
G蛋白偶联受体家族味觉受体的信号转导及应用研究进展
2016-11-11 07:34朱洲海曾婉俐黄海涛李雪梅夭建华
食品科学 2016年3期
高 茜,朱洲海,曾婉俐,黄海涛,周 岚,李雪梅,夭建华*
(云南中烟工业有限责任公司技术中心,云南 昆明 650106)
G蛋白偶联受体家族味觉受体的信号转导及应用研究进展
高 茜,朱洲海,曾婉俐,黄海涛,周 岚,李雪梅,夭建华*
(云南中烟工业有限责任公司技术中心,云南 昆明 650106)
味觉是口中的物质与味觉受体细胞发生反应而产生的一种感觉。当呈味物质刺激味觉受体时,信号通过神经感觉系统传导到大脑,经过综合神经中枢系统的分析,从而产生味觉。近年来味觉受体、信号转导及应用研究引起了极大的关注,对细胞的大量研究结果表明不同的味觉通过不同的味觉受体产生。本文就G蛋白偶联受体(guanosine-binding protein coupled receptor,GPCR)家族味觉受体的信号转导过程及应用研究的进展进行综述。
味觉受体;信号转导;应用
味觉是哺乳动物最为重要的感觉之一,它指引动物在摄取营养的同时避免有害毒素及无法消化的物质。味觉由甜、酸、苦、咸、鲜这5 种基本味感组成,近年来研究表明还存在着第六种味觉,脂味觉。甜味预示着作为能量来源的碳水化合物的存在,咸味表示钠盐和其他盐类的摄入,鲜味可能是反映食物中蛋白质的含量,苦味可以帮助识别自然界中潜在的致命的有毒有害物质,而酸味是为了避免过多的酸性物质从而维持体内的酸碱平衡。尽管如此,人类在进化中学会了忍受甚至是寻求一些苦的或酸的化合物,例如咖啡和柠檬酸。脂肪味与哺乳动物的肥胖密切相关,这种味觉与人类爱吃洋芋片、巧克力等高脂食品的相关。味觉受体基因多态性导致了偏好口味的多样化,从而影响食物的选择及营养物质的摄取[1]。
味觉是由存在于味蕾上的味觉受体细胞(taste receptor cell,TRC)表面的味觉受体蛋白介导产生的。当这些受体蛋白与相应的味觉分子(配体)结合后,便产生了兴奋性冲动,这种冲动通过末梢神经传入中枢神经,于是便会感受到不同的味道。近年来,味觉受体、信号转导及调节机制引起了极大的关注。第一个味觉受体基因T1R1(taste receptor family 1 member 1)发现于1999年,是Hoon等[2]在对小鼠舌头上的味觉细胞进行cDNA文库测序时发现,随后根据T1R1基因为基础又鉴定出T1R2基因。2001年,4 个研究小组研究人员在人类第4对染色体上鉴别出了一个与感觉甜味有关的基因T1R3,从而确定了第一个可能的甜味受体[3-6]。近年来随着研究的深入,人们对味觉受体的组成及机制的理论基础有了较为全面的了解,与之相应的应用研究也得到了广泛的关注。G蛋白偶联受体(G protein coupled receptor,GPCR)是细胞信号传导中的重要蛋白质,是已知的一大类涉及跨膜信号传导且具有7 次跨膜保守结构的膜受体。甜味、鲜味和苦味受体已经被证实是GPCR家族成员,这3 种味觉信号传导机制的研究比较全面和详细。脂肪味是近年来才被关注的一类味觉,目前的候选受体是GPR120和CD36(cluster of differentiation 36),GPR120也是属于GPCR家族成员,属于进化保守的视紫红质家族受体。