90%,两者之间有接近显著的差异(χ2=3.47,P=0.06)。
通过同质园40 d的生长实验,波斯婆婆纳的茎长增长显著(paired t=22.94,P<0.0001),各植株单茎平均增长(6.5±2.7)cm。
2.7 种子萌发
波斯婆婆纳的种子置于培养皿2 d后即开始萌发,4~5 d进入高峰期,7 d左右萌发结束,其种子萌发率为70%。
3 讨论
3.1 波斯婆婆纳年间繁殖能力的差异
实验发现自然状态下 2013年波斯婆婆纳的种子数显著高于 2014年。同为固定样方的随机取样和自然状态下的传粉,年间的气候差异可能对种子产量有着显著的影响。实验期间(3—5月)2013年武汉的平均温度为 20.75 ℃,降水量为 304.1 mm;而2014年为14.9 ℃,比2013年低5.85 ℃,降水量为317 mm,与2013年相差不大。在降雨量相当而温度较高的 2013年,波斯婆婆纳有更高的种子数量,意味着具更强的有性繁殖能力。温度对其有性繁殖可能有一定的制约作用,也许是通过影响植株的生长速率和传粉昆虫的活动造成的(朱建华等,2010;闫志强等,2014)。不过,从两年的结实率来看,即便是低水平的 2014年也达到了70%,表明波斯婆婆纳在一定的环境梯度内都具有较强的有性繁殖能力,这也与吴海荣(2006)的调查实验相吻合:在野外生境中,波斯婆婆纳的单株结实量及单果种子均显著高于同属的直立婆婆纳(Veronica arvensis)与睫毛婆婆纳(Veronica hederifolia)。
3.2 不同授粉方式对波斯婆婆纳有性繁殖能力的影响
波斯婆婆纳在补充授粉的情况下座果率略高于自然状态,这可能是因为波斯婆婆纳自然居群的授粉并没有达到完全饱和,存在一定程度的花粉受限,这与实验观测到的传粉昆虫低水平的访花频率是一致的。这一结果与同质园极低的座果率均体现了波斯婆婆纳的有性繁殖需要传粉昆虫的参与,且不具有自花授粉的能力(Tsuruuchi,1994)。
波斯婆婆纳自花授粉的座果率显著低于补充授粉下的座果率。造成该差异的原因可能是由于进行波斯婆婆纳自身花粉数量有限,无法达到较高水平的座果率。
此外,波斯婆婆纳自花授粉所得种子数与自然状态和补充授粉情况下的种子数差异不大,表明该物种自交亲和,在无其他植株提供花粉进行异交的情况下,可以依赖昆虫转移自花花粉产生后代。在入侵植物建立居群的初期可能只有少数的个体参与有性繁殖,其自交亲和性和低水平的自交衰退也许是波斯婆婆纳得以快速、大范围入侵的原因之一(Tsuruuchi,1994;曾建军等,2010)。
3.3 波斯婆婆纳种子的萌发
波斯婆婆纳的种子在15 ℃时(该物种在实验地区自然条件下萌发的正常温度)萌发速度快,萌发率达到70%。在Harris et al.(1980a)的实验中,在25 ℃的温度下波斯婆婆纳的种子萌发率达到了100%。结合其他研究(Boutin et al.,1991)表明,波斯婆婆纳种子萌发受季节因素限制较小,且越冬的种子死亡率较低,这为该物种的广布和快速入侵提供了有利条件。同时波斯婆婆纳种子千粒重小,相比同属的婆婆纳(Veronica polita)、直立婆婆纳和睫毛婆婆纳质量更轻(吴海荣,2006),使其适合于风力扩散和远距离传播,容易入侵到新生境中繁殖扩张。
3.4 波斯婆婆纳的营养构件和生长及其对无性繁殖的贡献
波斯婆婆纳自然居群的生长密度大,单株直立茎数量多,匍匐茎长,说明该物种具有很强的蔓延性,可依靠匍匐茎不断占领新的空间。随着生长周期日渐成熟,单株波斯婆婆纳的地面覆盖面积也会随之加大,利于其争夺水分和养分,影响同一环境中其他植物的生长繁殖,并形成优势居群。
同质园实验结果显示,波斯婆婆纳具有较高的移栽成活率,凡成活的植株均能开花,且具有短期生长迅速的特点。外来入侵杂草植株不定根发生率往往较高(桑卫国,2002),本实验也证实了这一点:当植株的直立茎过长导致部分贴服于地表时,能长出许多不定根,有利于形成新的克隆分株。已有研究指出,相比于V. agrestic和V. hederifolia,波斯婆婆纳具有更强的生长不定根的能力,甚至在其根尖或基部被人为截断的情况下都能短时间内长出大量新的不定根(Harris et al.,1980b)。也有研究表明,波斯婆婆纳的营养构件片段有很强的发育成独立个体的能力(淮虎银等,2004)。由此可见,波斯婆婆纳快速的不定根生长、营养器官发育成独立植株的能力和强大的存活力是其在全球范围内扩散的优势所在。
