
高寒草甸植物正构烷烃特征分析
2016-09-01 07:16段中华全小龙乔有明裴海昆何桂芳
草业学报 2016年6期
段中华,全小龙,乔有明*,裴海昆,何桂芳
(1.青海大学生态环境工程学院,青海 西宁 810016;2.三江源生态和高原农牧业国家重点实验室,青海 西宁 810016;3.青海大学分析测试中心,青海 西宁 810016)
高寒草甸植物正构烷烃特征分析
段中华1,2,3,全小龙1,乔有明1*,裴海昆3,何桂芳1
(1.青海大学生态环境工程学院,青海 西宁 810016;2.三江源生态和高原农牧业国家重点实验室,青海 西宁 810016;3.青海大学分析测试中心,青海 西宁 810016)
为研究高寒草甸植物正构烷烃分布特征,对采自黄河源区高寒草甸的58种植物进行了测定和分析。结果表明,单种植物中正构烷烃的碳数在C8和C40之间,含量分布范围为0.81~22.88 mg/kg,其中碳数为C27、C29、C31、C33正构烷烃的含量均较高;总正构烷烃(Ctotal)含量变化范围为35.00~78.64 mg/kg,长链正构烷烃(C25-36)含量范围为16.11~58.68 mg/kg,条叶垂头菊中总正构烷烃和长链正构烷烃含量最小,而阿尔泰狗娃花含量最大。58种植物中含量最高的正构烷烃(Cmax)分布特征为:C19有1种,C22有2种,C27有2种,C29有23种,C31有27种,C32有1种,C33有2种。植物中正构烷烃总正构烷烃碳优势指数(CPItotal)范围为1.07~4.32,CPI25-36范围为1.12~8.20,总的正构烷烃平均碳链长度(ACLtotal)范围为23.52~27.22,ACL27-33范围为28.03~31.07,蜡质指数(Pwax)变化范围为0.60~0.92。植物中长链正构烷烃丰富,奇数碳含量分布优势明显。
高寒草甸;植物;正构烷烃;气相色谱-质谱联用仪(GC-MS);黄河源区
高等植物的表皮覆盖着一层由长链有机化合物组成的蜡质。蜡质具有多种功能,它可以为植物提供物理化学保护屏障、减少非气孔水分的损失、控制温度、提供光保护、增强抗霜冻能力以及对有害物(包括害虫、真菌和细菌)攻击的抵抗力。植物蜡质是一种复杂的混合物,其成分是长链的脂肪族和环状化合物,包括碳水化合物、初级和次级醇、醛、酮、酯、脂肪酸以及三萜系化合物等[1]。植物蜡质不仅取决于生长阶段、组织类型等内源因素,也取决于光照、温度、水分等外源因素[2]。作为植物蜡质重要组成部分之一的正构烷烃在自然界植物的叶、茎、花以及果实等器官和组织的表面广泛存在[3],在海洋、湖泊、泥炭、石笋、土壤中均有分布[4-7],由于其性质稳定、难降解,且正构烷烃与对应的生物源前身有一定的结构联系或相关性[8],从而常用作分子标志物来指示生态环境特征与物源关系[9]。作为分子化石的正构烷烃,记录很多古植被及环境变化的信息,广泛应用于气候与环境变化领域、有机物质来源鉴定,为古植被恢复和重建提供信息支持[10-11]。目前植物正构烷烃也广泛应用于植物个体差异性、种群多样性和古环境演变等生态学研究[12]。不同环境中植物的正构烷烃分布有所差异,海洋植物中藻类优势正构烷烃表现出以短链为主,主要集中在C15、C17和C19,而陆地植物中的正构烷烃以长链烷烃占优势,优势正构烷烃为C23、C25、C27和C31[13]。此外,植物中正构烷烃分布还受到植物种类、组织器官、纬度和海拔等因素的影响[14]。而据报道的一些黄土和南方红土研究,对其中的正构烷烃进行抽提、分离后,试图通过分析其正构烷烃分子分布特征,来反演地质历史时期的植被类型或者其演化过程[15]。由于高寒草甸土壤中的正构烷烃记载了不同时期植被信息,通过对高寒草甸植物中正构烷烃的研究,比对土壤和植物中正构烷烃的分布特征,有利于揭示土壤正构烷烃与植物的物源关系,为高寒草甸植被演替提供证据,为植被恢复与重建提供物源信息。
1 材料与方法
1.1研究区概况
采样地位于黄河源区的青海省果洛州达日县和玛沁县(33°34′-34°28′ N,99°53′-100°29′ E),海拔3700~4100 m,年均温度为-0.5℃,年均降水量595 mm,年均蒸发量1205.9 mm,年日照时数2260 h,昼夜温差大,属较典型的高原大陆性气候。采样区内健康高寒草甸典型植被以小嵩草(Kobresiapygmaea)、矮嵩草(Kobresiahumilis)、西藏嵩草(Kobresiaschoenoides)、垂穗披碱草(Elymusnutans)和冷地早熟禾(Poacrymophila)等为优势种,退化高寒草甸典型植被以鹅绒委陵菜(Potentillaanserina)、矮火绒草(Leontopodiumnanum)、甘肃马先蒿(Pediculariskansuensis)、甘肃棘豆(Oxytropiskansuensis)、细叶亚菊(Ajaniatenuifolia)、西伯利亚蓼(Polygonumsibiricum)、密花香薷(Elsholtziadensa)、黄帚橐吾(Ligulariavirgaurea)以及铁棒锤(Aconitumpendulum)等为优势种。
1.2样品采集与制备
单种植物采集过程中,同一采样点每种植物采集5~8株,混合到一起,装入纸袋。采集后植物样品经风干、粉碎、过0.15 mm筛后,装入自封袋密封保存,供分析使用。
1.3提取方法
准确称取0.4000 g植物粉末样品置于40 mL棕色带塞的玻璃瓶中,加入5 mL二氯甲烷和甲醇的混合液(3∶1,V/V),盖好瓶塞摇匀,再将瓶置于超声波洗涤器中,超声浸提15 min,连续浸提3次,将3次提取液收集在一起,然后将提取液全部转移至填充了0.15 mm硅胶(5 g)层析柱(1 cm×10 cm)中,过夜直至提取液全部通过层析柱,然后连续2次加入10 mL正己烷进行洗脱,收集两次洗脱液,混匀,过0.45 μm滤膜后,将洗脱液氮吹浓缩至1 mL,直接上机分析(2015年4月),每个样品重复测定3次。