酸味受体候选基因PKD1L3(polycystin kidney dominant 1 like 3)和PKD2L1(polycystin kidney dominant 2 like 1)属于瞬时受体电位(transient receptor potential,TRP)通道成员[7],咸味受体主要有上皮细胞钠离子通道(epithelial Na+channel,ENaC)[8]和瞬时受体电位香草酸亚型1(transient receptor potential vanilloid1,TRPV1)[9]两种,酸味和咸味受体的遗传机制和功能研究目前报道都比较少,本文主要综述了GPCR家族味觉受体的信号转导及味觉受体的应用研究。
1 GPCR家族味觉受体的信号转导的分子机制
在口腔中,甜、鲜和苦味化合物激活不同的受体细胞上面的味觉GPCRs受体(图1),T1Rs(taste receptor type 1)是Ⅲ类GPCRs的二聚体,具有很大的N末端胞外结构域,这些结构域形成一个捕蝇草结构,其中包括捕蝇草结构域(venusflytrapdomain,VFT)、富半胱氨酸结构域(cysteine-rich domain CRD)和七螺旋跨膜结构域(heptahelical transmembrane domain,HD)[10]。VFT上具有多种配体结合位点,负责与配体的识别与结合。HD上的细胞内环和跨膜区段是G蛋白结合位点,因而高度保守;而细胞外环为配体结合区,在各家族成员间差异较大,正因为这种多态性,尽管味觉在哺乳动物中,甚至在更广泛的生物中具有相似的细胞信号传递途径,但是不同的动物味觉差异很大。
T1R1、T1R2和T1R3形成异源二聚体后具有味觉受体的功能(图1和图2),而不同的组合方式所产生的味觉受体对应于不同的味觉(图2)。
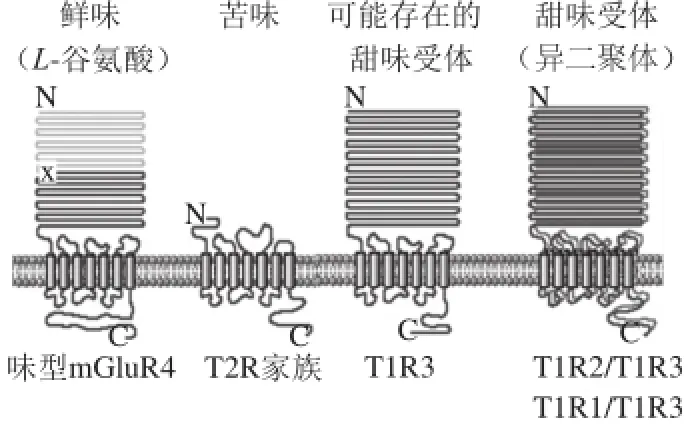
图1 苦味、甜味和鲜味受体结构示意图[18]Fig.1 Primary structures of taste receptors[18]
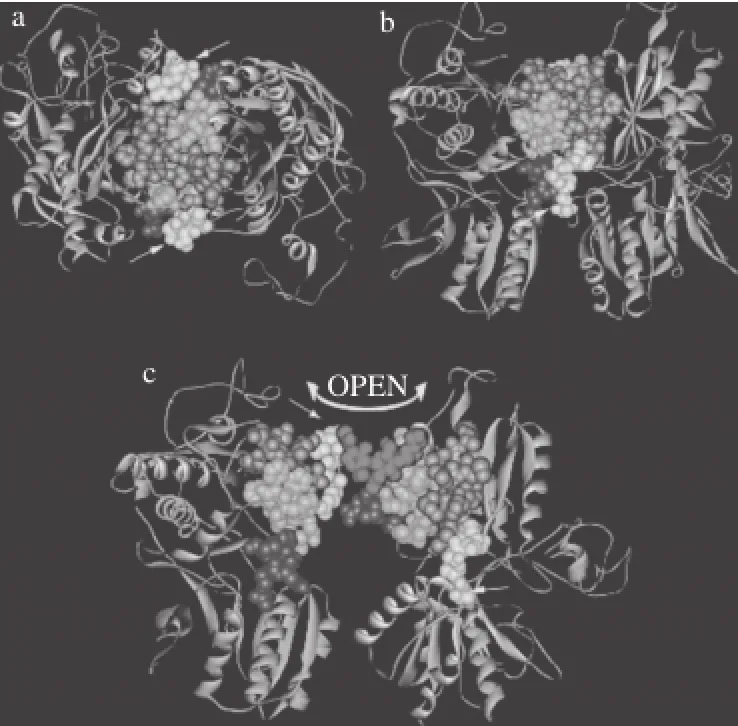
图2 T1R的三维结构示意图[24]Fig.2 Three-dimensional structure of T1R[24]
表达T1R2+T1R3的受体细胞能感受天然甜味剂(如蔗糖、甜菊糖等)及人工甜味剂(如糖精、阿斯巴甜等)的刺激。一些T1R3基因敲除的小鼠对一些糖类仍有反应,暗示着可能有其他甜味受体的存在,但是这一结果还没有被确认[11]。不同物种对甜味剂的识别不尽相同,如小鼠不能识别部分人工甜味剂如阿斯巴甜[12]。Jiang Peihua等[13]在2012年发现突变破坏了海狮、海狗、太平洋斑海豹、亚洲小爪水獭、斑鬣狗、马达加斯加长尾灵猫、条纹林狸和瓶鼻海豚的T1R2基因,这可能与它们进食的特殊进化直接相关,因为它们不通过咀嚼就将食物整个吞下去。
表达异源二聚体T1R1+T1R3的受体细胞响应鲜味的刺激,特别是对谷氨酸盐和肌苷-5-单磷酸(inosine 5-monophosphate,IMP)/鸟嘌呤核苷酸(guanosine 5-ribonucleotide,GMP)反应强烈[14-15],这些化合物大量存在于鱼、贝类、咸肉、蔬菜等食物中。对T1R1和T1R3的非同义单核苷酸突变研究表明,T1R1的N末端配体结构域的两个氨基酸的替换及T1R3跨膜结构域的两个氨基酸的替换,都会对受体响应谷氨酸钠的刺激产生严重的影响。这为不同人群之间对谷氨酸钠的敏感性差异提供了一个可能的解释[16]。鲜味受体和甜味受体共享一个T1R3,但能识别不同的味道,说明T1R1和T1R2的N末端胞外域决定了鲜甜受体的配体特异性。然而在T1R3敲除的小鼠中仍然对鲜味有明显的反应,味蕾上的不同细胞亚群和神经反应表明对于不同的鲜味物质也有多种识别模式,这些结果表明对于鲜味的识别可能也存在其他的受体[17]。总之,虽然T1Rs成员可以识别甜味和鲜味,但可能有其他的GPCRs也起着补充的作用。