绝大多数外来杂草为旱生阳生植物,例如北美车前草(Plantago virginica)、豚草(Ambrosia artemisiifolia)等,这一生理特性使得外来杂草适生于弃耕荒地、宅旁、路边等阳生环境中(吴海荣等,2004;罗文启等,2015)。同质园的生长实验土壤来源于自然裸露荒地,有机质含量低,在土质较差的土壤环境中,波斯婆婆纳的成活率仍达到83%,说明其具有较强的耐贫瘠性。在相对潮湿环境中,波斯婆婆纳存活率明显低于旱生环境,表明该物种更适应于路边、弃田荒地、宅旁等旱生生境。
4 结论
通过对波斯婆婆纳有性和无性繁殖能力的自然居群和同质园移栽实验,结果发现其自交亲和、自交衰退不明显;有较高的座果率和结实率;种子具有较快的萌发速度、较高的萌发率和较低的千粒重。表明波斯婆婆纳能依靠其强大的种子繁殖能力占据新领地及其在新生境中的快速萌发保证了其成功入侵。同时植株还具有较强的适应性和耐受力;利用不定根和匍匐茎扩大生长范围,增加植株构建数量,迅速扩张形成优势居群,提高自身与其他植物的竞争能力和抵御外来风险的能力。波斯婆婆纳的有性繁殖水平随着营养生长的增强而不断提高,这使得其在旧领地的巩固扩张和新领地的入侵达到互相促进的效果。上述特点给波斯婆婆纳防除治理带来了较大困难,由于其地上和地下的大量生物量积累和充足的土壤种子库,各种防除方法可能只能在短期内取得一定的成效。依据本研究结果,建议综合考虑波斯婆婆纳种子快速萌发和生长、依赖昆虫传粉的结实、种子散布方式以及生境的土壤水分状况等因素,制定适用于该物种的防除策略。
致谢:感谢杨玉雪、王亚妮在野外实验中的协助,李婷、覃道凤在解剖实验中的指导和帮助。感谢罗文杰在本文写作过程中的帮助。
参考文献:
ALPERT P, BONE E, HOLZAPFEL C. 2000. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants [J]. Perspectives in Plant Ecology, Evolution and Systematics, 3(1): 52-66.
BAKER H G. 1974. The evolution of weeds [J]. Annual Review of Ecology and Systematics, 5(1): 1-24.
BOUTIN C, HARPER J L. 1991. A comparative study of the population dynamics of five species of Veronica in natural habitats [J]. Journal of Ecology, 79(1): 199-221.
FORMAN J, KESSELI R V. 2003. Sexual reproduction in the invasive species Fallopia japonica (Polygonaceae) [J]. American Journal of Botany, 90(4): 586-592.
HARRIS G R, LOVELL P H. 1980a. Growth and Reproductive Strategy in Veronica spp [J]. Annals of Botany, 45(4): 447-458.
HARRIS G R, LOVELL P H. 1980b. Adventitious root formation in Veronica spp [J]. Annals of Botany, 45(4): 459-468.
