
大麦RIL群体内不同类型苗粉和籽粒元素的差异
2016-09-16 03:57曾亚文汪禄祥杨晓梦杨加珍普晓英杜丽娟杨树明
中国农业科学 2016年15期
曾亚文,汪禄祥,杨晓梦,杨加珍,杜 娟,普晓英,杜丽娟,杨树明,肖 亚,杨 涛
(1云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室,昆明650205;2云南省农业科学院质量标准与检测技术研究所,昆明650223;3西藏自治区日喀则市农业科学研究所,西藏日喀则 857000)
大麦RIL群体内不同类型苗粉和籽粒元素的差异
曾亚文1,汪禄祥2,杨晓梦1,杨加珍1,杜 娟1,普晓英1,杜丽娟2,杨树明1,肖 亚3,杨 涛1
(1云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室,昆明650205;2云南省农业科学院质量标准与检测技术研究所,昆明650223;3西藏自治区日喀则市农业科学研究所,西藏日喀则 857000)
【目的】研究大麦重组近交系群体(RIL)的变异类型分类与苗粉(麦苗)及籽粒的矿质元素含量及其遗传变异,揭示栽培大麦苗粉及籽粒矿质元素含量的遗传变异。【方法】采用栽培大麦变种分类方法,按照穗芒颜色、护颖宽窄、侧小穗有无内外稃、芒性状、粒色、穗密度及籽粒稃状对 194个紫光芒裸二棱×Schooner大麦RIL株系及其亲本进行变异类型分类。用ICP-AES测定194个紫光芒裸二棱×Schooner大麦RIL株系及其亲本的籽粒和苗粉中9种矿质元素(P、Zn、Fe、Mn、Mg、Ca、Cu、Na和K)含量。【结果】从Violiglabrum × Erectum培育的194个RIL株系及其亲本划分为Nutans、Hypianthinum、Lanthonudum、Violiglabrum、Medicum、Nudum、Erectum、Violimedicum、Neogenes、Glabrinudidubium、Jachanum、Luchuanicum和Violierectum 共13个变异类型,变异分类性状基因型间差异显著。籽粒矿质元素含量变异类型间差异显著(1.9—6.4倍),而苗粉差异较小(1.2—2.1倍)。前者籽粒元素变种类型最高倍数差异依次为 Cu(Glabrinudidubium/ Violierectum=6.4)>Mn(Violimedicum/Violierectum=3.8)>Fe(Neogenes/Glabrinudidubium=2.9)>K (Medicum/Violierectum=2.3)>Zn(Violiglabrum/Jachanum=2.2)>P(Violimedicum/Jachanum=2.1)=Ca (Medicum/Violierectum=2.1)>Mg(Violimedicum/Jachanum = 1.9)= Na(Violimedicum/Violierectum= 1.9),后者苗粉元素变种类型最高倍数差异依次为 Na(Violierectum/Luchuanicum=2.1)>Fe (Neogenes/Erectum=1.5)>Ca(Jachanum/Luchuanicum=1.4)=Cu(Violiglabrum/Hypianthinum=1.4)>Zn (Lanthonudum/Violimedicum=1.3)=Mg(Nudum/Luchuanicum=1.3)=K(Violierectum/Hypianthinum=1.3)>P(Luchuanicum/Glabrinudidubium=1.2)=Mn(Nudum/Erectum =1.2),而籽粒及苗粉中P、Zn、Mn、Mg、Ca 和K含量变种类型间差异显著或极显著。7种苗粉与籽粒元素比为1.1—51.4倍,13个变种类型苗粉与籽粒元素比值变种类型最高倍数差异依次为Na(Violierectum 51.4)>Ca(Violierectum 19.9)>K(Violierectum 15.4)>Fe (Glabrinudidubium 8.8)>Mg(Jachanum 3.4)>Mn(Violierectum 3.2)>Zn(Jachanum 2.9)>P (Jachanum 2.7)>Cu(Violierectum 2.1)。根据RIL及其亲本的籽粒、苗粉9种矿质元素含量或苗粉/籽粒元素含量比值均可将13个变种类型划分为4个类群,群间差异明显而群内差异不大。绝大多数变种类型的籽粒和苗粉矿质元素含量与原始的云南地方品种紫光芒裸二棱的含量相近。【结论】大麦籽粒和苗粉的矿质元素含量在变种类型间差异显著,而在变种类型内差异不显著,籽粒矿质元素含量差异显著而苗粉含量差异不显著,矿质元素在栽培大麦变种类型驯化中起重要作用。
大麦;ICP-AES;矿质元素;籽粒;苗粉
0 引言