1.4仪器参数
GC-MS分析采用美国Thermo Fisher Scientific公司DSQII型气质联用仪(NIST 2008版谱库)。色谱条件:DB-5MS石英毛细管柱(60.00 m×0.25 mm×0.25 μm);程序升温,柱温60℃,保持1 min,4℃/min升到180℃,2℃/min升到280℃,保持20 min,20℃/min 升到300℃,持续2 min;进样量1 μL,不分流进样;进样口温度280℃;载气为高纯氦(99.999%),流速1.0 mL/min。质谱条件:电离方式为电子轰击离子源模式(EI),电子能量70 eV,离子源温度250℃,GC与MS传送杆温度280℃,扫描方式为Full Scan 模式,质量扫描范围为40~700 amu,溶剂延迟7 min。
2 结果与分析
2.1植物中短链正构烷烃分布特征分析
由表1可以看出,58种高寒草甸常见植物中短链正构烷烃(C8~C20)的含量分布范围为0.81~7.46 mg/kg,白苞筋骨草(Ajugalupulina)的C12含量最低,而臭蒿(Artemisiahedinii)的C19含量最高。其中黄帚橐吾、矮火绒草、独一味(Lamiophlomisrotata)、山生柳(Salixoritrepha)、金露梅(Potentillafruticosa)5种植物中未检测到C8正构烷烃,铁棒锤、马尿泡(Przewalskiatangutica)中未检测到C19正构烷烃。对58种植物中正构烷烃经过GC-MS分离测定后共得到短链正构烷烃有效数据749个,其中正构烷烃含量小于或等于1 mg/kg占28.70%,含量介于1~2 mg/kg占70.37%,含量大于或等于2 mg/kg为0.93%。在58种植物中,47种植物的C10、56种植物的C12、57种植物的C16、55种植物的C18正构烷烃含量小于1 mg/kg,所有检测植物其他的短链正构烷烃含量均大于1 mg/kg。
2.2不同科植物长链正构烷烃分布特征
2.2.1毛茛科植物长链正构烷烃含量从图1可知,4种毛茛科植物长链正构烷烃(C21~C40)含量分布范围:密花翠雀(Delphiniumdensiflorum)为0.96~6.40 mg/kg,铁棒锤为0.86~2.89 mg/kg,露蕊乌头(Aconitumgymnandrum)为0.92~3.87 mg/kg,瓣蕊唐松草(Thalictrumpetaloideum)为0.96~7.22 mg/kg。4种植物中偶数碳长链正构烷烃(C22~C40)除C22、C34外含量差别不大,铁棒锤的 C22、C34含量要显著高于其他3种植物;奇数碳长链正构烷烃(C21~C35)中,密花翠雀和铁棒锤C21、C23和C25含量相近,而露蕊乌头和瓣蕊唐松草C21、C23和C25含量相近,但前者要高于后者。C27的含量大小为露蕊乌头>密花翠雀>瓣蕊唐松草>铁棒锤;C29、C31、C33在4种植物中有较高的含量,且C29、C31、C33的含量大小关系均为瓣蕊唐松草>密花翠雀>露蕊乌头>铁棒锤。
2.2.2唇形科植物长链正构烷烃含量由图2可得,唇形科4种植物长链正构烷烃(C21~C40)含量分布范围:白苞筋骨草为0.95~18.65 mg/kg,黄花粘毛鼠尾草(Salviaroborowskii)为0.96~3.42 mg/kg(其中未检测到C34),密花香薷为0.95~4.60 mg/kg,独一味为0.86~3.95 mg/kg。4种植物中偶数碳正构烷烃(C22~C40)含量基本相同,奇数碳正构烷烃(C21~C35)中C23~C27含量基本一致,黄花粘毛鼠尾草C21要明显高于其他3种植物。4种植物中C29、C31、C33的含量较高,密花香薷的C29含量高,而白苞筋骨草的C31、C33和C35含量要高于黄花粘毛鼠尾草、密花香薷、独一味,且白苞筋骨草的C31正构烷烃在这4种植物中含量最高。
2.2.3蔷薇科植物长链正构烷烃含量从图3可以看出,蔷薇科4种植物长链正构烷烃(C21~C40)含量分布范围:金露梅为0.96~9.01 mg/kg,鹅绒委陵菜为0.96~5.36 mg/kg,高山绣线菊(Spiraeaalpina)为0.97~11.74 mg/kg,二裂委陵菜(Potentillabifurca)为0.97~11.55 mg/kg。4种植物中偶数碳长链正构烷烃除C26、C28和C34外含量一致,鹅绒委陵菜的 C26、C28和C34含量比其他3种植物高;奇数碳长链正构烷烃(C21~C35)中C21、C23和C35含量基本相同,鹅绒委陵菜和高山绣线菊的C25含量一致,金露梅和二裂委陵菜的C25含量相同,但前者要高于后者。C27的含量大小为高山绣线菊>金露梅>二裂委陵菜>鹅绒委陵菜,4种植物中C29、C31和C33的含量较高,高山绣线菊的C29含量较其他植物高,C31、C35正构烷烃的含量大小为二裂委陵菜>金露梅>鹅绒委陵菜>高山绣线菊。
2.2.4玄参科植物长链正构烷烃含量由图4可知,玄参科2种植物中长链正构烷烃(C21~C40)含量分布范围:肉果草(Lanceatibetica)为0.88~2.58 mg/kg,甘肃马先蒿为0.92~5.71 mg/kg。2种植物中偶数碳长链正构烷烃(C22~C40)除C32外含量一致,肉果草的C32含量要高于甘肃马先蒿,奇数碳正构烷烃(C21~C35)中C21和C35含量基本相同,2种植物中C25~C33的含量较高,且甘肃马先蒿C25~C33的含量高于肉果草。

表1 植物中短链正构烷烃含量Table 1 The content of short chain n-alkanes in plants mg/kg

续表1 Continued
注:“-”为未检测到。
Note: “-” not detected.