RUAUX B, GREULICH S, HAURY J, et al. 2009. Sexual reproduction of two alien invasive Ludwigia (Onagraceae) on the middle Loire River,France [J]. Aquatic Botany, 90(2): 143-148.
SAKAI A K, ALLENDORF F W, HOLT J S, et al. 2001. The population biology of invasive species [J]. Annual Review of Ecology and Systematics, 32(1-1): 305-332.
TSURUUCHI T. 1994. Reproductive ecology of Veronica hederaefolia L. and V. persica Poir [J]. Weed Research (Japan), 39(2): 85-90.
WILLIAMS D G, MACK R N, BLACK R A. 1995. Ecophysiology of introduced Pennisetum setaceum on Hawaii: the role of phenotypic plasticity [J]. Ecology, 76(5): 1569-1580.
ZHANG Y Y, FENG J C, SANG W G, et al. 2012. Distribution of invasive plant species in relation to environmental and anthropogenic factors in five Nature Reserves in northern China [J]. Journal of Resources and Ecology, 3(3): 278-283.
曾建军, 肖宜安, 孙敏. 2010. 入侵植物剑叶金鸡菊的繁殖特征及其与入侵性之间的关系[J]. 植物生态学报, 34(8): 966-972.
冯玉龙, 廖志勇, 张茹, 等. 2009. 外来入侵植物对环境梯度和天敌逃逸的适应进化[J]. 生物多样性, 17(4): 340-352.
宫伟娜, 万方浩, 谢丙炎, 等. 2009. 表型可塑性与外来入侵植物的适应性[J]. 植物保护, 35(4): 1-7.
郝建华, 刘倩倩, 强胜. 2009. 菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系[J]. 植物学报, 44(6): 656-665.
贺俊英, 谢彩琴. 2009. 繁殖生物学特性在外来入侵植物入侵性中的意义[J]. 内蒙古师范大学学报: 自然科学汉文版, 38(2): 217-221.
淮虎银, 张彪, 张桂玉, 等. 2004. 波斯婆婆纳营养生长特点及其对有性繁殖贡献[J]. 扬州大学学报: 农业与生命科学版, 25(3): 70-73.
黄乔乔, 沈奕德, 李晓霞, 等. 2012. 外来入侵植物在中国的分布及入侵能力研究进展[J]. 生态环境学报, 21(5): 977-985.
李建波. 2003. 麦田婆婆纳生态经济阈值的研究[J]. 安徽农业科学,31(6): 1062-1062.
李美娜, 李志华. 2010. 紫花苜蓿茎叶浸提液对波斯婆婆纳化感效应的研究[J]. 中国草地学报, 32(1): 69-74.
李扬汉. 1998. 中国杂草志[M]. 北京: 中国农业出版社: 936-937
罗文启, 符少怀, 杨小波, 等. 2015. 海南岛入侵植物的分布特点及其对本地植物的影响[J]. 植物生态学报, 39(5): 486-500.
倪广艳, 朱丽薇, 牛俊峰, 等. 2014. 三种菊科入侵植物的生长与化学防御的关系研究[J]. 生态环境学报, 23(1): 1-6.
桑卫国. 2002. 我国外来杂草入侵特点的初步分析[J]. 中国农业科技导报, 4(5): 59-61.
田亮, 周金云. 2004. 婆婆纳属植物的研究进展[J]. 中药材, 27(1): 67-70.
吴海荣, 强胜, 段惠, 等. 2004. 波斯婆婆纳[J]. 杂草科学, 33(4): 46-49.
吴海荣, 强胜. 2008. 外来杂草波斯婆婆纳的化感作用研究[J]. 种子,27(9): 67-69.
吴海荣. 2006. 南京地区外来杂草调查及婆婆纳属外来杂草入侵性特征比较研究[D]. 南京: 南京农业大学.
徐汝梅, 叶万辉. 2003. 生物入侵: 理论与实践[M]. 北京: 科学出版社: 39-52.
闫志强, 刘黾, 吴小业, 等. 2014. 温度对五种沉水植物生长和营养去除效果的影响[J]. 生态科学, 33(5): 839-844.
