
水稻DNA多态标记的开发与验证
2016-10-31 09:20邱振楠徐乾坤王小琦马伯军
浙江农业学报 2016年3期
邱振楠,徐乾坤,王小琦,赵 娟,赫 磊,张 森,马伯军,钱 前,朱 丽,*
(1.浙江师范大学 化学与生命科学学院,浙江 金华 321004;2.中国水稻研究所 水稻生物学国家重点实验室,浙江 杭州310006)
水稻DNA多态标记的开发与验证
邱振楠1,2,徐乾坤1,王小琦2,赵娟2,赫磊2,张森2,马伯军1,钱前2,朱丽2,*
(1.浙江师范大学 化学与生命科学学院,浙江 金华 321004;2.中国水稻研究所 水稻生物学国家重点实验室,浙江 杭州310006)
利用203对分子标记对3个粳稻品种(日本晴、中花11、武运粳7号)和4个籼稻品种(9311、明恢63、台中1号及南京6号)进行多态性分析,筛选到92对在籼粳间存在差异的标记、34对粳稻品种间存在差异的标记及80对在籼稻品种间存在差异的标记。对部分在多个品种中存在多态性的标记进行测序,结果表明:大部分微卫星标记在不同品种间产物的大小差异是由重复序列重复次数的多少引起,而插入/缺失标记在品种间的序列差异则各有不同,如部分非重复序列的一次或多次重复、长片段的插入或缺失等。这些多态性好、扩增稳定的分子标记的开发和多态性验证为分子标记在种质鉴定、进化分析、遗传多样性检测、分子标记辅助选育等方面的广泛使用奠定了基础。
水稻;分子标记;多态性;品种鉴定
自20世纪70年代DNA水平的多态性鉴定技术问世以来,越来越多的分子标记被不断开发和利用[1-6]。水稻全基因组序列的公布及重要品种重测序计划的陆续启动和完成,更是极大的丰富了人们对水稻序列多态性的认识和了解,拓展了其应用领域及空间[7-11]。高密度的分子标记不仅是基因定位和克隆的基础,而且还可以在种质鉴定、进化分析、遗传多样性检测、分子标记辅助选育等方面发挥作用[12-16],已有的研究表明,6对简单序列长度多态性(simple sequence length polymorphism,SSLP)引物即可区分71个不同的品种[17]。李小湘等[18]用24对简单重复序列标记(simple sequence repeats,SSR)对136份湖南地方稻进行遗传差异分析,结果表明,SSR标记遗传相似性系数可以部分代表表型性状遗传距离,但仅使用24对SSR标记进行差异性分析,标记多态性不足,对遗传差异的分析还不够全面。
到目前为止,从DNA序列层面上已经获得了数量庞大的多态标记,但这些标记在不同水稻品种间的多态性验证并不多见,高通量测序虽然可以快速、精确地分析品种间的多态性,但对大量品种进行高通量测序,在成本方面显然不具操作性。用PCR的方法筛选更多稳定可靠的分子标记用于品种鉴定、亲缘关系分析及不同群体的基因定位和克隆则更加简便易行[19-21]。本研究利用203对分子标记,其中184对为新开发的分子标记,对3个粳稻品种及4个籼稻品种进行多态性分析,并对部分在多个品种中存在多态性的标记进行测序分析,筛选到一批多态性好、扩增稳定的分子标记,这些标记的开发和多态性验证不仅可为分子标记辅助育种提供更多有效的标记资源,还可广泛应用于种质鉴定、进化分析、遗传多样性检测等领域。