图1 毛茛科植物长链正构烷烃含量Fig.1 The content of long chain n-alkanes in Ranunculaceae plants

图2 唇形科植物长链正构烷烃含量Fig.2 The content of long chain n-alkanes in Labiatae plants

图3 蔷薇科植物长链正构烷烃含量Fig.3 The content of long chain n-alkanes in Rosaceae plants
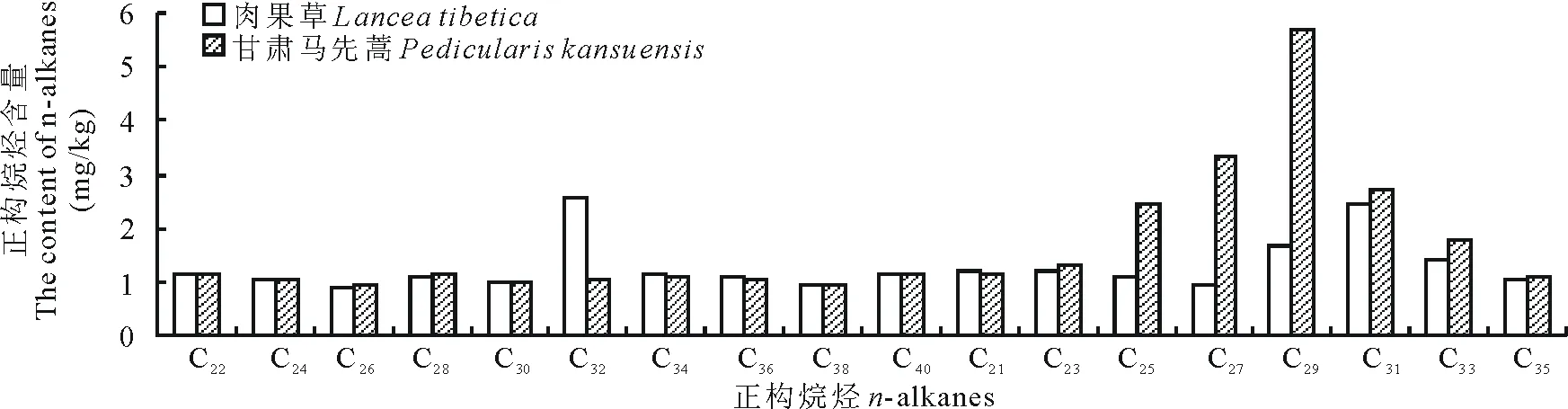
图4 玄参科植物长链正构烷烃含量Fig.4 The content of long chain n-alkanes in Scrophulariaceae plants
2.2.5豆科植物长链正构烷烃含量由图5可得,豆科2种植物中长链正构烷烃(C21~C40)含量分布范围:甘肃棘豆为0.86~3.08 mg/kg,黄花棘豆(Oxytropisochrocephala)为0.96~5.09 mg/kg。2种植物中偶数碳长链正构烷烃(C22~C40)含量基本相同,其中甘肃棘豆C22的含量高于黄花棘豆,黄花棘豆C26的含量高于甘肃棘豆;奇数碳正构烷烃(C21~C35)中C21、C23和C35含量基本相同,2种植物中C25~C33的含量较高,且黄花棘豆C25~C33的含量高于甘肃棘豆。
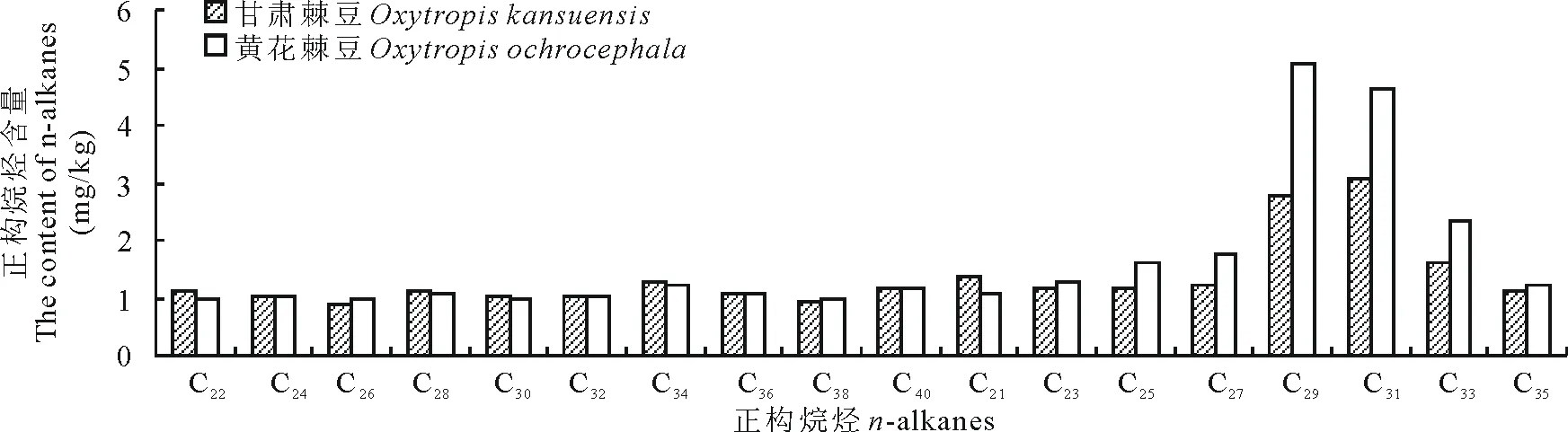
图5 豆科植物中长链正构烷烃含量Fig.5 The content of long chain n-alkanes in Leguminous plants
2.2.6龙胆科植物长链正构烷烃含量从图6看出,龙胆科4种植物长链正构烷烃(C21~C40)含量分布范围:达乌里秦艽(Gentianadahurica)为0.96~3.12 mg/kg,线叶龙胆(Gentianafarreri)为0.96~4.99 mg/kg,湿生扁蕾(Gentianopsispaludosa)为0.96~10.74 mg/kg,麻花艽(Gentianastraminea)为0.86~4.51 mg/kg。