T2Rs类似于Ⅰ类G蛋白偶联受体,同T1Rs一样,也是在跨膜螺旋处具有保守的G 蛋白结合位点,而胞外区则是与苦味配体结合的区域,具有明显的多态性[19]。与一般GPCR家族蛋白不同的是,T2Rs具有较短的N末端结构域。表达T2R家族的细胞识别苦味化合物,该家族已鉴定有20~35 个成员,大多数基因都在染色体上连锁存在。不同的苦味受体识别范围差异较大,有些特异性的结合2~4 种苦味化合物,另一些能被很多种配体所激活[20],单个味觉受体细胞内可能表达多个苦味受体,任何一个苦味受体被激活,连接该细胞和脑的神经均可获得相同的信息[21]。T2R家族的表达模式和整个基因家族的多态性使人和动物能辨别自然界中广泛而多样的有毒苦味物质[22],2012年Clark等[23]发现由于许多药物味微苦,如氯喹、氟哌啶醇、红霉素、普鲁卡因胺、氧氟沙星等,这些药物可以与口腔、胃肠道及呼吸系统的T2R家族受体结合,这也为不同药物的脱靶效应提供了一个新的假设。
Gustducin是一种特异性的味觉信号偶联蛋白,同其他G蛋白一样,也由α、β、γ 3 个亚基组成,当甜味、鲜味和苦味的味觉受体结合配体后,α-Gustducin被激活[25]。除了Gustducin之外,味觉受体细胞中还存在一些其他G蛋白的α亚基,如Gαi-3、Gα14、Gα15等[26]。味觉信号的传导通过G蛋白的β和γ亚基,包括Gγ13和Gβ1或者Gβ3[27]。当受体在与配体结合后,G蛋白的β和γ亚基从G蛋白偶联受体上释放出来,Gα活化磷酸二酯酶(phosphodiestera se,PDE),水解环磷酸腺苷(cyclic adenosine monophosphate,cAMP),解除了环单磷酸核苷酸(cyclic nucleotide monophosphates,cNMP)对离子通道的抑制,钙离子浓度升高,导致膜的去极化和神经递质的释放。Gβγ直接与PLCβ2结合[26]。PLCβ2的敲除会导致味觉感受严重削弱,但是不会完全丧失味觉[28]。PLCβ2激活磷脂酰肌醇磷酸(phosphatidylinositol biphosphate,PIP2)产生甘油二酯(diacylglycerol,DAG)和肌醇三磷酸(inositol triphosphate,IP3),IP3与三磷酸肌醇受体3(inositol triphosphate receptor 3,IP3R3)结合,导致胞内细胞器膜上IP3门控钙离子通道的开放,胞内贮存的钙离子被释放出来,引起胞质内钙离子浓度上升(图3)。这些细胞内 增加的钙离子引起TRPM5通道开放,导致细胞膜的去极化和神经递质的释放[29]。
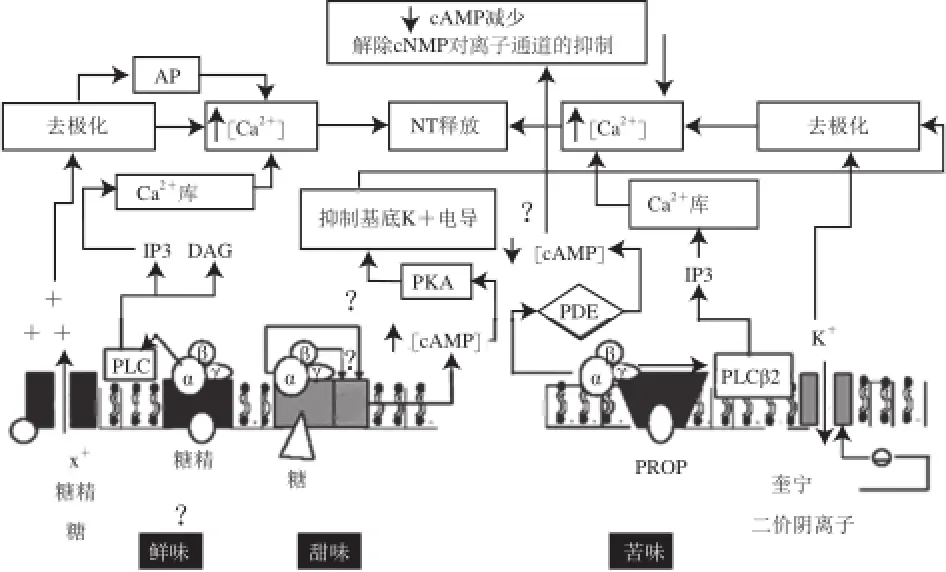
图3 苦味、甜味和鲜味信号传导机制示意图[24,35]Fig.3 Signal transduction mechanisms of bitter, sweet and umami tastes (modified from previous data)[24,35]