1 材料与方法
1.1材料
选用的研究材料分别为水稻品种日本晴(NIP,粳稻),中花11(ZH11,粳稻),武运粳7号(WY7,粳稻),9311(籼稻),明恢63(MH63籼稻),台中1号(TN1,籼稻),南京6号(NJ6,籼稻),均为本实验室保存,全部材料的种植地均为浙江富阳。其中NIP和9311为已测序品种,用于验证多态性的正确性。ZH11,WY7,MH63,TN1及NJ6均为生产中常用的水稻品种。203对分子标记中184对为新开发的标记,19对为已有的RM标记[22],所有的标记及对应的测序引物合成均由英潍捷基(上海)贸易有限公司完成。
1.2方法
1.2.1水稻基因组DNA提取
按照卢扬江等[23]报道的方法提取水稻基因组DNA。
1.2.2引物设计及PCR检测
根据NCBI(http://www.ncbi.nlm.nih.gov/)及Gramene (http://www.gramene.org/genome browser/index.html) 上的水稻全基因组信息,Blast分析日本晴和9311间序列的差别,并近似认为这些差别在待测的7个品种间也存在,利用这些差异借助生物软件Primer Premier 5.0和DNAMAN设计引物,在7个品种间筛选有多态性的标记,分析不同品种间多态性。15 μL PCR反应体系:20 mmol·L-1Tris-HCl,10 mmol·L-1(NH4)2SO4,10 mmol·L-1KCl,2 mmol·L-1MgCl2,1% Triton X-100,pH 8.8,10.0 nkat Taq酶,0.17 mmol·L-1dNTPs,0.33 μmol·L-1引物,100 ng模板DNA。在Applied Biosystems 9700 PCR仪上进行扩增,反应条件为94℃ 4 min;94℃ 30s,退火(退火温度随引物各异)30 s,72℃ 1 min,35个循环;72℃ 10 min。扩增产物在4%琼脂糖凝胶中电泳分离,溴化乙锭染色,紫外灯下观察。
1.2.3多态性序列分析
将筛选获得的在多个品种间均有多态的标记,重新设计测序引物,PCR扩增对应多态品种,产物回收后由上海桑尼生物科技有限公司完成测序。用DNASTAR中的SeqMan软件分析序列多态性。多态标记的设计基于NCBI中提供的籼稻和粳稻测序数据比对完成。在水稻数据库Gramene中对标记的旁邻序列进行Blast分析(http://blast.gramene.org/Multi/blastview)。
2 结果与分析
利用203对SSR标记、插入/缺失标记(insertion/deletion,InDels)及序列标签位点(sequence-tagged site,STS)标记在7个品种间进行多态性筛选,其中184对为新开发标记,19对为RM引物。所有标记中有9对在7个品种中均无多态。在全部可正常扩增而且在不同材料中有多态的194对标记中,只在籼粳间存在差异的标记有92对(图1,表1为部分结果),粳稻品种间存在差异的标记有34对(图2,表2为部分结果),籼稻品种间存在差异的标记有80对(图3,表3为部分结果)。