【研究意义】随着生活水平的提高,人们的主食也由糙米糙面转变为精米精面,饮食中矿质元素(尤其是Ca、Fe和Zn)的缺乏成为全球的健康问题[1]。大麦不仅是谷物中快速消化淀粉含量与血糖生成指数最低、多酚和β-葡聚糖含量最高及其防治心血管疾病功效显著[2-3]的功能食品,也是谷物中Ca、Fe、Zn和K元素含量最高及其解决人类矿质元素缺乏的关键食品。因此,研究大麦重组近交系群体(RIL)不同变异类型的苗粉(麦苗)及籽粒矿质元素及其遗传变异,对功能大麦育种及其功能食品研发十分重要。【前人研究进展】埃塞俄比亚、中国(西藏、云南)和近东是大麦起源中心。徐庭文[4]是中国栽培大麦变种分类研究的奠基人,1982年编写了栽培大麦变种检索表。其后王建林等[5]明确了西藏高原拥有栽培大麦601个变种类型,西藏栽培大麦变种是本土分化形成的。POETS等[6]报道大麦野生种群对原始地方品种适应生态环境变化的贡献较大。ZENG等[7]已绘制出适应高原气候具有36 151蛋白质编码基因的3.89 Gb西藏青稞草图集。埃塞俄比亚是国际公认的人类和大麦起源中心及其抗癌等慢性病的协同进化基地[8-9];基于分子标记揭示了西藏周边地区及近东新月沃土地带是栽培栽培大麦的驯化中心[10-12]及藏族抗癌青稞食品的协同进化基地[8]。大麦富含半胱氨酸的金属硫蛋白(MT),在细胞内Zn和Cu动态平衡中发挥作用,MT3在金属动态平衡中起关键作用,而MT4是籽粒发育及成熟中起存储Zn功能而排除Cd的作用[13]。低植酸(Hvlpa1)决定了大麦籽粒胚乳的总磷含量[14]。大麦苗粉及籽粒矿质元素的研究对研发大麦功能食品具有较高的实用价值。大麦苗粉蛋白质含量(26.8%)是大麦籽粒(14.5%)的1.9倍[15]以及大麦苗粉K和Ca含量高具有改善睡眠的作用[16]。因此,大麦的变种分类、驯化及营养功能研究中国已走在世界前列。【本研究切入点】目前,国内外基于分子标记和形态性状(尤其是变种分类性状),揭示了西藏是中国栽培大麦的驯化中心,但关于大麦变种类型间籽粒及苗粉中的矿质元素差异国内外鲜见报道,大麦籽粒及其苗粉元素对人类健康影响知之甚少。【拟解决的关键问题】本研究采用ICP-AES法测定大麦亲本和194个RIL群体株系的苗粉及籽粒中的9种矿质元素,按栽培大麦变种分类方法进行RIL的变种类型划分及聚类分析,揭示大麦变种类型间的苗粉及其籽粒矿质元素差异及其遗传变异,为培育功能大麦新品种和研发大麦功能食品提供参考。
1 材料与方法
1.1材料种植及变种分类
目前,紫光芒裸二棱是世界上大麦籽粒蛋白质含量最高和成熟时全株紫色的二棱裸大麦,为云南特有的Violiglabrum新变种。Schooner是中国进口量最大的优质啤酒大麦和成熟时全株黄色的二棱皮大麦,为Erectum变种。2012年秋季,云南省玉溪市研和镇砂壤土蔬菜区种植亲本(紫光芒裸二棱和Schooner)及其194个F8代RIL株系,采用灌排水较好和土壤肥力均匀的田块,设置2个重复,行长2 m,每行100粒,行距0.30 m,顺序排列。按当地常规农艺措施管理,拔节期割苗晒干成大麦草,麦田浇水施肥再生大麦除株高降低15 cm外其余性状发育正常。根据棱型、穗芒颜色、护颖宽窄、侧小穗有无内外稃、芒性状、粒色、穗密度及籽粒稃状,对Violiglabrum×Erectum杂交培育的194个RIL株系及其亲本进行变种类型分类[4,17]。
1.2矿质元素的测定
亲本及194个RIL株系各取10 g种子和大麦苗干草,用打粉机打成粉末状低温干燥贮存;准确称取0.50 g置于0.1 L烧杯中,加入5 mL硝酸和1 mL高氯酸,电热板上加热消解至溶液清亮,蒸至近干,加入1∶1盐酸5 mL溶解残渣,转入50 mL容量瓶定容待测;样品经湿法消解后,用盐酸溶解残渣,使待测元素转变为无机离子态。每个样品重复3次,同时进行样品空白处理,采用 ICP-AES测定矿质元素含量。用SPSS16.0统计软件进行数据统计及聚类分析。
1.3方法的精密度及回收率试验
按检出限低、灵敏度高及干扰元素少等原则选择元素分析线。通过加标回收试验验证方法的可靠性,添加0.100 μg·mL-1的标准溶液,9种元素的回收率在93.5%—110.2%,12次平行测定RSD为0.8%—4.6%,说明ICP-AES测定方法具有较高的准确度及精密度,符合分析要求(表1)。
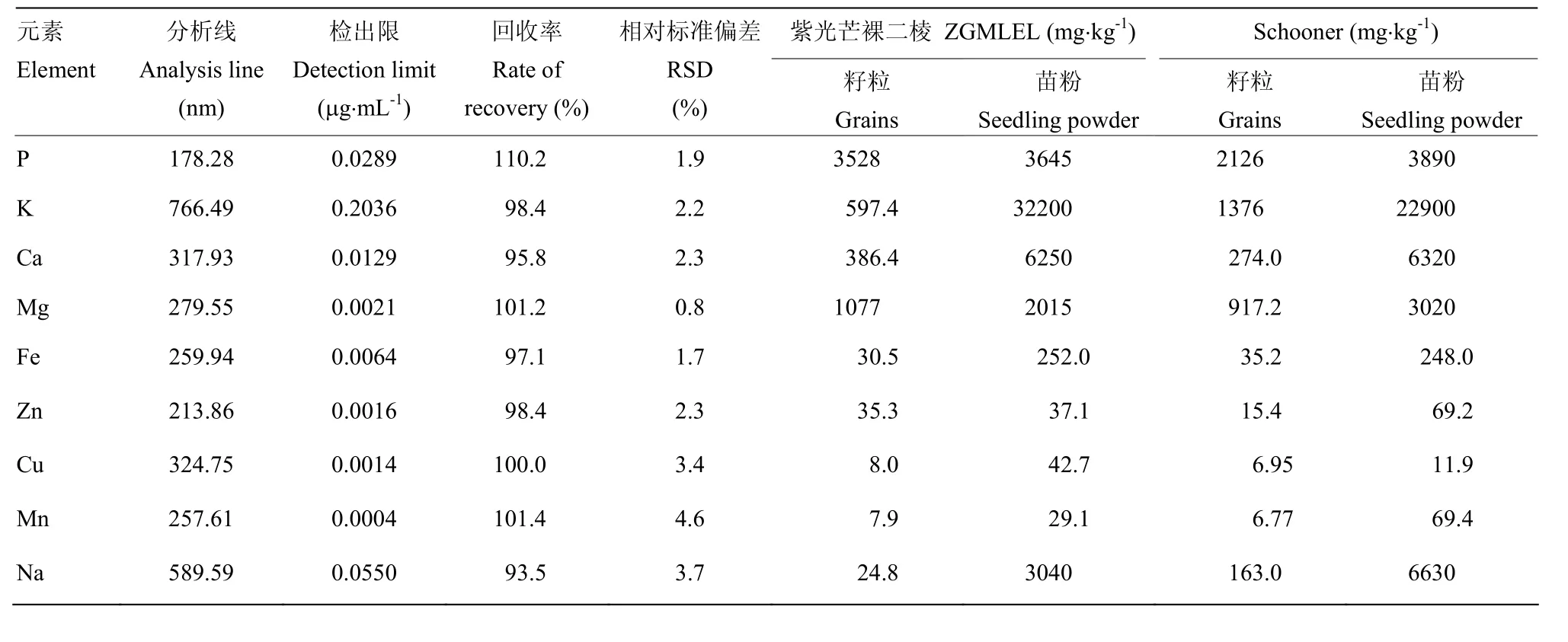
表1 检测方法的精密度、回收率试验和籽粒、苗粉的元素含量Table 1 Precision and recovery of the method as well as element content from barley grain and seedling powder
2 结果
2.1亲本及其RIL群体变种类型鉴定