4种植物中偶数碳长链正构烷烃(C22~C40)除C28外含量基本一致,达乌里秦艽、麻花艽、线叶龙胆的C28含量高于其他偶数碳长链烷烃;奇数碳正构烷烃(C21~C35)中C21、C23和C35含量基本相同,4种植物中C25~C33的含量较高,其中湿生扁蕾中C27的含量最高。

图6 龙胆科植物长链正构烷烃含量Fig.6 The content of long chain n-alkanes in Gentianaceae plants
2.2.7菊科植物长链正构烷烃含量由表2可知,13种菊科植物长链正构烷烃(C21~C40)含量范围为:0.86~22.88 mg/kg,其中夏河紫菀(Asteryunnanensis)中C26含量最低,阿尔泰狗娃花(Heteropappusaltaicus)中C31含量最高,大籽蒿(Artemisiasieversiana)未检测到C34。13种菊科植物中共检测到18种长链正构烷烃有效数据233个,其中正构烷烃含量小于2 mg/kg占81.12%,而含量大于或等于2 mg/kg为18.88%。臭蒿(Artemisiahedinii)中含量大于2 mg/kg正构烷烃有8种,分别为C23、C30、C28、C25、C31、C26、C27和C29(含量依次增大)。黄帚橐吾(Ligulariavirgaurea)中含量大于2 mg/kg正构烷烃有6种,分别为C31、C33、C23、C29、C27和C22(含量依次增大)。
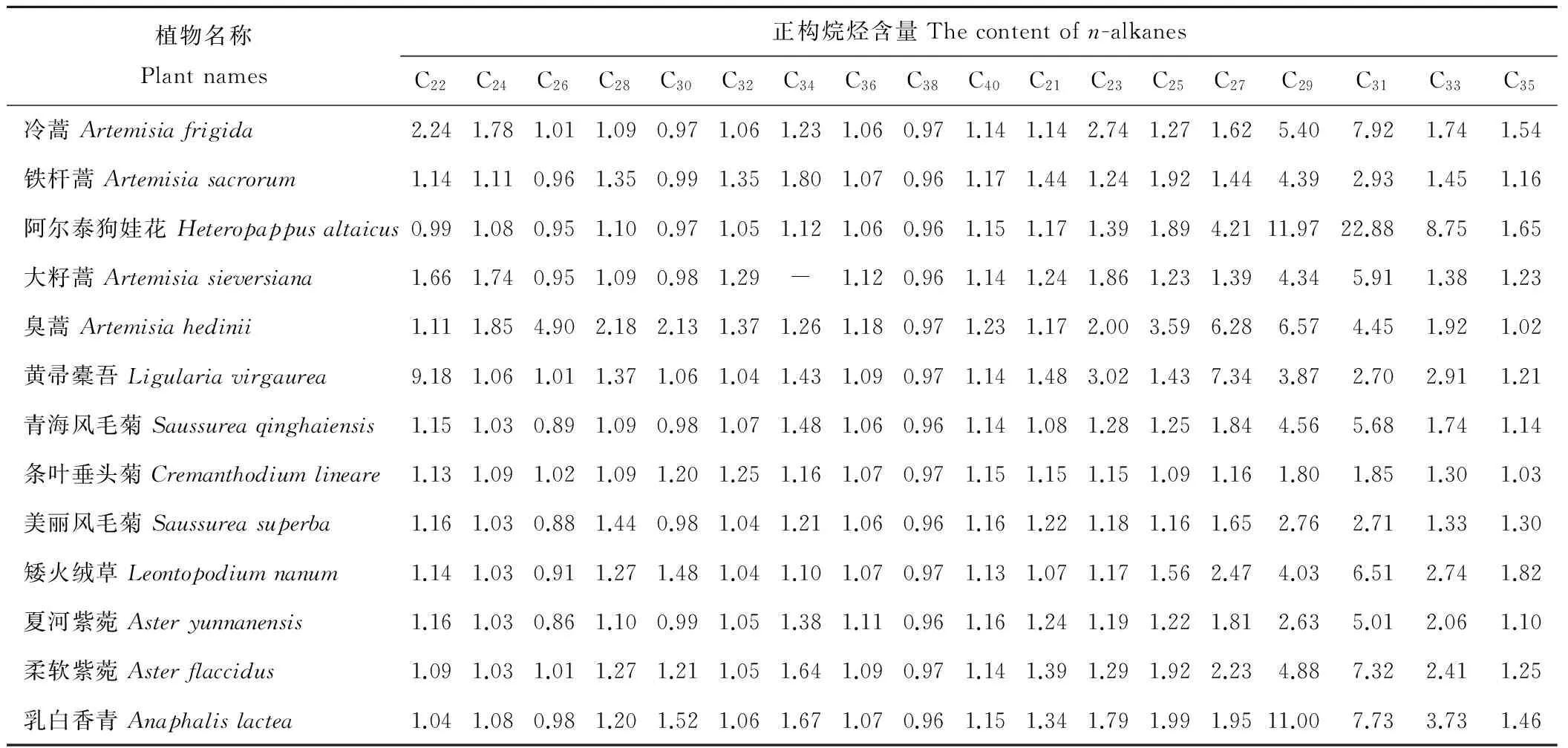
表2 菊科植物长链正构烷烃的含量Table 2 The content of long chain n-alkanes in Compositae plants mg/kg
注:“-”为未检测到。
Note: “-” not detected.