部分标记在粳稻间存在差异,可区分不同品种的粳稻,如标记ZB1-1,ZB2-26,ZB2-11,ZB3-13等在武运粳7号的带型与日本晴和中花11不同;ZB2-15,ZB7-1,ZB7-2和KS9-24等可特异地区分中花11,其带型与日本晴和武运粳7号不同;ZB10-2,ZB10-6,ZB11-13和ZB11-15等可特异地区分日本晴,其带型与中花11和武运粳7号不同(图2,表2为部分结果)。
部分标记在籼稻间存在差异,可区分不同品种的籼稻品种,如标记ZB2-15,ZB4-5,ZB4-6,ZB6-3,KS7-25等可特异区分9311与其他3个籼稻品种;ZJ8-7,ZB2-26,ZB2-11,ZB3-2,ZB8-4等可特异区分明恢63;ZB1-11,ZB1-7,ZB2-10,ZB9-4,ZB7-3,ZB7-4等可特异区分台中1号;ZB2-25,ZB8-11,ZB9-8,ZJ8-3和ZB12-4等可特异区分南京6号;此外,还有一些标记可区分两种籼稻品种,如ZB1-2在明恢63、南京6号中的带型与在9311和台中1号的带型不同;ZB2-24在9311和南京6号中的带型与在明恢63和台中1号的带型不同(图3,表3为部分结果)。
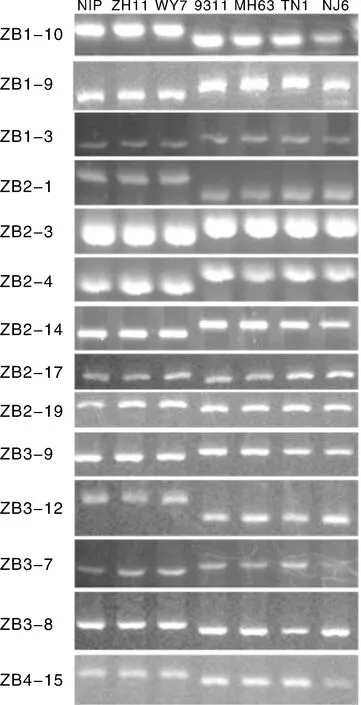
图1 部分只在籼粳稻间存在差异的标记Fig.1 Several markers showed polymorphism among indica and japonica rice
为了进一步明确标记在不同品种中序列的差异,对部分标记进行测序验证,结果显示,大部分微卫星标记在不同品种间产物的大小差异由重复序列重复次数的多少引起,而插入/缺失标记品种间序列差异则各有不同,如部分非重复序列的一次或多次重复、长片段的插入或缺失等(图4)。对203对标记多态位点的旁邻序列进行分析,结果表明,91.7%的标记多态性发生在基因的非编码区或尚未被预测功能的区段内,其中发生在尚未被预测功能的区段的有52.9%,基因内含子、5’UTR、ATG前面的外显子及TAG(TAA,TGA)后面的外显子的有38.9%,只有8.3%的多态位点发生在基因编码区(表4)。
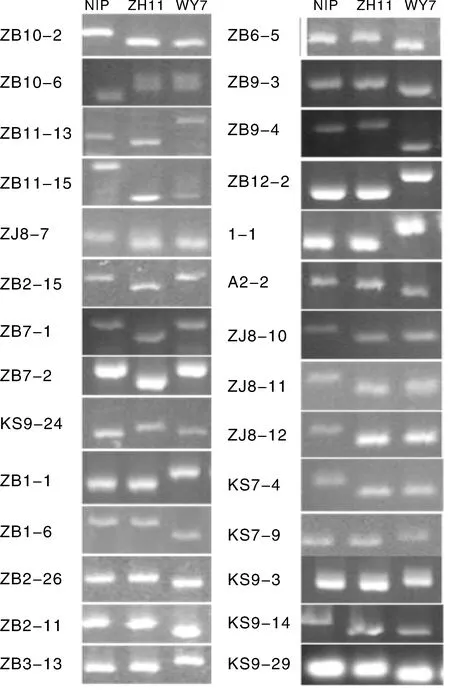
图2 部分粳稻间多态标记的PCR检测结果Fig.2 PCR results of several markers polymorphism among japonica rice
表1部分在籼粳间存在差异的标记序列
Table 1Sequences of several markers polymorphism among indica and japonica rice
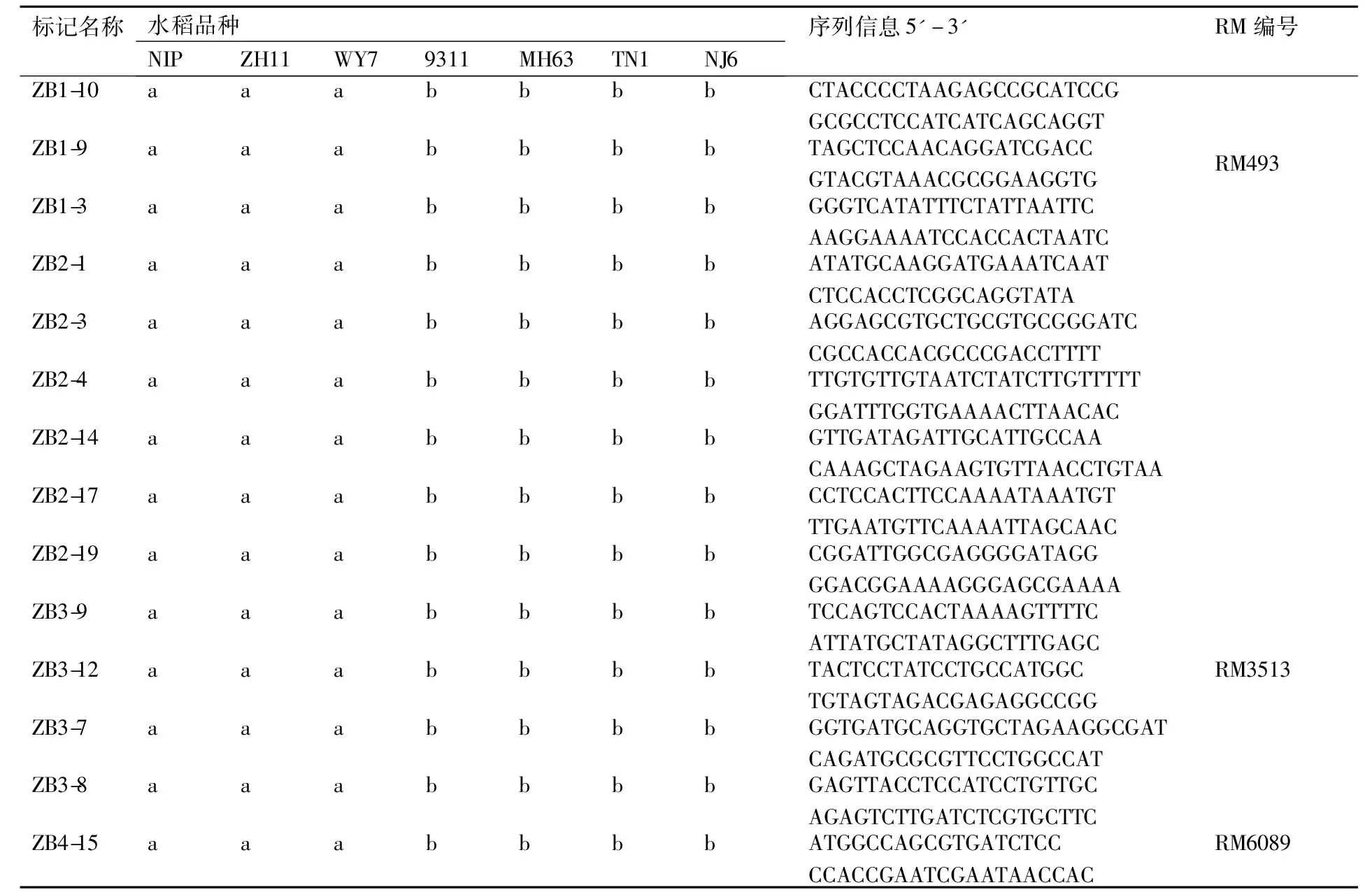