以母本紫光芒裸二棱(Violiglabrum)与父本Schooner(Erectum)杂交培育的 194个 F8后代 RIL及其亲本,按穗芒颜色、护颖宽窄、侧小穗有无内外稃、芒性状、粒色、穗密度及籽粒稃状进行变种类型分类[4-5,10],划分为Nutans、Hypianthinum、Lanthonudum、Violiglabrum、Medicum、Nudum、Erectum、Violimedicum、 Neogenes、Glabrinudidubium、Jachanum、Luchuanicum 和Violierectum共13个变种类型(表2),因亲本及其RIL群体的棱型均为二棱、护颖宽窄均为窄护颖和芒长均为长芒,表2中作为分类性状的棱型、护颖宽窄和芒形状尚未列出。紫光芒裸二棱为疏穗、光芒、紫色裸大麦,Schooner为中穗、齿芒、黄色皮大麦。其中齿芒和光芒之比为156∶38,约为4∶1,而皮裸之比为143∶51,约为3∶1。表明大麦杂种后代的变种分类性状基因型间差异明显。
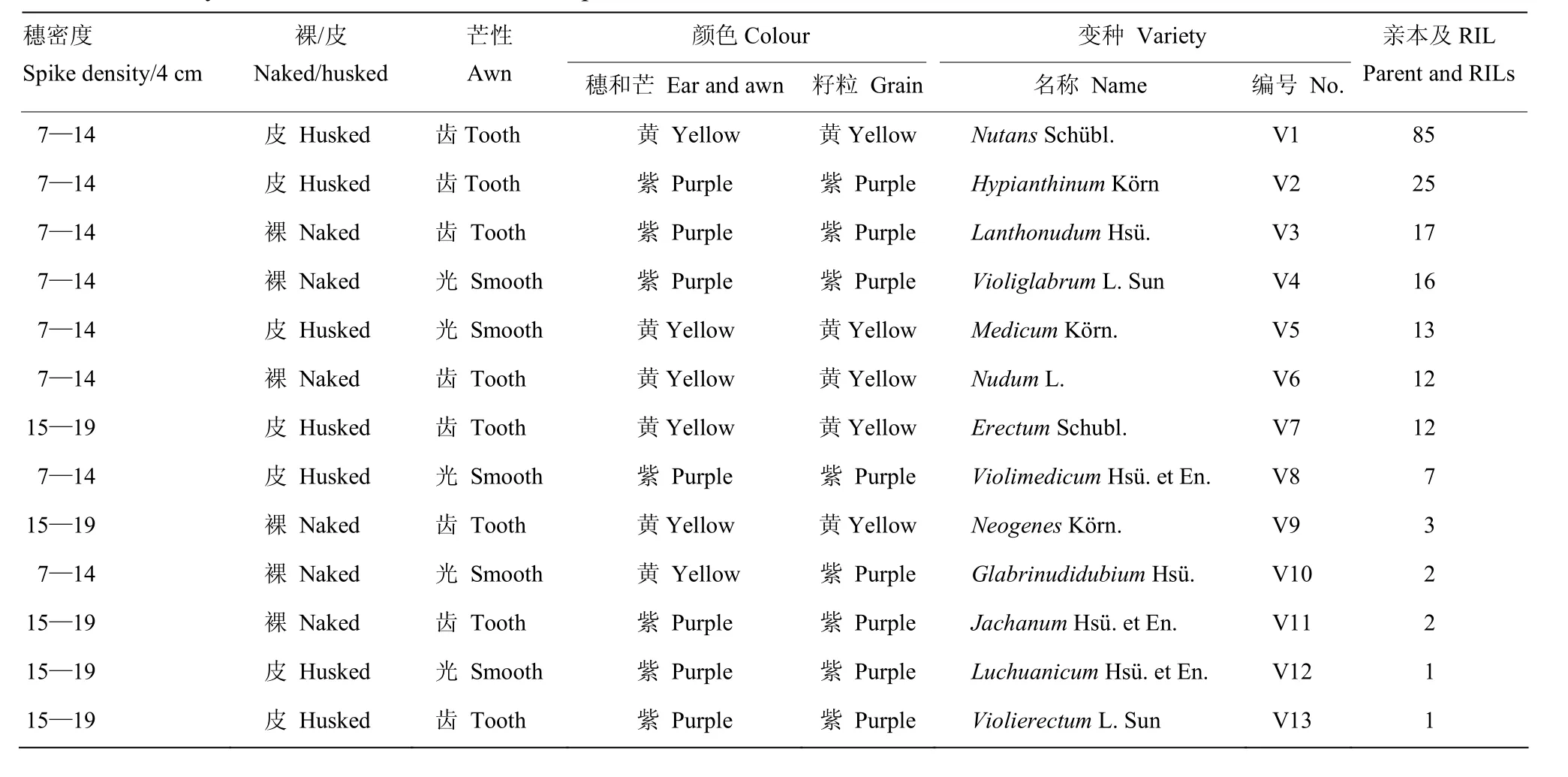
表2 亲本及其RIL的变种类型分类Table 2 Variety classification of RILs and their parents
2.2RIL群体变种类型间籽粒矿质元素差异
194个RIL及其亲本的13个变种类型籽粒9种矿质元素含量平均值及其标准差列于表3可知:亲本及其杂交后代群体籽粒矿质元素含量(mg·kg-1)依次为K(4 802)>P(2 593)>Mg(1 250)>Ca(568.3)>Na(190.5)>Fe(52.7)>Zn(39.5)>Mn(29.3)>Cu(14.1)。Violiglabrum与Erectum杂交后代13个变种类型间 9种矿质元素含量遗传变异差异较大为1.9—6.4倍,即籽粒P元素含量V8为V11的2.1倍,Zn元素V4为V11的2.2倍,Fe元素V9为V10 的2.9倍,Mn元素V8为V13的3.8倍,Mg元素V8为V11的1.9倍,Ca元素V5为V13的2.1倍,Cu元素V10为V13的6.4倍,Na元素V8为V13的1.9倍,K元素V5为V13的2.3倍。RIL的10个株系以上变种类型经差异显著性检验表明,P元素含量V5与V1、V3差异显著,V7与V2、V4差异显著。Zn元素V7与V1—V6共6个变种类型差异显著或极显著,V1与V3—V5共3个变种类型差异极显著,V2与 V4、V5差异极显著或显著。Mn元素V5与V1—V6共5个变种类型差异极显著或显著。Mg元素V7与V1—V6共6个变种类型差异显著或极显著。Ca元素V5与V1—V7共6个变种类型差异极显著或显著,V7与V2、V3差异显著。K元素V5与V7差异显著。
2.3RIL群体变种类型间苗粉矿质元素差异
194个RIL及其亲本的13个变种类型,苗粉中测定的9种矿质元素含量平均值及其标准差列于表4。亲本及其RIL群体的苗粉矿质元素含量(mg·kg-1)依次为K(31 860)>Ca(6 790)>Na(4 330)>P(4 191)>Mg(2 471)>Fe(241.3)>Zn(56.8)>Mn(41.3)>Cu(10.5)。Violiglabrum与Erectum杂交后代13个变种类型间,苗粉中9种矿质元素含量的遗传变异差异较大,为1.2—2.1倍,即苗粉P元素含量V12为V10的1.2倍,Zn元素V3为V8的1.3倍,Fe元素V9为V7的1.5倍,Mn元素V6为V7的1.2倍,Mg元素V6为V12的1.3倍,Ca元素V11为V12的1.4倍,Cu元素V4为V2的1.4倍,Na元素V13为V12的2.1倍,K元素V13为V2的1.3倍。RILs的10个株系以上变种类型经差异显著性检验表明,P元素含量V2与V4差异显著;Zn元素V4既与V1—V5共4个变种类型差异极显著,又与V7差异显著。Mn元素V7与V1—V6共6个变种类型差异极显著或显著,V1与V5和V6差异显著;Mg元素V1与V3、V5、V6共3个变种类型差异显著。Ca元素V7与V1、V2、V6差异显著,V2与V3差异显著。Cu元素V3与V1、V2差异显著。Na元素V4与V1—V6共5个变种类型差异极显著或显著;K元素V1与V7差异显著。
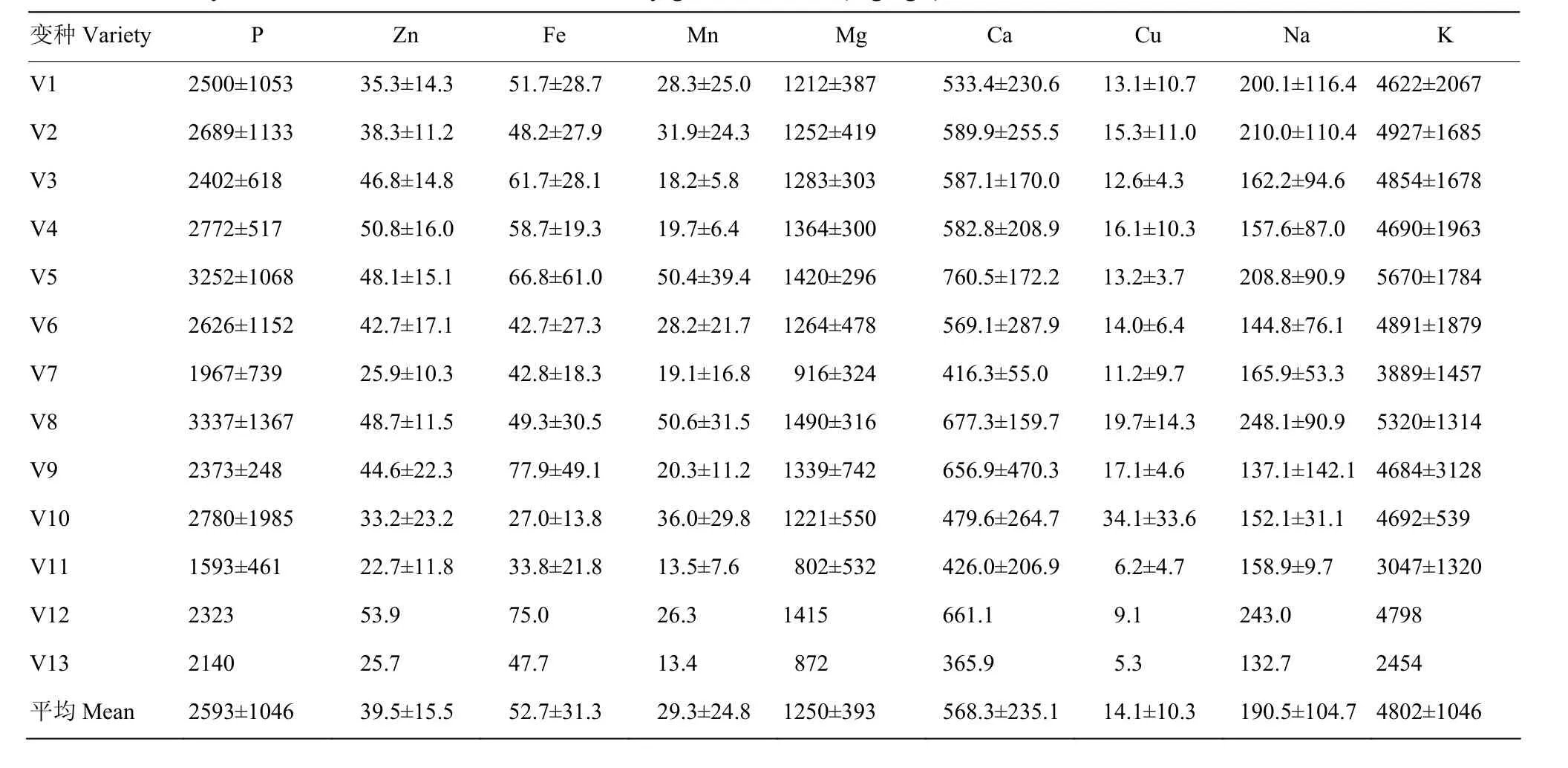
表3 大麦RIL群体籽粒元素含量变种类型间差异Table 3 Variety difference of element content in barley grains of RILs (mg·kg-1)
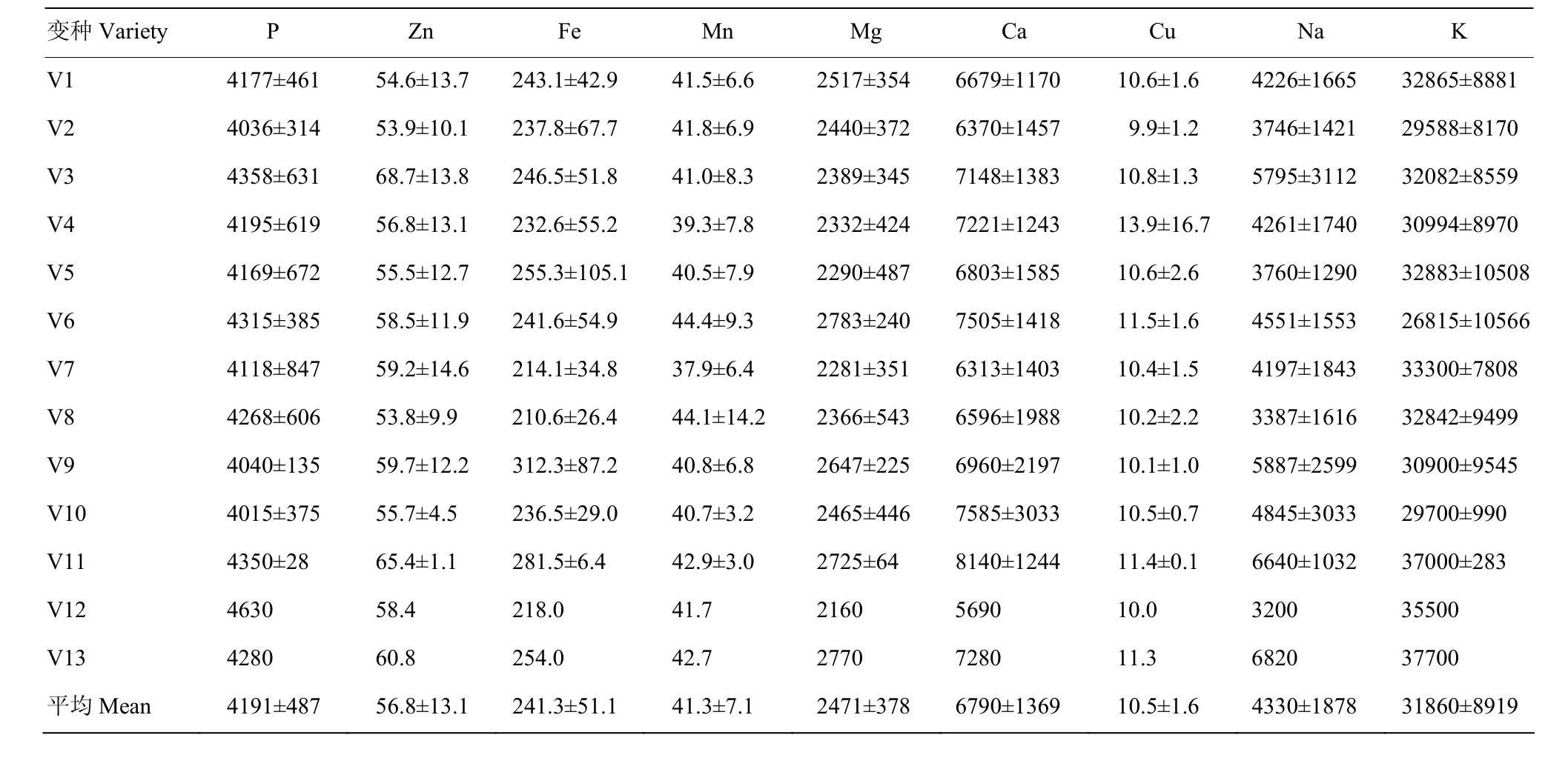
表4 大麦RIL群体苗粉元素含量变种类型间差异Table 4 Variety difference of element content in barley seedling powder of RILs (mg·kg-1)
2.4RIL变种类型间苗粉与籽粒矿质元素含量比
通过对194个RIL及其亲本13个变种类型的苗粉与籽粒9种矿质元素含量检测(表5)。Violiglabrum与Erectum杂交后代的13个变种类型之间,苗粉与籽粒矿质元素含量比值的遗传变异差异较大,其中,P、Zn、Fe、Mg、Ca、Na和K 7种矿质元素在苗粉与籽粒中的含量比值为1.08—51.39,尤其是Na(13.65—51.39)、Ca(8.61—19.90)和Fe(2.91—8.76)差异较大。