虽然在甜、鲜、和苦味物质传导过程中,通过Gustducin介导的通路被认为是主要的信号通路,但是其他一些实验结果表明,可能存在其他G蛋白或者其他信号通路来传导味觉。最有力的证据来自敲除了Gustducin基因的突变型小鼠,与野生型相比较,这些小鼠对于苦味物质的反应只是减少,并不会完全丧失[25]。除去经典的神经递质,味蕾细胞还可表达部分神经肽类,如血管活性肠肽、胆囊收缩素(cholecystokinin,CCK)、神经肽Y、胰高血糖素样肽-1(glucagon like peptide-1,GLP-1)等,同样发挥神经递质的作用[30]。
近年来,研究人员发现另外一类GPCR类特异性受体,可与游离脂肪酸(free fatty acids,FFAs)相结合,包括GPR40、41、43、84和120[31]。其中GPR120是长链不饱和FFA的受体,属于GPCRs超家族中的视紫红质样受体家族。该受体在人及啮齿类动物的组织中表达非常广泛,如脑、垂体、心脏、肺、胃、肠道和脂肪组织中均有表达[32]。同时在小鼠、大鼠及人的味蕾细胞及周围的上皮细胞中,GPR120也有丰富的表达,GPR120基因敲除小鼠对亚油酸和油酸的选择减少,对这些脂肪酸的味觉神经反应减弱[33],这些提示其可能参与了人对脂肪酸的味觉感受。目前的研究表明,GPR120的配体主要为多不饱和长链脂肪酸,其他类型的配体如为奇果菌素衍生物、灰叶酸和灰叶酸甲醚也可与GPR120结合[34],这些新型配体的发现为研究GPR120的生理功能和分子机制提供了研究手段。在表达GPR120的小鼠肠L细胞株STC-1的研究中发现,GPR120与配体结合后与甜、苦、鲜味的分子机制类似,通过激活PIP2产生DAG和IP3,从而引发胞内钙信号,调控GLP-1的释放[32]。
2 味觉应答机理
味觉受体细胞和突触前细胞释放不同的神经递质,受体细胞释放ATP,以前认为ATP的释放可能是通过缝隙连接蛋白家族成员pannexin和connexin通道蛋白,但是最新 的研究表明味觉感受细胞中的钙稳态调节蛋白1(calcium homeostasis modulator 1,CALHM1),是感知甜味、苦味和鲜味时必不可少的新型离子通道蛋白。2013年来自9 所研究机构的科学家们联合发现敲除CALHM1基因的小鼠无法释放ATP来给大脑传递甜味、苦味和鲜味的信息,但对酸味和咸 味的感知则基本正常[36]。而突触前细胞分泌5-羟色胺(5-hydroxytryptamine,5-HT)和去甲肾上腺素(norepinephrine,NE)。这些生物胺的分泌可能是通过传统的钙依赖的胞吐[37]。
味觉过程是一个定量化的神经传感过程,不同浓度的味觉成分会获得不同的味觉感受,而且这在不同的动物,甚至同一种动物的不同个体之间都有明显差异。当甜、苦或鲜味物质刺激味蕾时,味觉受体细胞释放ATP刺激味觉传入神经纤维,同时ATP刺激相邻的突触前细胞释放5-HT和/或NE。ATP还可刺激味觉受体细胞本身产生正反馈,增加ATP的分泌从而抵消ATP酶对ATP的降解[38]。突触前细胞分泌的5-HT也具有多重作用,其中的一个作用是抑制释放ATP的味觉受体细胞,作为味觉受体细胞的一个负反馈,这种负反馈会降低味觉信号的输入从而导致感觉适应。在接收味觉刺激后这种正反馈和负反馈联合塑造从味蕾传入脑的信号[39]。