标记名称水稻品种NIPZH11WY79311MH63TN1NJ6序列信息5'-3'RM编号ZB1-10aaabbbbCTACCCCTAAGAGCCGCATCCGGCGCCTCCATCATCAGCAGGTZB1-9aaabbbbTAGCTCCAACAGGATCGACCGTACGTAAACGCGGAAGGTGRM493ZB1-3aaabbbbGGGTCATATTTCTATTAATTCAAGGAAAATCCACCACTAATCZB2-1aaabbbbATATGCAAGGATGAAATCAATCTCCACCTCGGCAGGTATAZB2-3aaabbbbAGGAGCGTGCTGCGTGCGG-GATCCGCCACCACGCCCGACCTTTTZB2-4aaabbbbTTGTGTTGTAATCTATCTT-GTTTTTGGATTTGGTGAAAACTTAACACZB2-14aaabbbbGTTGATAGATTGCATTGCCAACAAAGCTAGAAGTGTTAACCT-GTAAZB2-17aaabbbbCCTCCACTTCCAAAATAAATGTTTGAATGTTCAAAATTAGCAACZB2-19aaabbbbCGGATTGGCGAGGGGATAGGGGACGGAAAAGGGAGCGAAAAZB3-9aaabbbbTCCAGTCCACTAAAAGTTTTCATTATGCTATAGGCTTTGAGCZB3-12aaabbbbTACTCCTATCCTGCCATGGCTGTAGTAGACGAGAGGCCGGRM3513ZB3-7aaabbbbGGTGATGCAGGTGCTAGAAG-GCGATCAGATGCGCGTTCCTGGCCATZB3-8aaabbbbGAGTTACCTCCATCCTGTTGCAGAGTCTTGATCTCGTGCTTCZB4-15aaabbbbATGGCCAGCGTGATCTCCCCACCGAATCGAATAACCACRM6089
注:“a”“b”用于区分不同的带型。
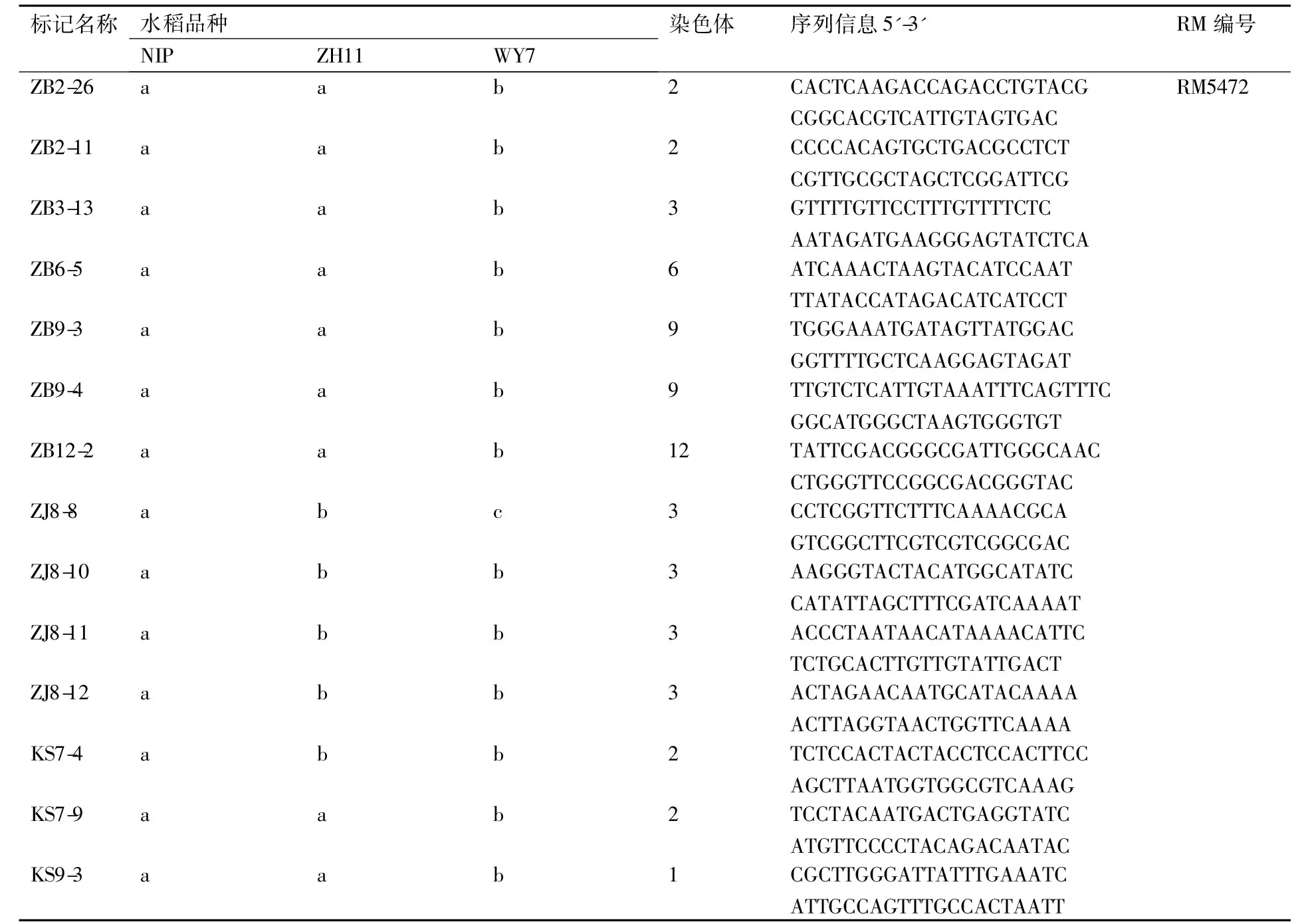
表2部分粳稻间多态性标记序列
Table 2Sequences of several markers polymorphism among japonica rice
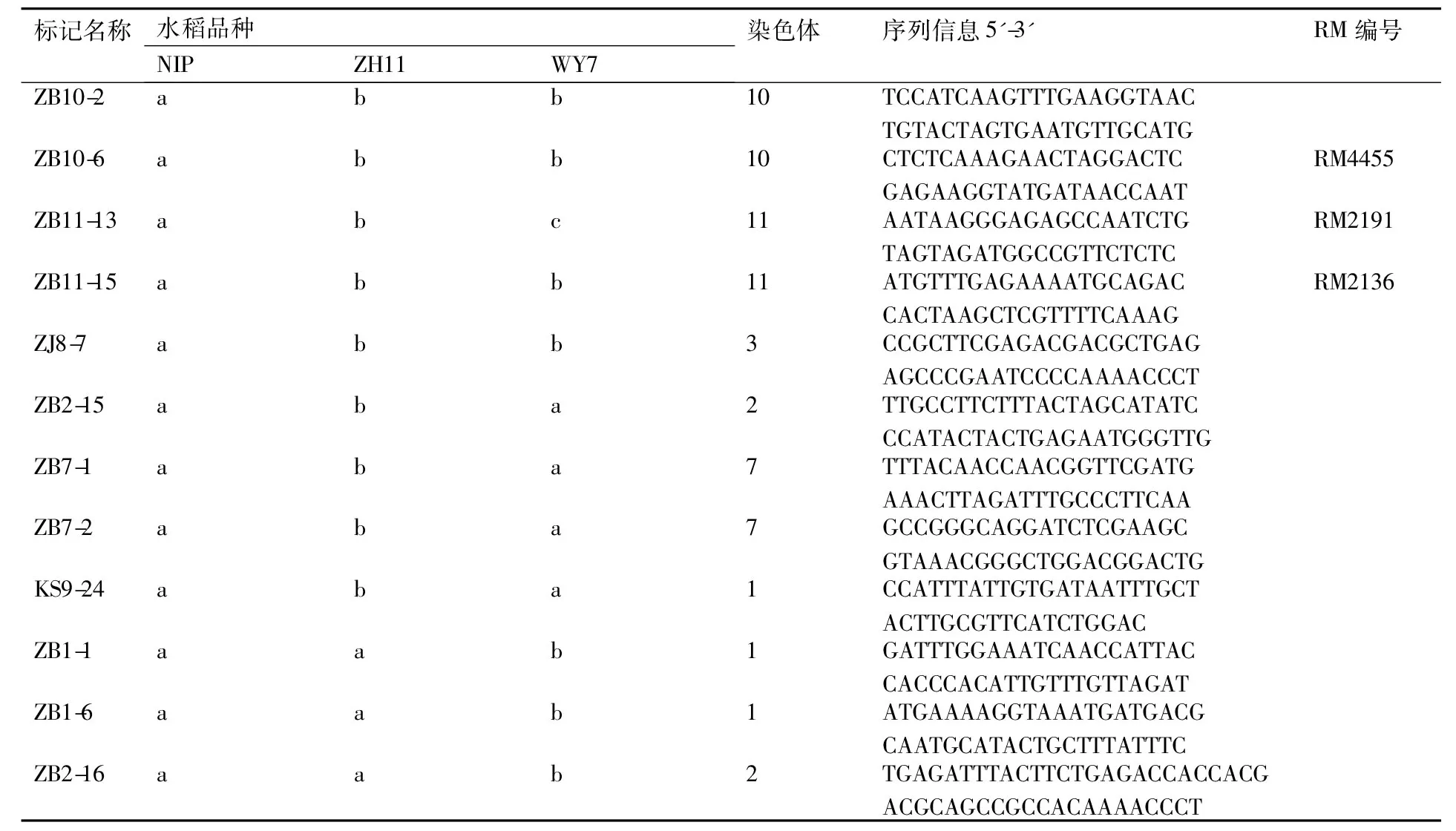
标记名称水稻品种NIPZH11WY7染色体序列信息5'-3'RM编号ZB10-2abb10TCCATCAAGTTTGAAGGTAACTGTACTAGTGAATGTTGCATGZB10-6abb10CTCTCAAAGAACTAGGACTCGAGAAGGTATGATAACCAATRM4455ZB11-13abc11AATAAGGGAGAGCCAATCTGTAGTAGATGGCCGTTCTCTCRM2191ZB11-15abb11ATGTTTGAGAAAATGCAGACCACTAAGCTCGTTTTCAAAGRM2136ZJ8-7abb3CCGCTTCGAGACGACGCTGAGAGCCCGAATCCCCAAAACCCTZB2-15aba2TTGCCTTCTTTACTAGCATATCCCATACTACTGAGAATGGGTTGZB7-1aba7TTTACAACCAACGGTTCGATGAAACTTAGATTTGCCCTTCAAZB7-2aba7GCCGGGCAGGATCTCGAAGCGTAAACGGGCTGGACGGACTGKS9-24aba1CCATTTATTGTGATAATTTGCTACTTGCGTTCATCTGGACZB1-1aab1GATTTGGAAATCAACCATTACCACCCACATTGTTTGTTAGATZB1-6aab1ATGAAAAGGTAAATGATGACGCAATGCATACTGCTTTATTTCZB2-16aab2TGAGATTTACTTCTGAGACCAC-CACGACGCAGCCGCCACAAAACCCT