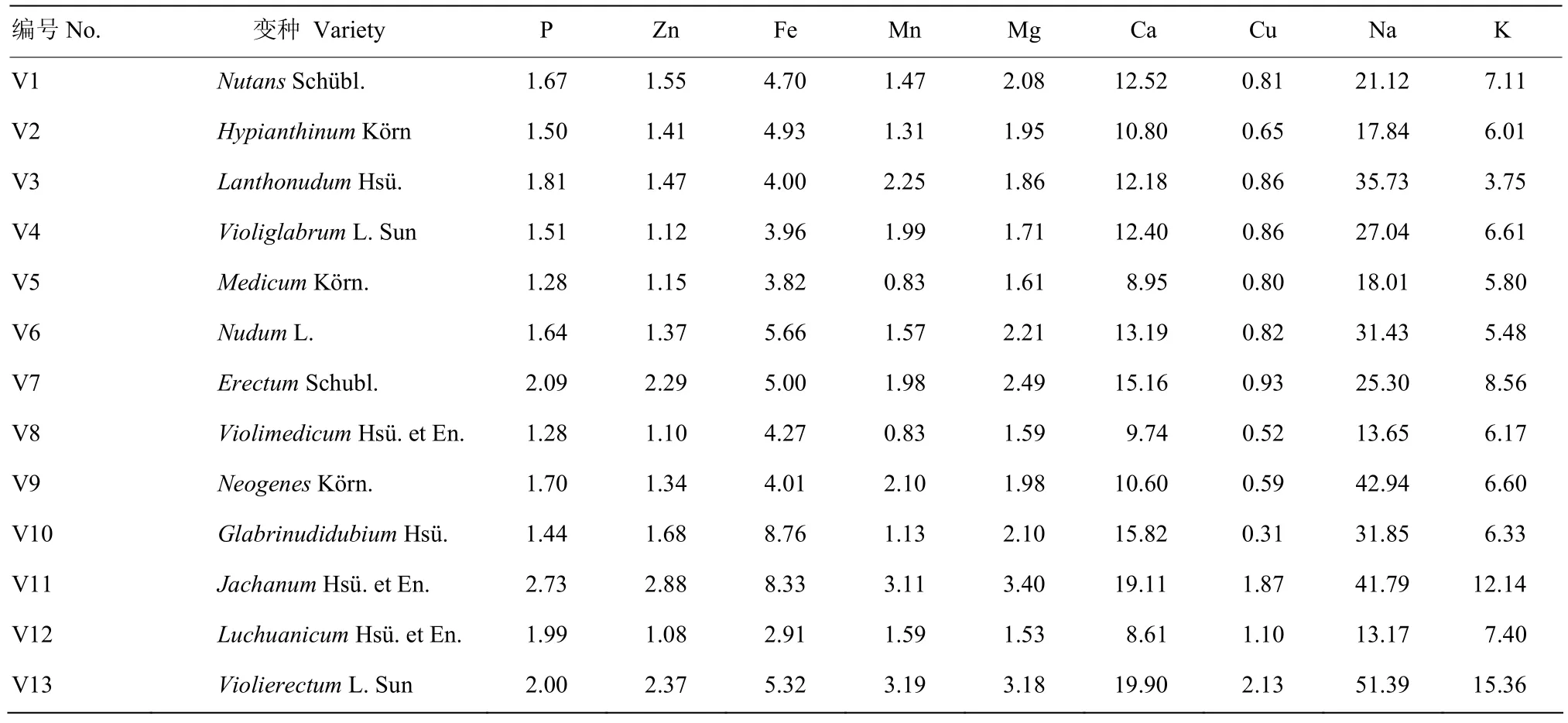
表5 大麦RIL群体苗粉/籽粒元素含量比值变种类型间差异Table 5 Ratio of element content for seedling powder vs grain among varieties from barley RILs
2.5RIL变种类型间籽粒及苗粉矿质元素含量聚类分析
通过对大麦RIL及其亲本13个变种类型籽粒和苗粉中9种矿质元素进行聚类分析(图1—图3)。194 个RIL株系及其亲本,13个变种类型间按D=6.0,基于籽粒9种矿质元素含量,可以划分为Ⅰ、Ⅱ、Ⅲ和Ⅳ共4个类群,群间差异明显而群内差异不大。Ⅰ由Schooner及其所属中穗型黄色皮大麦Erectum变种组成。Ⅱ由中穗型紫色裸大麦 Jachanum及其皮大麦Violierectum共2个变种组成。Ⅲ由疏穗型长光芒的黄色皮大麦Medicum和紫色皮大麦Violimedicum共2个变种组成。Ⅳ由长光芒紫粒大麦(Glabrinudidubium、Luchuanicum、Violiglabrum及紫光芒裸二棱)、长齿芒黄粒大麦(Nutans、Nudum、Neogenes)和长齿芒紫粒大麦(Hypianthinum、Lanthonudum、Jachanum、Violierectum)共10个变种组成。基于苗粉9种矿质元素含量,可以划分为Ⅰ、Ⅱ、Ⅲ和Ⅳ类群,群间差异明显而群内差异不大。Ⅰ由Schooner及其所属的中穗型黄色皮大麦(Erectum)、疏穗型黄色皮大麦(Nutans、Medicum)和光芒紫色皮大麦(Violimedicum、Luchuanicum)共5个变种类型组成。Ⅱ由疏穗型长齿芒紫色大麦(Hypianthinum、Lanthonudum)、疏穗型长光芒紫色裸大麦(Glabrinudidubium、Violiglabrum及紫光芒裸二棱)和中穗型黄色裸大麦(Neogenes)共5个变种组成。Ⅲ由疏穗型黄色裸大麦Nudum变种类型组成。Ⅳ由中穗型长齿芒紫色大麦(Jachanum和Violierectum)2个变种类型组成。基于籽粒与苗粉 9种矿质元素含量,可以划分为Ⅰ、Ⅱ、Ⅲ和Ⅳ共4个类群,群间差异明显而群内差异不大。Ⅰ由Schooner及其所属中穗型黄色皮大麦(Erectum)、长光芒紫色裸大麦(Glabrinudidubium、Violiglabrum及其紫光芒裸二棱)及疏穗型长齿芒裸大麦(Lanthonudum和Nudum)共 5个变种类型组成。Ⅱ由紫色皮大麦(Violimedicum、Luchuanicum和Hypianthinum)和黄色皮大麦(Medicum、Nutans)共5个变种类型组成。Ⅲ类由中穗型长齿芒紫色大麦(Jachanum 和Violierectum)共2个变种类型组成。Ⅳ由中穗型长齿芒黄色裸大麦(Neogenes)变种类型组成。这表明紫光芒裸二棱与澳大利亚优质啤酒大麦Schooner杂交后代,绝大多数变种类型与原始保守的云南地方品种紫光芒裸二棱的籽粒及苗粉矿质元素相近。
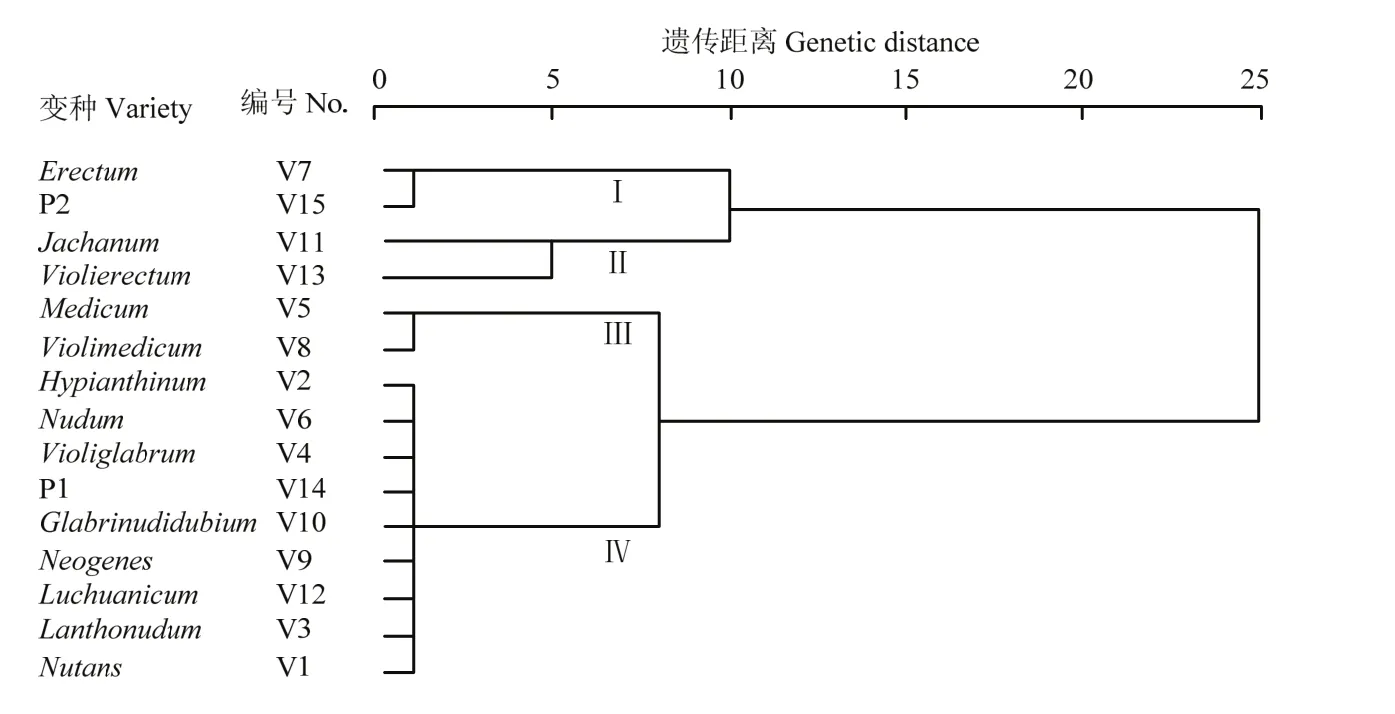
图1 亲本及RIL籽粒元素含量变种聚类Fig. 1 Variety cluster of element content in grain of RILs and their parents
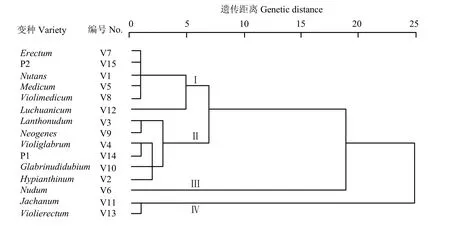
图2 亲本及RIL苗粉元素含量变种聚类Fig. 2 Variety cluster of element content in barley seedling powder of RILs and their parents
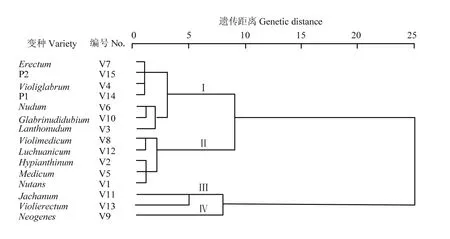
图3 亲本及RIL元素含量籽粒/苗粉变种聚类Fig. 3 Cluster of element content in grain/seedling powder of RILs and their parents
3 讨论
3.1中国原始的地方青稞品种
中国12 470份大麦种质26个表型性状的平均多样性指数,在28个省(市、区)之间的对比揭示云南是中国地方大麦种质最大的表型多样性中心[18]。云南栽培大麦含1个种2个亚种305个变种类型,其中新变种类型154个,二棱大麦和多棱大麦变种类型分别占变种类型总数的 42.3%和 57.7%,但西藏二棱大麦变种类型(43个)明显低于云南(129个),西藏多棱皮大麦变种类型(57个)明显低于云南(100个),云南高原是中国栽培大麦次生起源地和遗传多样性中心的一部分[10,19]。大麦黄色籽粒不含花色苷成分,紫、紫黑、黑、蓝和褐色籽粒分别含有 24、22、14、11 和2种成分,紫色籽粒以矢车菊素-3-O-葡萄糖苷、芍药素-3-O-葡萄糖苷和天竺葵素-3-O-葡萄苷为主,黑色籽粒以飞燕草素-3-O-葡萄糖苷为主,蓝色籽粒以飞燕草素-3-O-葡萄糖苷或矮牵牛素-3-O-葡萄糖苷为主[20]。本文以紫光芒裸二棱(Violiglabrum)与Schooner (Erectum)杂交培育的194个RIL及其亲本,划分为13个变种类型(其中紫色变种类型8个,黄色变种类型5个),分别占西藏和云南二棱大麦变种类型的30% 和 10.0%,揭示了紫光芒裸二棱是中国最原始的地方青稞品种及其独特类群形成的基因源,进一步揭示了云南高原地方大麦深色型籽粒比例高的成因[19]。