余香琴, 冯玉龙, 李巧明. 2010. 外来入侵植物飞机草的研究进展与展望[J]. 植物生态学报, 34(5): 591-600.
张仁波, 窦全丽. 2009. 国内婆婆纳属药用植物研究进展[J]. 科技资讯,31: 242-242.
朱建华, 徐宁, 王助引, 等. 2010. 龙眼传粉昆虫种类及其传粉活动与温度的关系[J]. 热带作物学报, 31(4): 646-650.
§对本文具有同等贡献
现地址:①武汉大学资源与环境科学学院,湖北 武汉 430072;②中国科学院城市环境研究所,福建 厦门 361021;③香港中文大学太空与地球信息科学研究所,香港 999077
DOI:10.16258/j.cnki.1674-5906.2016.05.010
中图分类号:X173; Q948
文献标志码:A
文章编号:1674-5906(2016)05-0795-06
基金项目:国家自然科学基金项目(31270279);教育部留学回国人员科研启动基金项目(第46批)
作者简介:谢翠容(1990年生),女,硕士研究生,研究方向为植物生态。E-mail: 243176799@qq.com
*通信作者:戴璨(1983年生),女,副教授,博士,主要从事植物生态与进化方面的研究。E-mail: daican@hubu.edu.cn
收稿日期:2015-12-23
The Reproductive Capacity of Veronica Persica and Implications for Its Worldwide Invasion
XIE Cuirong1§①, TANG Linbing1§②, LIU Mingfeng1③, DAI Can1, 2, 3*
1. School of resources and Environmental Science, Hubei University, Wuhan 430062, China;2. Hubei Collaborative Innovation Center for Green Transformation of Bio-Resources, Wuhan 430062, China;3. Hubei Key Laboratory of Regional Development and Environmental Response, Wuhan 430062, China Current Address: ① School of Resources and Environmental Science, Wuhan University, Wuhan 430070, China;② Institute of Urban Environment, Chinese Academy of Sciences, Xiamen 361021, China;③ Institute of Space and Earth Information Science, The Chinese University of Hong Kong, Hong Kong 999077, China § These authors had equal contribution to this paper
Abstract:Invasive plants have become a hot research topic in recent years. For successful invasive species, the capacity of sexual and asexual reproduction contributes particularity and advantages to their invasion. Exploring the reproductive characteristics of certain invasive species can help us effectively respond and manage invasion from both ecological and economic perspectives. In this study, we examined the reproductive ability of Veronica persica, with the aim of explaining how it can expand its distribution in a short period of time and finally become a worldwide invasive weed. We have conducted observational experiments on flowers and pollinators, surveys on the density of natural populations and segments of individual plant. We have also quantified the level of sexual and asexual reproduction in V. persica through the experiments of natural and artificial pollination, and common garden transplant and growth. The results showed that the sexual reproduction of V. persica was pollinator-dependent and self-compatible. There was high level of fruit-set and seed-set under natural, self, and supplemental pollination, and the seeds germinated fast with 70% germination rate, suggesting a relatively strong sexual reproductive ability. Meanwhile, the density of V. persica in natural populations was high, with highly developed stolons and around 8 ramets per genet. The transplant experiment of V. persica yielded 83% survivorship and rapid growth of 6.5 cm in 40 days, indicating a high level of asexual reproduction. Our results have also pointed out that there is strong correlation between the vegetative growth and sexual reproduction in V. persica. As a whole, the strong reproductive ability and few restrictive environmental factors probably contribute a lot in the successful worldwide invasion of V. persica.
Key words:asexual reproduction; pollination; sexual reproduction; Veronica persica
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
科学之友(2021年12期)2021-12-23
中国蜂业(2021年4期)2021-04-25
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
生物学教学(2019年1期)2019-01-11
大科技·百科新说(2018年3期)2018-04-11
科学种养(2017年11期)2017-11-28
中国蜂业(2016年7期)2016-01-29
中国蜂业(2015年7期)2015-01-24