续表2
注:“a”“b”“c”分别表示不同的带型。下表同。
表3部分籼稻间多态性标记序列
Table 3Sequences of several markers polymorphism among indica rice
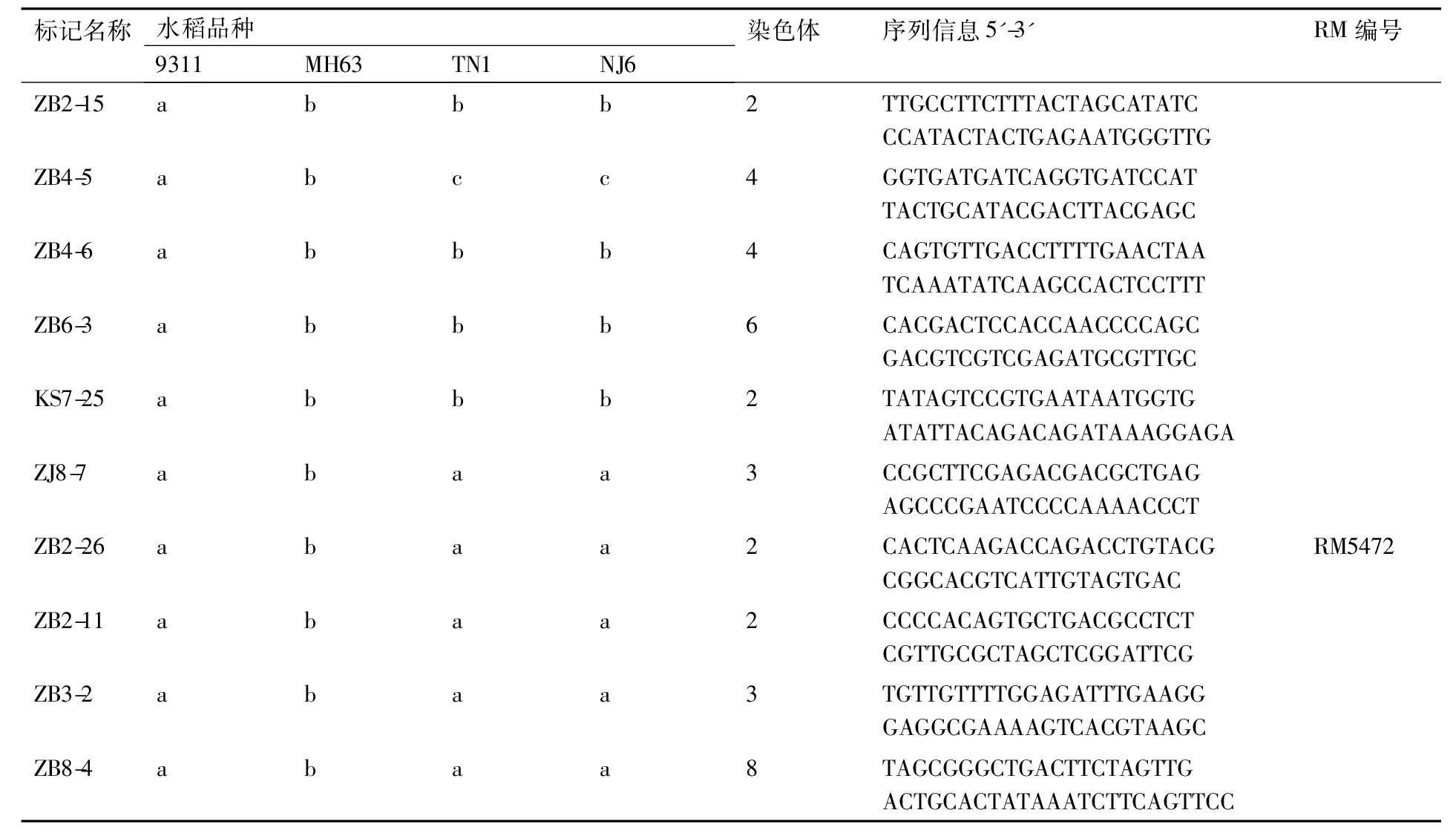
标记名称水稻品种9311MH63TN1NJ6染色体序列信息5'-3'RM编号ZB2-15abbb2TTGCCTTCTTTACTAGCATATCCCATACTACTGAGAATGGGTTGZB4-5abcc4GGTGATGATCAGGTGATCCATTACTGCATACGACTTACGAGCZB4-6abbb4CAGTGTTGACCTTTTGAACTAATCAAATATCAAGCCACTCCTTTZB6-3abbb6CACGACTCCACCAACCCCAGCGACGTCGTCGAGATGCGTTGCKS7-25abbb2TATAGTCCGTGAATAATGGTGATATTACAGACAGATAAAGGA-GAZJ8-7abaa3CCGCTTCGAGACGACGCTGAGAGCCCGAATCCCCAAAACCCTZB2-26abaa2CACTCAAGACCAGACCTGTACGCGGCACGTCATTGTAGTGACRM5472ZB2-11abaa2CCCCACAGTGCTGACGCCTCTCGTTGCGCTAGCTCGGATTCGZB3-2abaa3TGTTGTTTTGGAGATTTGAAGGGAGGCGAAAAGTCACGTAAGCZB8-4abaa8TAGCGGGCTGACTTCTAGTTGACTGCACTATAAATCTTCAGT-TCC
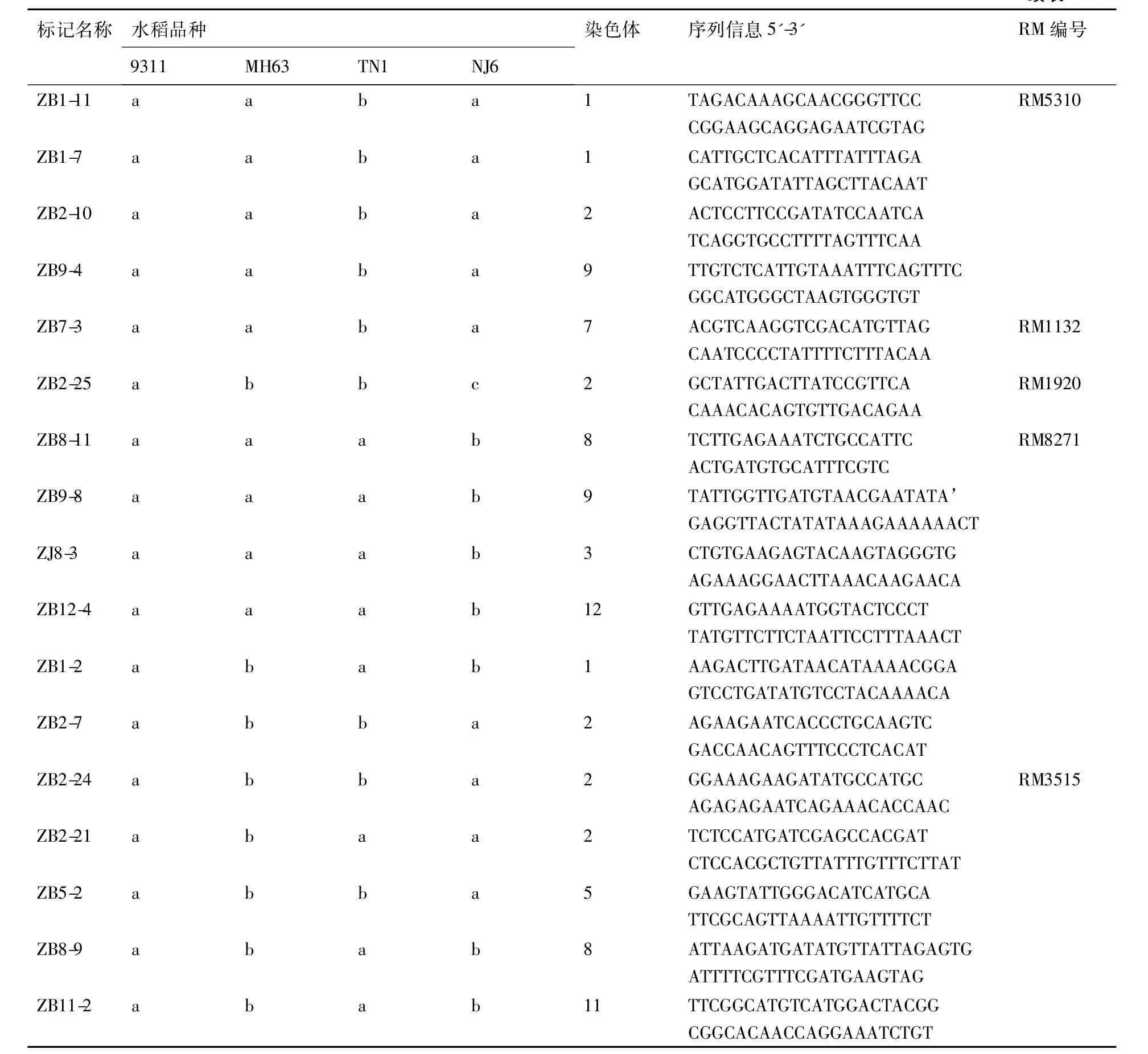
续表3
3 结论与讨论
水稻经过长期的自然演化和人工选择产生了丰富多样并各具特色的种质资源,为高产、优质、抗逆等新品种培育提供了重要的遗传基础。传统的表型分析受自然环境、栽培方式等条件的影响,无法稳定、准确地判断种质遗传背景。分子标记以DNA为模板,不受外界环境影响,判断的稳定性和准确性可大幅提高[15-17]。但是少量的多态标记很难细致区分不同品种间高度同源的序列,因此发掘更多的分子标记,尽可能多地体现不同品种间的多态性,有着非常重要的意义,表型分析与多个分子标记辅助鉴定相结合,将会使不同稻种资源遗传相似性鉴定更为准确、细致[18-21]。虽然水稻全基因组测序工作已经完成,但对于非测序品种来说也只能提供序列参考,与重测序技术高昂的成本相比,用现有序列提供的信息,通过PCR技术发掘并验证更多的多态性分子标记则更为简便易行。本研究通过发掘和验证新的分子标记在3个粳稻品种和4个籼稻品种间的多态性,筛选到92对在籼粳间存在差异的标记,34对粳稻品种间存在差异的标记及80对在籼稻品种间存在差异的标记。除了本研究中选用的7个水稻材料,通过筛选更多的品种,可使这些新开发的标记广泛应用于种质鉴定、进化分析、遗传多样性检测、分子标记辅助选育等领域,应用前景广阔。