3.2大麦 RIL变种类型间籽粒及苗粉矿质元素含量的遗传变异
玉溪市蔬菜区种植的亲本及其 RIL,籽粒和苗粉中K、Ca、P、Mg和Na共5种矿质元素含量较高,分别为190.5—4 802 mg·kg-1和2 471—31 860 mg·kg-1。而Fe、Zn、Mn和Cu共5种矿质元素含量较低,分别为14.1—52.7 mg·kg-1和10.5—241.3 mg·kg-1;苗粉与籽粒中的含量比值依次为Na (22.7)>Ca(11.9)>K(6.6)>Fe(4.6)>Mg(2.0)>P(1.6)>Zn (1.4)=Mn(1.4)>Cu(0.7)。大麦苗粉和籽粒中的Ca含量分别是精米的68.3倍、5.7倍,K和Fe含量分别是精米的30.4倍和20.4倍[21]。另外,云功牌大麦苗粉的矿质元素含量(mg·kg-1)依次为K(51 600)>Ca(5 380)>P(3 440)>Mg(1 470)>Na(298)>Fe(206)>Mn(33.2)>Zn(29.9)>Cu(9.84)>Se(0.043)(未出版数据),K/Na比(173∶1)明显高于蔬菜区生产的大麦苗粉(7.4∶1),可见大麦苗粉的Ca、K和Fe含量极高,是其他作物和原粮食品难以媲美的。
Violiglabrum与Erectum杂交后代的13个变种之间,籽粒中9种矿质元素含量的遗传变异较大,为1.9 —6.4倍,尤其是P、Zn、Mn、Mg、Ca和K等元素含量差异显著或极显著;苗粉的矿质元素含量差异相对较小,为1.2—2.1倍,有8种含量变种类型间差异显著或极显著。苗粉与籽粒矿质元素含量的比值遗传变异较大,7种元素比值为1.1—51.4,尤其是Na(13.7 —51.4)、Ca (8.6—19.9)和Fe(2.9—8.8)。大麦富含半胱氨酸的金属硫蛋白(MT),在细胞内Zn和Cu动态平衡中发挥作用[13],在Mn动态平衡中,对细胞高尔基体上膜结合蛋白MTP转运具有重要作用[22],大麦遭受缺氧和低 pH/铝交叉胁迫时,保护和维持细胞内K+的动态平衡[23]。
2011—2012年云功牌大麦苗粉在中国 14个省(市)3 000人食用评价,据不完全统计,该中试产品具有改善睡眠、精力充沛、调节血糖、防治高血压、改善低血压、增强免疫力和肝功能、排毒去痘养颜、防便秘、改善肠胃功能、减轻痛风及高尿酸症、伤骨易恢复、改善心脏病、减轻神经性皮炎、抗癌、防衰老、减肥降血脂、减轻癫痫病、防痴呆、防脱发、消炎和抗溃疡等功效[1]。埃塞俄比亚是国际公认的人类和大麦起源中心及其抗癌等慢性病的协同进化基地[8]。350万年前古人类的饮食习惯从猿类以水果为食转变成以草为食[24]。目前,人类慢性病爆发与人类主食结构变化有关,推测从古人到现代人主食结构大体分为水果蔬菜型、肉质嫩草型、五谷杂粮型、精米精面型和精米精面草粉型。精米精面型是目前人类慢性病爆发的重要诱因之一,精米精面草粉型是人类健康主食的重要组成部分。大麦苗粉及其功能食品(如大麦苗粉米线、大麦苗粉饵丝和大麦苗粉面条等)是以精米为主食的亚洲人群解决Ca和Fe缺乏最经济有效的途径。因此,培育高功能成分的大麦苗粉新品种,构建亚洲现代人群的精米(或精面)+大麦苗粉的主食结构是解决人类慢性病最经济有效的途径。
3.3大麦 RIL变种类型苗粉与籽粒矿质元素含量聚类
中国栽培大麦变种以疏穗、窄护颖、长齿芒、穗粒深色型为主,高原紫外线强、干旱和冷凉等逆境下易驯化成深色型种质,二棱大麦Nutans和Erectum分布较广[4,16]。194个RIL株系及其亲本无论是基于籽粒和苗粉的9种矿质元素含量,还是苗粉/籽粒含量比值的聚类分析,13个变种类型间按D=6.0,均划分为Ⅰ、Ⅱ、Ⅲ和Ⅳ共4个类群。其中,中穗型Jachanum与Violierectum 共同组成一类, 光芒型 Medicum 与Violimedicum均归属同类,紫青稞型 Lanthonudum、Violiglabrum、Glabrinudidubium和Violiglabrum均归属同类群。紫光芒 Violimedicum、Violiglabrum和Luchuanicum共3个变种类型是云南特有的新变种[5]。籽粒黑色基因位于第 1染色体上,与单核苷酸基因CAPS027-CAPS029标记连锁[25]。大麦有11个位点影响籽粒原花青素,花青素ant17在色素组织表达且在2HL臂上与抗穗发芽的黄烷酮3-羟化酶共分离[26]。青藏高原及其周边地区是栽培大麦驯化中心之一,抗旱及耐冷等优异种质极为丰富[10]。西藏野生大麦是中国地方大麦的祖先[11]。二棱大麦变种类型云南 (129个)为西藏的3倍,而多棱皮大麦变种类型云南(100个)为西藏的 1.8倍[12,19]。表明大麦籽粒及苗粉中的 P、Zn、Mn、Mg、Ca和K元素可能在大麦变种类型演化中起重要作用。