2.2.8禾本科植物长链正构烷烃含量由图7可得,禾本科6种植物长链正构烷烃(C21~C40)含量分布范围:紫花针茅(Stipapurpurea)为0.95~11.87 mg/kg,中华羊茅(Festucasinensis) 为0.96~6.60 mg/kg,垂穗披碱草(Elymusnutans)为0.95~6.01 mg/kg,冷地早熟禾(Poacrymophila)为0.96~5.28 mg/kg,梭罗草(Kengyiliathoroldiana)为0.97~5.41 mg/kg,麦宾草(Elymustangutorum)为0.95~5.71 mg/kg。6种植物中偶数碳长链正构烷烃(C22~C40)除C28、C30外含量基本一致,梭罗草中C28、C30含量要高于其他5种植物;奇数碳正构烷烃(C21~C35)中C21~C25和C35含量基本相同,6种植物中C27~C33的含量较高,其中紫花针茅C31的含量最高。
2.2.9莎草科植物长链正构烷烃含量由图8可知,莎草科6种植物长链正构烷烃(C21~C40)含量分布范围:黑褐苔草(Carexastrofusca)为0.96~2.73 mg/kg,青藏苔草(Carexmoorcroftii)为0.97~3.36 mg/kg,西藏嵩草(Kobresiaschoenoides)为0.96~4.72 mg/kg,矮嵩草(Kobresiahumulis)为0.96~3.40 mg/kg,双柱头藨草(Scirpusdistigmaticus)为0.95~11.20 mg/kg,小嵩草(Kobresiapygmaea)为0.97~9.22 mg/kg。6种植物中偶数碳长链正构烷烃(C22~C40)除C34外含量基本一致,小嵩草中C34含量要高于其他5种植物;奇数碳正构烷烃(C21~C35)中C21~C25和C35含量基本相同,6种植物中C27~C33的含量较高,其中双柱头藨草C29的含量最高,正构烷烃C29的含量大小关系为双柱头藨草>小嵩草>矮嵩草>西藏嵩草>黑褐苔草>青藏苔草,正构烷烃C31的含量大小关系为小嵩草>双柱头藨草>西藏嵩草>青藏苔草>矮嵩草>黑褐苔草。

图8 莎草科植物长链正构烷烃含量Fig.8 The content of long chain n-alkanes in Cyperaceae plants
2.2.10其他科植物长链正构烷烃含量由表3可以得出,13科13种植物中长链正构烷烃(C21~C40)含量变化范围为0.86~11.54 mg/kg,海乳草(Glauxmaritime)、拉拉藤(Galiumaparine)、甘青大戟(Euphorbiamicractina)、圆萼刺参(Morinachinensis)中C26含量最低,羌活(Notopterygiumincisum)中C27含量最高,其中马尿泡(Przewalskiatangutica)、羌活中未检测到C38。13科13种植物中共检测到18种长链正构烷烃有效数据232个,其中长链正构烷烃含量小于2 mg/kg占86.21%,而含量大于2 mg/kg为13.79%。甘青大戟中大于2 mg/kg的正构烷烃有6种,分别为C35、C27、C33、C34、C29和C31(含量依次增大)。