表4多态位点位于基因编码区的标记及对应基因
Table 4Markers polymorphism in the coding region and the corresponding genes
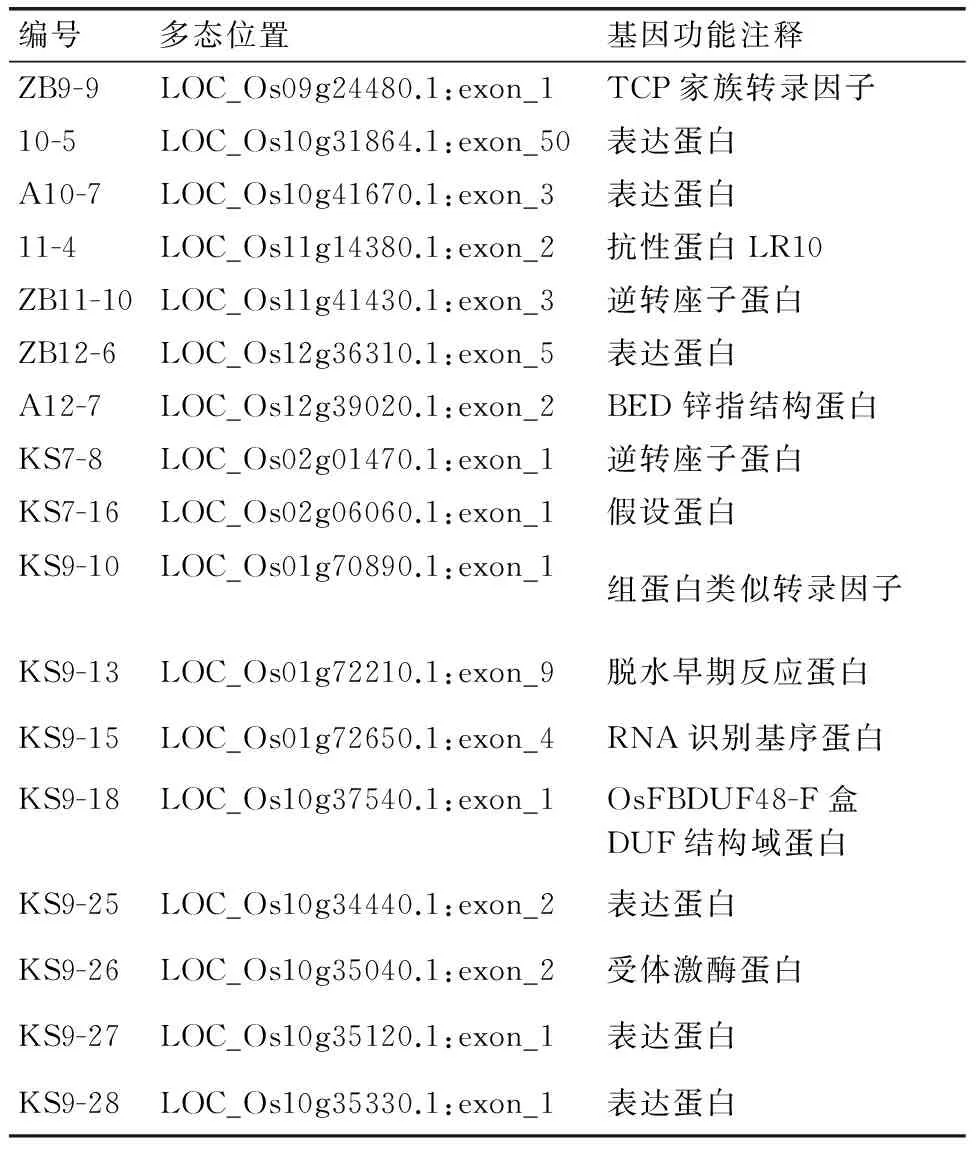
编号多态位置基因功能注释ZB9-9LOC_Os09g24480.1:exon_1TCP家族转录因子10-5LOC_Os10g31864.1:exon_50表达蛋白A10-7LOC_Os10g41670.1:exon_3表达蛋白11-4LOC_Os11g14380.1:exon_2抗性蛋白LR10ZB11-10LOC_Os11g41430.1:exon_3逆转座子蛋白ZB12-6LOC_Os12g36310.1:exon_5表达蛋白A12-7LOC_Os12g39020.1:exon_2BED锌指结构蛋白KS7-8LOC_Os02g01470.1:exon_1逆转座子蛋白KS7-16LOC_Os02g06060.1:exon_1假设蛋白KS9-10LOC_Os01g70890.1:exon_1组蛋白类似转录因子KS9-13LOC_Os01g72210.1:exon_9脱水早期反应蛋白KS9-15LOC_Os01g72650.1:exon_4RNA识别基序蛋白KS9-18LOC_Os10g37540.1:exon_1OsFBDUF48-F盒DUF结构域蛋白KS9-25LOC_Os10g34440.1:exon_2表达蛋白KS9-26LOC_Os10g35040.1:exon_2受体激酶蛋白KS9-27LOC_Os10g35120.1:exon_1表达蛋白KS9-28LOC_Os10g35330.1:exon_1表达蛋白



图4 部分品种间多态性标记的测序结果Fig.4 Sequencing validations of several markers polymorphism among several rice cultivars
在全部可正常扩增而且在不同材料中有多态的194对标记中,在籼粳间、粳稻品种间及籼稻品种间存在差异的标记有206对,超过了194对的标记总数,这是由于部分标记在籼稻和粳稻中各自存在差异,为了便于分类使用,在统计过程中重复统计所致,如图5中的ZB9-4,ZB9-8,ZB6-5等,在粳稻间、籼稻间各自存在差异。为了验证多态的正确性,对一些标记进行测序,结果表明,部分在电泳图中表现出的微弱差别,可能是由于电泳条件的影响,如KS7-4在电泳图中NIP比MH63产物略大,而测序结果却表明二者在序列上没有差异;同样8-8在电泳图中各个品种的产物大小顺序为WY7>NIP=9311>ZH11=MH63,测序结果却表明,WY7与NIP的产物大小一样,均比9311多了6个碱基,相邻的泳道更容易区分产物的大小差异(图4)。