4 结论
以Violiglabrum与Erectum变种杂交后代RIL及其亲本划分为13个变种类型,约占云南二棱大麦变种类型 10.0%,即用原始独特的地方青稞品种紫光芒裸二棱作亲本杂交后代遗传变异大。籽粒的P、Zn、Mn、Mg、Ca和K元素含量变种类型间差异显著。苗粉的8种矿质元素含量变种类型间差异显著。苗粉与籽粒中的矿质元素含量比值遗传变异较大。13个变种类型划分为4个类群。其中,中穗型Jachanum与Violierectum同类,光芒型Medicum与Violimedicum同类,紫青稞型Lanthonudum、Violiglabrum、Glabrinudidubium和Violiglabrum同类。籽粒的矿质元素含量变种类型间差异显著而苗粉中含量差异较小,类群间差异明显而群内差异不显著。矿质元素在大麦变种类型驯化中起重要作用。
References
[1] 曾亚文, 普晓英, 张京, 郭刚刚, 杜娟, 杨涛, 杨树明, 杨加珍. 中
国西南大麦产业发展综合研究利用. 中国农业科技导报, 2013,15(3): 48-56. ZENG Y W, PU X Y, ZHANG J, GUO G G, DU J, YANG T, YANG S M, YANG J Z. Synthetic research and utilization on industrial development of barley in Southwestern China. Journal of Agricultural Science and Technology, 2013, 15(3): 48-56. (in Chinese)
[2] ATKKINSON F, FOSTER-POWELL K, BRAND-MILLER J C. International table of glycemic index and glycemic load values 2008. Diabetes Care, 2008, 31(12): 2281-2283.
[3] SOONG Y Y, TAN S P, LEONG L P, HENRY J K. Total antioxidant capacity and starch digestibility of muffins baked with rice, wheat, oat,corn and barley flour. Food Chemistry, 2014: 462-469.
[4] 徐廷文. 中国栽培大麦的分类和变种鉴定. 中国农业科学, 1982,15(6): 36-47. XU T W. Classification and identification of varieties of Chinese cultivated barley. Scientia Agricultura Sinica, 1982, 15(6): 36-47. (in Chinese)
[5] 王建林, 栾运芳, 大次卓嘎, 胡单. 西藏栽培大麦变种组成和分布规律研究. 中国农业科学, 2006, 39(11): 2163-2169. WANG J L, LUAN Y F, DACIZHUOGA, HU D. Distribution and composition study of Tibetan cultivated barley varieties. Scientia Agricultura Sinica, 2006, 39(11): 2163-2169. (in Chinese)
[6] POETS A M, FANG Z, CLEGG M T, MORRELL P L. Barley landraces are characterized by geographically heterogeneous genomic origins. Genome Biology, 2015, 6: 173.
[7] ZENG X, LONG H, WANG Z, ZHAO S, TANG Y, HUANG Z,WANG Y, XU Q, MAO L, DENG G, YAO X, LI X, BAI L, YUAN H,PAN Z, LIU R, CHEN X,WANGMU Q, CHEN M, YU L, LIANG J,DUNZHU D, ZHENG Y, YU S, LUOBU Z, GUANG X, LI J, DENG C, HU W, CHEN C, TABA X, GAO L, LV X, ABU Y B, FANG X,NEVO E, YU M, WANG J, TASHI N. The draft genome of Tibetan hulless barley reveals adaptive patterns to the high stressful Tibetan Plateau. Proceedings of the National Academy of the Sciences of the United States of America, 2015, 27, 112(4): 1095-1100.