2.3单种植物中正构烷烃的Cmax、CPI、ACL和Pwax分布特征
从表4可知,单种植物中总正构烷烃(Ctotal)含量变化范围为35.00~78.64 mg/kg,长链正构烷烃(C25-36)含量范围为16.11~58.68 mg/kg,条叶垂头菊中总正构烷烃和长链正构烷烃含量最小,阿尔泰狗娃花中总正构烷烃和长链正构烷烃含量最大。58种植物中含量最高的正构烷烃(Cmax)及植物种类分布特征为:C19正构烷烃有1种,C22正构烷烃有2种,C27正构烷烃有2种,C29正构烷烃有23种,C31正构烷烃有27种,C32正构烷烃有1种,C33正构烷烃有2种。总正构烷烃碳优势指数(CPItotal)范围为1.07~4.32,长链正构烷烃碳优势指数(CPI25-36)范围为1.12~8.20,植物中的正构烷烃奇数碳含量分布优势明显。植物中总的正构烷烃平均碳链长度(ACLtotal)范围为23.52~27.22,长链正构烷烃平均碳链长度(ACL27-33)范围为28.03~31.07。植物中蜡质指数(Pwax)变化范围为0.60~0.92,Pwax值最大的植物为阿尔泰狗娃花,Pwax值最小的植物为铁棒锤。
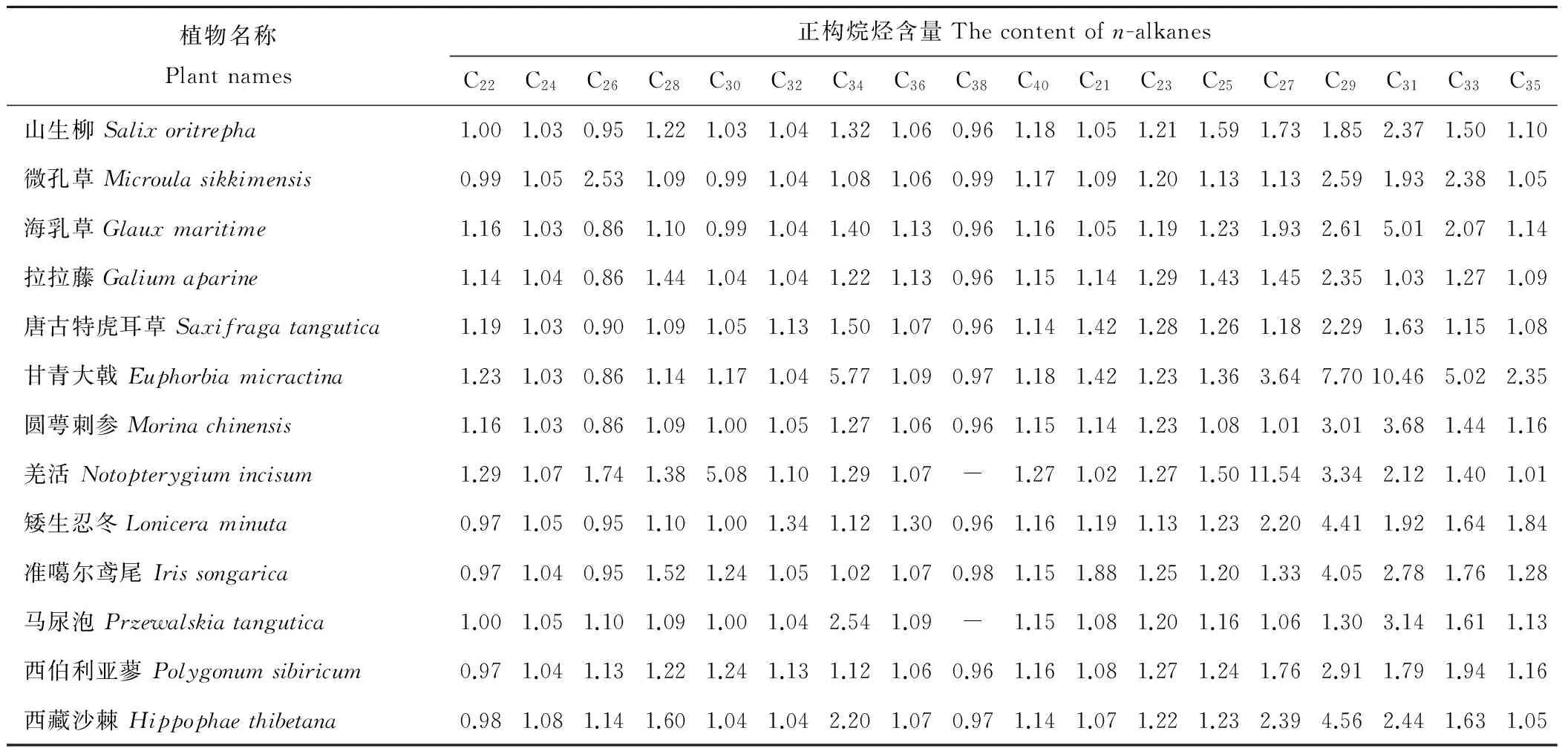
表3 其他科植物长链正构烷烃的含量Table 3 The content of long chain n-alkanes in other family plants mg/kg
注:“-”为未检测到。
Note: “-” not detected.
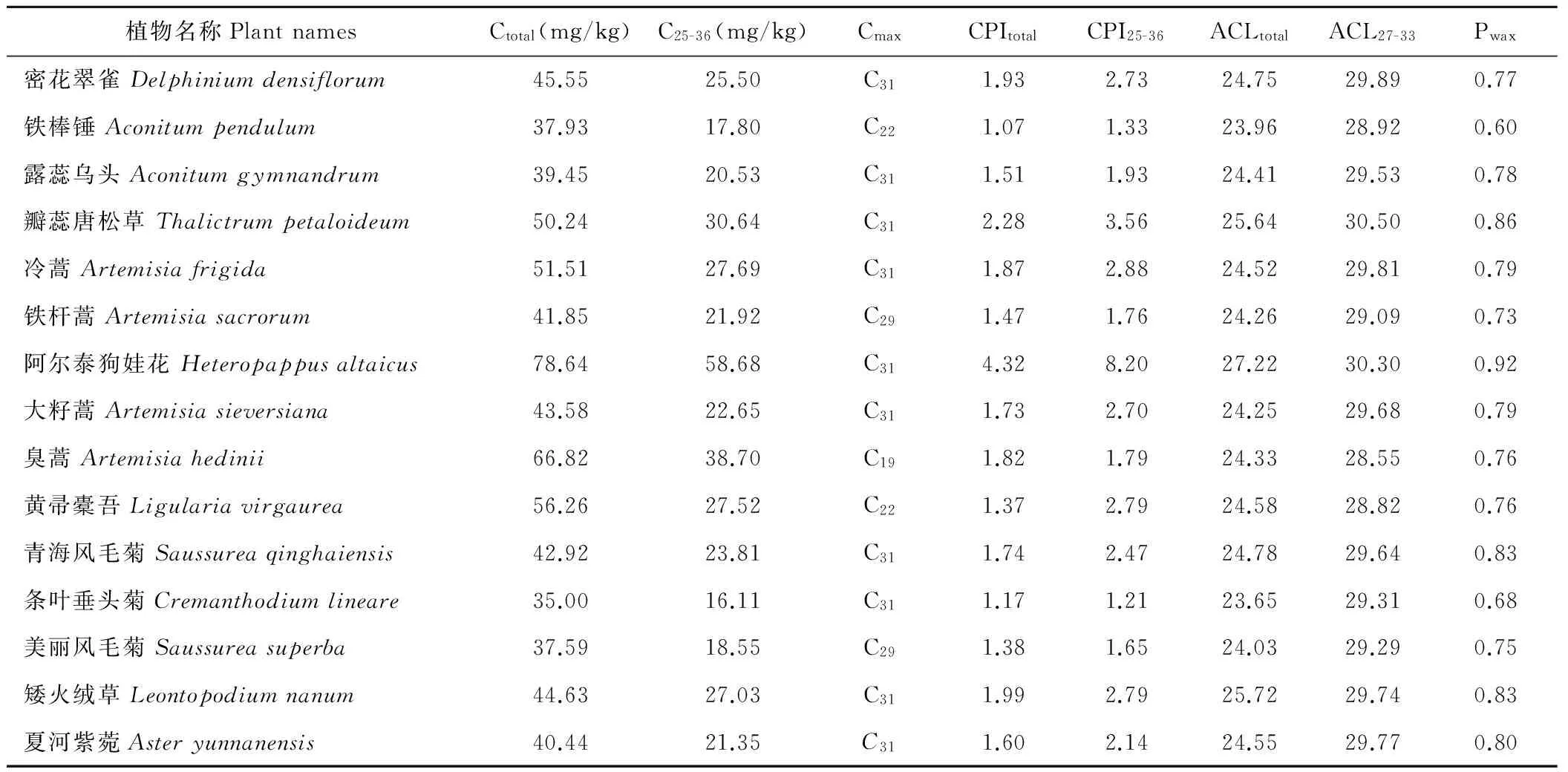
表4 单种植物中正构烷烃的Cmax、CPI、ACL、Pwax分布特征Table 4 The distribution characteristics of Cmax, CPI, ACL and Pwax of n-alkanes in individual plant

续表4 Continued
Ctotal=∑[C8-40];C25-36=∑[C25-36];CPItotal=odd∑[C9-35]/even∑[C10-36];CPI25-36=odd∑[C25-35]/even∑[C26-36];ACLtotal=∑[n×Cn]/∑[Cn],n=8-40;ACL27-33=(27×C27+29×C29+31×C31+33×C33)/(C27+C29+C31+C33);Pwax=(C27+C29+C31)/(C23+C25+C27+C29+C31).