一些标记在不同品种间的差异非常大,如3-5在9311和NIP之间有16个碱基的差异,但在NIP和WY7之间有266个碱基的差异,这种类型的标记在NCBI比对同源序列的过程中通常会被自动打断,因此在设计分子标记时考虑两段比对序列接头处的多态,往往也能获得多态性非常好的标记。通过测序还发现,部分序列在非常邻近的序列有多个多态位点,如ZB5-2在9311和NIP之间,多态位点附近600 bp的序列中有2个稳定的序列插入和缺失差异;ZB6-3多态位点附近的序列在NIP,9311和NJ6之间出现了多处插入缺失变异及SNP变异(图4),这些序列可能是水稻进化过程中变异频率比较高的位置,可用于区分多个品种间的差异。
不同水稻品种间农艺性状的差异,本质上是由基因变异所致,因此利用DNA分子标记鉴定不同品种的基因型,可为指导杂交亲本选配、开展籼粳稻亚种间杂种优势的利用、优质抗病等定向遗传改良等方面提供参考。此外,分子标记辅助育种将基因型鉴定和表型鉴定相结合,提高了水稻育种的目的性和中间材料的筛选效率,在生产上具有非常重要的应用价值。近年来的研究结果表明,现有的分子标记在籼粳稻亚种间多态性较好,但在亚种内多态性较低[18,24-25]。王林友等[24]用19对InDel标记对48个籼稻、粳稻及中间型材料进行籼粳属性分析后发现,由于亚种内品种遗传相似性较高,仅用19对标记进行分析会造成不同品种的误判概率升高,需要开发更多更准确的标记才能精确区分亚种内的差别。本研究新开发的标记中有34对标记可用于区分粳稻亚种间的差别,80对标记可用于区分籼稻亚种间的区别,用这些标记对更多亚种进行多态性分析,可大幅提高亚种内品种的辨识率,为分子标记的在新品种选育方面的广泛使用提供参考。

图5 不同品种间部分多态标记的PCR检测结果Fig.5 PCR results of several polymorphism markers in different cultivars
[1]AKAGI H,YOKOZEKI Y,INAGAKI A,et al.Microsatellite DNA markers for rice chromosomes [J].Theoretical and Applied Genetics,1996,93(7):1071-1077.
[2]WU J,MAEHARA T,SHIMOKAWA T,et al.A comprehensive rice transcript map containing 6591 expressed sequence tag sites [J].The Plant Cell Online,2002,14(3):525-535.
[3]ZHANG N,XU Y,AKASH M,et al.Identification of candidate markers associated with agronomic traits in rice using discriminant analysis [J].Theoretical and Applied Genetics,2005,110(4):721-729.
[4]赵中秋,郑海雷,张春光.分子标记的发展及其在植物研究中的应用 [J].生命科学研究,2000,4(2):68-72.