然而,这些反馈的途径究竟是如何协调平衡的还需要进一步的实验证实。有可能5-HT可能承担一个侧面抑制的作用,例如当某一个特定味觉受体(如甜味受体)被激活的时候,5-HT可能抑制与之相邻的受体(如苦味受体)的信号输出。5-HT的其他作用位点可能包括突触细胞和突触前细胞的神经纤维。突触前细胞产生的初级信号只有咸味物质,当突触前细胞出问题时这是唯一会缺失的味觉[40]。对甜、苦和鲜味这几种味觉来说,突触前细胞的次级反应是否是味觉感受所必需的仍有争论。
味觉传入神经纤维将信号从味蕾传至大脑。至于受体细胞和突触前细胞在受到味觉刺激时是如何将这些不同的神经感受分别开来仍不清楚。关于这个问题有两种不同的意见,一种认为不同的味道有其专属的神经纤维,如苦味有苦味的神经纤维。另一种则认为多个神经纤维形成一个整合的系统,任何单一味道的刺激都能引起多个神经纤维的信号传导[18]。电生理记录表明一些神经元对某些味道(如甜味)具有强烈的反应,但对某些其他的味道(如苦味)也有微弱的反应,但是有些神经元对多种味道都有强烈的反应,这些神经元的活动模式显示了味蕾细胞反应具有异质性[41-43]。总之,虽然多种味道都通过ATP这一神经递质传入信号,但是这些信息如何整合成为最终的味觉感受还需要研究。因此对于味觉受体信号转导通路来说,仍然存在许多悬而未决的问题。
3 味觉受体的应用研究
随着味觉受体的研究逐渐深入,如何将这些研究结果应用到实际中成了一个新的热点。全球食品香料的领军企业,美国Senomyx公司构建了能够稳定表达T1R2/T1R3的细胞系,并在此基础上开发出了一套用于甜味剂高通量筛选的平台SweetScreenHT,作为其核心技术开发出了多种人工甜味剂。Senomyx公司于2010年已和可口可乐公司签署了协议,利用其基于甜味受体细胞的甜味剂高通量筛选技术,为可口可乐公司开发新型甜味剂,用于无糖汽水的生产。这一技术是否可应用于其他行业的甜味剂筛选如卷烟烟气的甜韵感分析,还有待进一步研究。
了解味觉与情绪、食欲、肥胖和饱腹感的联系可以揭示人类多种生理病理反应机制,从而开发更多食品药品领域的新方法新技术。味觉受体作为营养传感系统发挥作用,可以作为很多代谢综合征治疗药物靶点,是由于这些病源自现代人对营养,特别是能量物质的过度摄入、吸收和积累所造成。2014年来自日本东京工业大学和广岛大学的研究人员发现,人对某种食物的味觉感受会反映在脸部皮肤的血流量上,可以通过检测脸部血流量的变化来判断某人是否喜欢某种口味,这一技术将来可用于为这类患者提供可口食物,还可用于食品开发领域[44]。味觉指引着我们的食物选择,是选择甜、咸还是鲜味,味觉超常敏感者对绿色蔬菜的苦味感受更强烈,导致摄入蔬菜量太少,从而可能导致患结肠癌的风险更高[45]。另外,Kamble等[46]在2013年指出多种药物如抗抑郁药和镇静催眠药都会影响味觉的感受,这些问题的解决将为多种食物及药物的开发起着重要的指引作用。