[8] ZENG Y W, DU J, PU X Y, YANG J Z, YANG T, YANG S M, YANG X M. Coevolution between human's anticancer activities and functional foods from crop origin center in the world. Asian Pacific Journal of Cancer Prevention, 2015, 16(6): 2119-2128.
[9] LORDKIPANIDZE D, PONCE DE LEÓN M S, MARGVELASHVILI A,RAK Y, RIGHTMIRE G P, VEKUA A, ZOLLIKOFER C P. A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 2013, 342: 326-331.
[10] DAI F, NEVO E, WU D, COMADRAN J, ZHOU M, QIU L, CHEN Z, BEILES A, CHEN G, ZHANG G. Tibet is one of the centers of domestication of cultivated barley. Proceedings of the National Academy of the Sciences of the United States of America, 2012,109(42): 16969-16973.
[11] REN X, NEVO E, SUN D, SUN G. Tibet as a potential domestication center of cultivated barley of China. PLoS ONE, 2013, 8(5): e62700.
[12] DAI F, CHEN Z H, WANG X L, LI Z F, JIN G L, WU D Z, CAI S G,WANG N, WU F B, NEVOD E, ZHANG G P. Transcriptome profiling reveals mosaic genomic origins of modern cultivated barley. Proceedings of the National Academy of the Sciences of the United States of America, 2014, 111(37): 13403-13408.
[13] HEGELUND J N, SCHILLER M, KICHEY T, HANSEN T H,PEDAS P, HUSTED S, SCHJOERRING J K. Barley metallothioneins:MT3 and MT4 are localized in the grain aleurone layer and show differential Zinc binding. Plant Physiology, 2012, 59(3): 1125-1137.
[14] RABOY V, CICHY K, PETERSON K, REICHMAN S, SOMPONG U, SRINIVES P, SANEOKA H. Barley (Hordeum vulgare L.) low phytic acid 1-1: An endosperm-specific, filial determinant of seed total phosphorus. Journal of Heredity, 2014, 105(5): 656-665.
[15] 辛培尧, 普晓英, 杜娟, 杨涛, 曾亚文. 大麦籽粒和苗粉蛋白质含量的检测. 麦类作物学报, 2016, 31(1): 58-61. XIN P Y, PU X Y, DU J, YANG T, ZENG Y W. Protein content determination of barley grain and seedling powder. Journal of Triticeae Crops, 2016, 31(1): 58-61. (in Chinese)
[16] Zeng Y W,Yang J Z, Du J,Pu X Y, Yang X M,Yang S M, Yang T. Strategies of functional foods promote sleep in human being. Current Signal Transduction Therapy, 2014, 9(3): 148-155.