3 讨论
植物中正构烷烃的生成需要经过很复杂的过程,受诸多因素的影响,生长在同一地区同科植物中各长链正构烷烃含量有着明显差异。58种植物中各种正构烷烃奇偶优势明显,且C27、C29、C31含量较高,这和段毅等[14]、孙丰瑞等[16]研究结果一致。正烷烃的分布与植物类型有关,木本植物中C27和C29占优势,而草本植物C31更占优势[17],因此土壤中C27或者C29为主峰的长链正构烷烃可以用来指示来源植被为森林植被,以C31为主峰的长链正构烷烃指示来源植被为草原植被[15]。但是在高寒草甸植物中,不管是草本还是木本植物中的正构烷烃,大部分是C31占优势,这可能和这些植物生长的环境有关。Schefuβ等[18]提出,植物的烷烃组成在植物适应环境的过程中起了重要的作用,较高的烷烃含量可能有助于植物体抵抗寒冷。不同季节采集的样品中正构烷烃含量有明显差异,在5和8月正构烷烃较低,且没有明显的变化,而到11月份却有明显增大的趋势[1],这是不同研究者对同一地区同一种植物中正构烷烃含量不一致的根本原因。植物中正构烷烃的CPI随着植物不同而不同,这与崔景伟等[1]研究结果相似。植物中正构烷烃的平均碳链长度ACL值与植物的类型及其生长地区环境有关[14],因此通过分析土壤中正构烷烃ACL值来推测土壤形成时植被类型以及环境要素的变化趋势。植物中正构烷烃的Pwax值能反映植物生长环境中湿润程度[4],通过对植物中Pwax值分析,Pwax值小的植物适宜在湿润环境中生长,反之Pwax值大的植物适宜于干旱环境。地质基质土壤、沉积物中正构烷烃的Pwax值能反映土壤、沉积物形成过程古气候水分丰沛程度变迁规律。
4 结论
1)单种植物中正构烷烃含量分布范围为0.81~22.88 mg/kg,其中C27、C29、C31、C33的含量均较高。58种植物中Cmax分布特征为:C19正构烷烃有1种,C22正构烷烃有2种,C27正构烷烃有2种,C29正构烷烃有23种,C31正构烷烃有27种,C32正构烷烃有1种,C33正构烷烃有2种。
2)植物中正构烷烃CPItotal范围为1.07~4.32,CPI25-36范围为1.12~8.20,植物中的正构烷烃奇数碳含量分布优势明显。ACLtotal范围为23.52~27.22,ACL27-33范围为28.03~31.07。Pwax值变化范围为0.60~0.92,Pwax值最大的植物为阿尔泰狗娃花,Pwax值最小的植物为铁棒锤。
References:
[1]Cui J W, Huang J H, Xie S C.Seasonal variation ofn-alkanes and olefins in modern plant leaves of Qingjiang in Hubei. Chinese Science Bulletin, 2008, 53(11): 1318-1323.
[2]Li J J, Huang J H, Xie S C. Plant wax and its response to environmental conditions: an overview. Acta Ecologica Sinica, 2011, 31(2): 0565-0574.
[3]Zhong Y X, He J, Xue Q.Distribution ofn-alkane in modern ligneous plants from the western loess plateau. Journal of Lanzhou University (Natural Sciences), 2011, 47(2): 32-36.
[4]Pu Y, Zhang H C, Wang Y L,etal. Climatic and environmental implications fromn-alkanes in glacially eroded lake sediments in Tibetan Plateau: An example from Ximen Co. Chinese Science Bulletin, 2011, 56(14): 1503-1510.
[5]Martina G, Yakov K, Guido L B W.Differentiation of plant derived organic matter in soil, loess and rhizoliths based onn-alkane molecular proxies. Biogeochemistry, 2013, 112: 23-40.
[6]Manuel S A, Riaz A, Marta C A,etal. Changes in the chemical composition of soil organic matter over time in the presence and absence of living roots: a pyrolysis GC/MS study. Plant and Soil, 2015, 391: 161-177.
[7]Fatma A, Monem K, Mohamed D,etal. Even-numberedn-alkanes/n-alkenes predominance in surface sediments of Gabes Gulf in Tunisia. Environment Earth Science, 2010, 61: 1-10.
[8]Chang C, Wang S H, Zhu C,etal. Biomarker geochemistry of marine organic matter in the Hushan and Chaohu areas, Lower Yangtze region. Chinese Journal of Geochemistry, 2011, 30: 145-152.
[9]Bai Y, Fang X M, Wang Y L,etal. Branched alkanes with quaternary carbon atoms in Chinese soils: Potential environmental implications.Chinese Science Bulletin, 2006, 51(9): 1115-1122.
[10]Kristen I, Wilkes H, Vieth A,etal. Biomarker and stable carbon isotope analyses of sedimentary organic matter from Lake Tswaing: evidence for deglacial wetness and early Holocene drought from South Africa. Journal of Paleolimnology, 2010, 44: 143-160.
[11]Gerd L, Ralf W.n-Alkanes as indicators of natural and anthropogenic organic matter sources in the Siak River and its estuary, E Sumatra, Indonesia. Bulletin of Environmental Contamination and Toxicology, 2009, 83: 403-409.
[12]He Y J, Guo N, Gao J H,etal.The response of waxn-alkanes inFicustikoualeaf to growing environments. Acta Agrestia Sinica, 2015, 23(1): 114-119.