除了众所周知的口腔以外,味觉受体在其他部位的表达同样有重要的应用研究,2010年来自马里兰大学医学院的研究人员报道称苦味受体不仅存在于口腔中,还存在于人的肺中,他们发现肺中的味觉受体与舌头上的受体结构完全相同,然而在功能上却存在很大差异。舌头上的味觉受体常成簇集中在味蕾,可将味觉信号直接传送至大脑。而肺内的味觉受体并不会成簇积聚,也不会直接向大脑传送信号,而是对带有苦味的物质产生反应[47]。2012年来自南京大学模式动物研究所、温州医学院、麻省大学医学院等处的研究人员发现苦味化合物与肺部的苦味受体结合后可打开气道,其效应甚至强于那些用于治疗哮喘或慢性阻塞性肺疾患的药物。因此可以用于开发出一些治疗哮喘、肺气肿或慢性支气管炎的新药[48]。胃肠道中也表达能识别甜、苦、鲜味的受体蛋白,通过引发多肽类物质的释放从而刺激局部中枢神经反应和(或)迷走神经从而将信号传递至大脑[49-50],2012年比利时鲁汶天主教大学转化研究中心从事胃肠道功能紊乱研究的Janssen和Depoortere在研究报道上指出,当食物摄入后,肠道通过激素的释放控制饱腹感和血糖水平,位于胃部的传感器或受体能对过剩的食物摄取产生响应,如果它们发生了故障,则很可能会导致肥胖、糖尿病和相关代谢性疾病的发展,但还需要进一步的研究证明哪种肠道味觉受体有潜力能够成为预防和治疗肥胖和糖尿病的有效药物靶标[51]。2013年美国Monell化学感觉中心的科学家们的研究成果表明味觉受体蛋白还在睾丸和精子中表达,并在精子的发育中发挥了重要的作用。将味觉受体抑制剂添加到小鼠的饮食中会导致精子畸形及数量减少,小鼠变得不育。而在除去饮食中的抑制剂之后,小鼠的不育快速地得到逆转[52]。
4 结 语
虽然近几年味觉的研究取得了很大的进展,但是还有很多问题没有得到解决。一个主要的问题是其他味觉受体的鉴定。已知的味觉受体并不对所有的甜味和鲜味的配体结合,这表明很可能存在其他的甜味和鲜味的受体。一些其他味道如酸味、脂肪、金属和涩味的转导机制也研究较少。解决这些问题可能要对人或小鼠的群体进行分子生物学、群体遗传分析和蛋白表达研究。另外一个需要研究的领域是味觉信号在神经系统中是如何编码的。我们仍然不知道大脑是如何区分甜、酸、咸等味道。如果味觉不是遵循一个简单的代码,味觉信号是如何发送和破译的仍不得而知。而如何将这些基础研究进一步应用于食品药品等的多种生活领域,仍是一个任重而道远的过程。
[1] SHIGEMURA N, SHIROSAKI S, SANEMATSU K, et al. Genetic and molecular basis of individual differences in human umami taste perception[J]. PLoS ONE, 2009, 4(8): e6717. DOI:10.1371/journal. pone.0006717.
[2] HOON M A, ADLER E, LINDEMEIER J, et al. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity[J]. Cell, 1999, 96(4): 541-551.
[3] NELSON G, HOON M A, CHANDRASHEKAR J, et al. Mammalian sweet taste receptors[J]. Cell, 2001, 106(3): 381-390. DOI:10.1016/ S0092-8674(01)00451-2.
[4] MAX M, SHANKER Y G, HUANG L, et al. Tas1r3, encoding a new candidate taste receptor, is allelic to the sweet responsiveness locus Sac[J]. Nature Genetics, 2001, 28(1): 58-63.