[17] 孙立军. 中国栽培大麦变种及其分布特点. 中国农业科学, 1988,21(2): 25-31. SUN L J. Varieties of cultivated barley and their distribution in China. Scientia Agricultura Sinica, 1988, 21(2): 25-31. (in Chinese)
[18] 张京, 曹永生. 我国大麦基因库的群体结构和表型多样性研究. 中国农业科学, 1999, 32(4): 20-26. ZHANG J, CAO Y S. Population structure and phenotypic diversity of barley in China’s gene bank. Scientia Agricultura Sinica, 1999,32(4): 20-26. (in Chinese)
[19] 曾亚文, 王建军. 云南省栽培大麦的分类研究. 作物品种资源,1998(2): 7-9. ZENG Y W, WANG J J. Classification of varieties of Yunnan cultivated barley. Crop Genetic Resources, 1998(2): 7-9. (in Chinese)
[20] 谢文英, 陈升位, 王楠, 王家曦, 杨映宝, 王乐. 25个地方大麦籽粒黄酮类化合物及其含量的差异分析. 云南农业大学学报, 2015, 30(5): 657-664. XIE W Y, CHEN S W, WANG N, WANG J X, YANG Y B, WANG L. The difference analysis of the grain flavonoiad components and their contents of 25 local barley varieties. Journal of Yunnan Agricultural University, 2015, 30(5): 657-664. (in Chinese)
[21] 曾亚文, 汪禄祥, 杜娟, 杨树明, 王雨辰, 黎其万, 孙正海, 普晓英, 杜威. ICP-AES法检测云南稻精米和糙米与土壤矿质元素间的关联性. 光谱学与光谱分析, 2009, 29(5): 1413-1417. ZENG Y W, WANG L X, DU J, YANG S M, WANG Y C, LI Q W,SUN Z H, PU X Y, DU W. Correlation of mineral elements between milled and brown rice and soils in Yunnan studied by ICP-AES. Spectroscopy and Spectral Analysis, 2009, 29(5): 1413-1417. (in Chinese)
[22] PEDAS P, SCHILLER STOKHOLM M, HEGELUND J N,LADEGÅRD A H, SCHJOERRING J K, HUSTED S. Golgi localized barley MTP8 proteins facilitate Mn transport. PLoS ONE, 2014, 9(12):e113759.
[23] MA Y, ZHU M, SHABALA L, ZHOU M, SHABALA S. Conditioning of roots with hypoxia increases aluminum and acid stress tolerance by mitigating activation of K+efflux channels by ROS in barley: Insights into cross-tolerance mechanisms. Plant & Cell Physiology, 2016,57(1): 160-173.
[24] WYNN J G, SPONHEIMER M, KIMBEL W H, ALEMSEGED Z,REED K, BEDASO Z K, WILSON J N. Diet of Australopithecus afarensis from the Pliocene Hadar Formation, Ethiopia. Proceedings of the National Academy of the Sciences of the United States of America, 2013, 110(26): 10495-10501.
[25] BUNGARTZ A, KLAUS M, MATHEW B, LÉON J, NAZ A A. Development of new SNP derived cleaved amplified polymorphic sequence marker set and its successful utilization in the genetic analysis of seed color variation in barley. Genomics, 2016, 107(2/3):100-107.
[26] HIMI E, TAKETA S. Barley Ant17, encoding flavanone 3-hydroxylase (F3H), is a promising target locus for attaining anthocyanin /proanthocyanidin-free plants without pleiotropic reduction of grain dormancy. Genome, 2015, 58(1): 43-53.
(责任编辑 李莉)
Difference of Elements in Different Types of Seedling Powder and Its Grains of Barley Recombinant Inbred Lines
ZENG Ya-wen1, WANG Lu-xiang2, YANG Xiao-meng1, YANG Jia-zhen1, DU Juan1, PU Xiao-ying1,DU Li-juan2, YANG Shu-ming1, XIAO Ya3, YANG Tao1
(1Biotechnology and Genetic Resources Institute, Yunnan Academy of Agricultural Sciences/Agricultural Biotechnology Key Laboratory of Yunnan Province, Kunming 650205;2Institute of Quality Standards and Testing Technology, Yunnan Academy of Agricultural Sciences, Kunming 650223;3Shigatse City Agricultural Research Institute of Tibet Autonomous Region, Shigatse 857000, Tibet )
【Objective】Variety type and element content as well as genetic variation based on seedling powder and grain for barley recombinant inbred lines (RILs) provide not only the basic information of barley grain and seedling powder for developing functional food that is better for human health, but also the evidences about genetic variation of cultivated barley. 【Method】Varieties classification of 194 RILs from a cross Ziguangmangluoerleng (ZGMLEL) × Schooner and their parents can be made according to the types of ear and awn color, glume size, palea and lemma of lateral spikelets, awn shape, grain color, spike density,naked or husked seven traits. The contents of 9 mineral elements (P, Zn, Fe, Mn, Mg, Ca, Cu, Na and K) in seedling powder and grains of 194 barley recombinant inbred lines (RILs) and their parents were determined by using ICP-AES. 【Result】The 194 RILs based on hybridization breeding between Violiglabrum and Erectum as well as their parents were divided into 13 variety types,including Nutans, Hypianthinum, Lanthonudum, Violiglabrum, Medicum, Nudum, Erectum, Violimedicum, Neogenes, Glabrinudidubium,Jachanum, Luchuanicum, and Violierectum. It is obvious in genotype difference of variety classification traits. There are significant differences of minerial element contents among variety types in grains (1.9 to 6.4 times) and lower difference among variety types in barley seedling powder (1.2 to 2.1 times). Differences of the highest times for variety types in grain element contents is in order of Cu (Glabrinudidubium/Violierectum=6.4)>Mn(Violimedicum/Violierectum=3.8)>Fe (Neogenes/Glabrinudidubium=2.9)>K(Medicum/Violierectum=2.3)>Zn(Violiglabrum/Jachanum=2.2)>P(Violimedicum/Jachanum=2.1)=Ca (Medicum/Violierectum=2.1)>Mg (Violimedicum/Jachanum=1.9)=Na (Violimedicum/Violierectum=1.9); Differences of the highest times for variety types in seedling element contents is in order of Na (Violierectum/Luchuanicum=2.1)>Fe (Neogenes/Erectum=1.5)>Ca (Jachanum/ Luchuanicum=1.4) = Cu (Violiglabrum/Hypianthinum=1.4)>Zn (Lanthonudum/Violimedicum=1.3) = Mg (Nudum/Luchuanicum=1.3)= K (Violierectum/Hypianthinum=1.3)>P (Luchuanicum/Glabrinudidubium=1.2) =Mn (Nudum/Erectum = 1.2), however, the difference of 6 element contents (P, Zn, Mn, Mg, Ca and K) among varieties in grains and seedling powder is significant. The ratio of 7 element contents of barley seedling powder versus grains is 1.1-51.4 times. Differences of the highest times for variety types in element contents for seedling vs grains ratio is in order of Na (Violierectum 51.4)>Ca (Violierectum 19.9)>K (Violierectum 15.4)>Fe (Glabrinudidubium 8.8)>Mg (Jachanum 3.4)>Mn (Violierectum 3.2)>Zn (Jachanum 2.9)>P (Jachanum 2.7)>Cu (Violierectum 2.1). There are 194 RILs lines and their parents can be divided into 4 classes based on 9 mineral element contents for barley grains or seedling powder or seedling powder / grains, and the difference between groups is significant but the difference within a group is insignificant. The most variant type of their progenies are similar in mineral element contents in grains or seedling powder of Ziguangmangluoerleng with the original Yunnan landraces.【Conclusion】There is a significant difference between groups but lower difference among variety types in minerial element contents in grain and seedling powder. There is a significant difference in mineral element contents in grain but lower difference in seedling powder. These evidences support that mineral elements play an important role in the domestication of cultivated barley variety type.
barley; ICP-AES; mineral element; grains; seedling powder
2016-02-29;接受日期:2016-04-25
国家自然科学基金(31260326)、国家现代农业产业技术体系建设专项(CARS-05)
联系方式:曾亚文,Tel:0871-65894145;Fax:0871-65894145;E-mail:zengyw1967@126.com
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
环境卫生工程(2021年4期)2021-10-13
意林·作文素材(2021年15期)2021-10-08
疯狂英语·新读写(2021年2期)2021-02-25
学生天地(2019年33期)2019-08-25
中成药(2018年11期)2018-11-24
现代园艺(2017年21期)2018-01-03
小学生导刊(高年级)(2016年4期)2016-04-08
华人时刊(2016年19期)2016-04-05
植物营养与肥料学报(2012年1期)2012-10-26