[13]Punyu V R, Harji R R, Bhosle N B,etal.n-Alkanes in surficial sediments of Visakhapatnam harbour, east coast of India. Journal of Earth System Science, 2013, 122(2): 467-477.
[14]Duan Y, Wu B X, Xu L,etal. The compositions ofn-alkanes and their isotopic in different latitudes plants. Acta Geologica Sinica, 2011, 85(2): 262-271.
[15]Rao Z G, Wu Y, Zhu Z Y,etal. Is the maximum carbon number of long-chainn-alkanes an indicator of grassland or forest: Evidence from surface soils and modern plants. Chinese Science Bulletin, 2011, 56(10): 774-780.
[16]Sun F R, Li Y M, Chen J F,etal. Odd-even preference ofn-alkanes in plateau plants and its environmental indication significance. Global Geology, 2011, 30(3): 469-473.
[17]Long L Q, Fang X M, Miao Y F,etal. Northern Tibetan Plateau cooling and aridification linked to Cenozoic global cooling: Evidence fromn-alkane distributions of Paleogene sedimentary sequences in the Xining Basin. Chinese Science Bulletin, 2011, 56(15): 1569-1578.
[18]Schefuβ E, Ratmeyer V, Stuut J W,etal. Carbon isotope analyses ofn-alkanes in dust from the lower atmosphere over the central eastern Atlantic. Journal of The Geochemical Society and The Meteoritical Society, 2003, 67(10): 1757-1767.
[1]崔景伟, 黄俊华, 谢树成.湖北清江现代植物叶片正构烷烃和烯烃的季节性变化.科学通报, 2008, 53(11): 1318-1323.
[2]李婧婧, 黄俊华, 谢树成.植物蜡质及其与环境的关系.生态学报, 2011, 31(2): 0565-0574.
[3]钟艳霞, 贺婧, 薛骞. 黄土高原西部地区现代木本植被叶片正构烷烃分布模式. 兰州大学学报(自然科学版), 2011, 47(2): 32-36.
[12]和玉吉, 郭娜, 高建花, 等.生长环境对地瓜藤叶表皮烷烃含量及组成的影响.草地学报, 2015, 23(1): 114-119.
[14]段毅, 吴保祥, 徐丽, 等.不同纬度地区植物中正构烷烃及其同位素组成.地质学报, 2011, 85(2): 262-271.
[15]饶志国, 吴翼, 朱照宇, 等.长链正构烷烃主峰碳数作为判别草本和木本植物指标的讨论: 来自表土和现代植物的证据.科学通报, 2011, 56(10): 774-780.
[16]孙丰瑞, 李玉梅, 陈践发, 等.高原植物正构烷烃的奇偶优势及其环境指示意义.世界地质, 2011, 30(3): 469-473.
*Characterization of plantn-alkanes in alpine meadow
DUAN Zhong-Hua1,2,3, QUAN Xiao-Long1, QIAO You-Ming1*, PEI Hai-Kun3, HE Gui-Fang1
1.College of Eco-Environmental Engineering, Qinghai University, Xining 810016, China; 2.Stake Key Laboratory of Plateau Ecology and Agriculture, Qinghai University, Xining 810016, China; 3.Instrumental Analysis Center of Qinghai University, Xining 810016, China
Alpine meadow plants were collected and analysed to characterize then-alkanes present. The results showed that the carbon number ofn-alkanes of individual plants varied from C8to C40andn-alkanes content ranged from 0.8 to 22.9 mg/kg. The C27, C29, C31and C33n-alkanes contents were greater than that of remainingn-alkanes; totaln-alkane (Ctotal) ranged from 35.0 to 78.6 mg/kg, while the long chainn-alkane (C25-36) content ranged from 16.1 to 58.7 mg/kg. The totaln-alkane and long chainn-alkane content was highest inHeteropappusaltaicusand least inCremanthodiumlineare. Analysis of then-alkane distribution of the Cmax among 58 plants sampled revealed one species with a Cmaxof C19, two species with C22, two species with C27, 23 species with C29, 27 species with C31, one species with C32and two species with C33. The CPItotalranged from 1.1 to 4.3, and the CPI25-36from 1.1 to 8.2. The ACLtotalranged from 23.5 to 27.2, and the ACL27-33from 28.0 to 31.1. The Pmaxranged from 0.6 to 0.9. The sampled plants contained abundant long-chainn-alkanes with a significant odd carbon number preference.
alpine meadow; plants;n-alkanes; gas chromatography-mass spectrometer (GC-MS); headwater region of the Yellow River
10.11686/cyxb2016036
http://cyxb.lzu.edu.cn
2016-01-21;改回日期:2016-03-08
国家自然基金项目(31260573),青海省科技厅国际合作项目(2012-H-806),教育部创新团队(IRT13074)和教育部美大项目(2014)资助。
段中华(1980-),男,湖南邵阳人,副教授,在读博士。E-mail:zhonghuaduan@126.com
Corresponding author. E-mail:ymqiao@aliyun.com
段s中华,全小龙,乔有明,裴海昆,何桂芳. 高寒草甸植物正构烷烃特征分析. 草业学报, 2016, 25(6): 136-147.
DUAN Zhong-Hua, QUAN Xiao-Long, QIAO You-Ming, PEI Hai-Kun, HE Gui-Fang. Characterization of plantn-alkanes in alpine meadow. Acta Prataculturae Sinica, 2016, 25(6): 136-147.
猜你喜欢
润滑油(2022年4期)2022-09-16
纺织标准与质量(2022年3期)2022-08-10
金桥(2022年2期)2022-03-02
昆明医科大学学报(2021年4期)2021-07-23
Plasma Science and Technology(2021年3期)2021-03-22
塑料助剂(2019年3期)2019-07-24
中国环境科学(2019年5期)2019-06-06
浙江大学学报(工学版)(2016年11期)2016-06-05
中国塑料(2015年10期)2015-10-14
医学研究杂志(2015年4期)2015-06-10