[5]闫华超,高岚,李桂兰.分子标记技术的发展及应用[J].生物学通报,2006,41(2):17-19.
[6]严松,黄福灯,李春寿,等.产量相关性状分子标记遗传距离与籼稻杂种优势的相关性[J].浙江农业学报,2010,22(5):545-551.
[7]PROJECT I R G S.The map-based sequence of the rice genome [J].Nature,2005,436(7052):793-800.
[8]HUANG X,KURATA N,WEI X,et al.A map of rice genome variation reveals the origin of cultivated rice [J].Nature,2012,490(7421):497-501.
[9]GAO Z Y,ZHAO S C,HE W M,et al.Dissecting yield-associated loci in super hybrid rice by resequencing recombinant inbred lines and improving parental genome sequences [J].Proceedings of the National Academy of Sciences,2013,110(35):14492-14497.
[10]SAKAI H,KANAMORI H,ARAI-KICHISE Y,et al.Construction of pseudomolecule sequences of the aus rice cultivar kasalath for comparative genomics of Asian cultivated rice [J].DNA Research,2014,21(4):1131-1142.
[11]刘艳,王逸超,樊继伟,等.分子标记辅助选择Xa23基因在选育抗白叶枯病水稻新品系中的应用[J].浙江农业学报,2011,23(2):248-251.
[12]曾正明,况浩池,罗俊涛,等.利用分子标记辅助筛选改良‘泸恢602’稻瘟病抗性的研究[J].浙江农业学报,2012,24(1):66-70.
[13]邓其明 王世全 郑爱萍,等.利用分子标记辅助育种技术选育高抗白叶枯病恢复系[J].中国水稻科学,2006,20(2):153-158.
[14]袁力行.利用RFLP,SSR,AFLP和RAPD标记分析玉米自效系遗传多样性的比较研究[J].遗传学报,2000,27(8):725-733.
[15]刘艳华,牟建民,王志德,等.分子标记技术在烟草遗传育种中的应用[J].植物遗传资源学报,2007,8(1):118-122.
[16]王黎明,焦少杰,姜艳喜.利用分子标记分析高粱的遗传多样性及其在种质创新中的应用[J].植物遗传资源学报,2015,16(2):288-293.
[17]OLUFOWOTE J O,XU Y,CHEN X,et al.Comparative evaluation of within-cultivar variation of rice (Oryza sativa L.) using microsatellite and RFLP markers [J].Genome,1997,40(3):370-378.
[18]李小湘,肖军治,段永红,等.湖南同名地方稻资源SSR标记及表现型的比较分析[J].植物遗传资源学报,2014,15(2):248-254.
[19]贾继增.分子标记种质资源鉴定和分子标记育种[J].中国农业科学,1996,29(4):1-10
[20]高宁,景蕊莲,陈耀锋,等.作物抗旱相关分子标记及其辅助选择的研究进展[J].植物遗传资源学报,2003,4(3):274-278.
[21]徐云碧,申宗坦,朱立煌,等.水稻形态性状与分子标记的相互关联及其检测[J].浙江农业学报,1994,6(1):1-6.
[22]SHEN Y J,JIANG H,JIN J P,et al.Development of genome-wide DNA polymorphism database for map-based cloning of rice genes [J].Plant Physiology,2004,135(3):1198-1205.
[23]卢扬江,郑康乐.提取水稻 DNA的一种简易方法[J].中国水稻科学,1992,6(1):47-48.
[24]王林友,张礼霞,勾晓霞,等.利用InDel标记鉴定水稻育种材料的籼粳属性[J].核农学报,2013,27(7):913-921.
[25]冯芳君,罗利军,李荧,等.水稻 InDel和SSR标记多态性的比较分析[J].分子植物育种,2006,3(5):725-730.
(责任编辑侯春晓)
The development and verification of nucleotide polymorphism markers in rice
QIU Zhen-nan1,2,XU Qian-kun1,WANG Xiao-qi2,ZHAO Juan2,HE Lei2,ZHANG Sen2,MA Bo-jun1,QIAN Qian2,ZHU Li2,*
(1.College of Chemistry and Life Sciences,Zhejiang Normal University,Jinhua 321004,China;2.State Key Laboratory of Rice Biology,China National Rice Research Institute,Hangzhou 310006,China)
In this work,polymorphic analysis among 3 japonica cultivars (NIP,ZH11 and WY7) and 4 indica cultivars (9311,MH63,TN1 and NJ6) were conducted with 203 molecular markers,92 polymorphism markers between japonica cultivars and indica cultivars,34 polymorphism markers in japonica cultivars and 80 polymorphism markers in indica cultivars had been screened.Sequencing analysis results showed that the polymorphism of microsatellite markers among different varieties was mainly caused by the different repetitions of repetitive sequence.However,the polymorphism of insertion/deletion marks was determined by the repetition times of non-simple repetitive sequence and insertion or deletion of long fragment.Besides the development of new markers,several RM markers also had been verified whether possessed polymorphism in different varieties.Taken together,these new polymorphism markers were bound to be of great service to use in germplasm identification,evolution,genetic diversity detection and molecular marker-assisted selection.
rice;molecular markers;polymorphism;cultivar identification
浙江农业学报Acta Agriculturae Zhejiangensis,2016,28(3):361-370http://www.zjnyxb.cn
邱振楠,徐乾坤,王小琦,等.水稻DNA多态标记的开发与验证[J].浙江农业学报,2016,28(3):361-370.
10.3969/j.issn.1004-1524.2016.03.01
2015-08-10
国家自然科学基金面上项目(31171532);浙江省“转基因农作物新品种培育”科技创新团队项目(2011R50021);中央级公益性科研院所专项资金项目(2015RG001-2)
邱振楠(1989—),男,山东德州人,硕士研究生,研究方向为水稻遗传育种。E-mail:zhnanqiu@126.com
,朱丽,E-mail:zhuli05@caas.cn
S511
A
1004-1524(2016)03-0361-10
猜你喜欢
古今农业(2022年2期)2022-08-15
安徽农学通报(2022年8期)2022-05-06
今日农业(2021年15期)2021-10-14
农民致富之友(2020年32期)2020-12-03
新世纪智能(高一语文)(2020年12期)2020-06-01
江西农业(2018年9期)2018-02-11
河南农业(2017年7期)2017-07-25
科技视界(2016年27期)2017-03-14
浙江农业科学(2016年11期)2016-05-04
中国医药导报(2015年27期)2015-02-28