[5] SAINZ E, KORLEY J N, BATTEY J F, et al. Identification of a novel member of the T1R family of putative taste receptors[J]. Journal of Neurochemistry, 2001, 77(3): 896-903. DOI:10.1046/j.1471-4159.2001.00292.x.
[6] MONTMAYEUR J P, LIBERLES S D, MATSUNAMI H, et al. A candidate taste receptor gene near a sweet taste locus[J]. Nature Neuroscience, 2001, 4(5): 492-498.
[7] ISHIMARU Y, INADA H, KUBOTA M, et al. Transient receptor potential family members PKD1L3 and PKD2L1 form a candidate sour taste receptor[J]. Proceedings of the National Academy of Sciences, 2006, 103(33): 12569-12574. DOI:10.1073/pnas.0602702103.
[8] KRETZ O, BARBRY P, BOCK R, et al. Differential expression of RNA and protein of the three pore-forming subunits of the amiloridesensitive epithelial sodium channel in taste buds of the rat[J]. Journal of Histochemistry and Cytochemistry, 1999, 47(1): 51-64. DOI:10.1177/002215549904700106.
[9] LYALL V, HECK G L, VINNIKOVA A K, et al. The mammalian amiloride-insensitive non-specific salt taste receptor is a vanilloid receptor-1 variant[J]. The Journal of Physiology, 2004, 558(1): 147-159. DOI:10.1113/jphysiol.2004.065656.
[10] LI Xiaodong. T1R receptors mediate mammalian sweet and umami taste[J]. The American Journal of Clinical Nutrition, 2009, 90(Suppl 3): 733-737. DOI:10.3945/ajcn.2009.27462G.
[11] DAMAK S, RONG M, YASUMATSU K, et al. Detection of sweet and umami taste in the absence of taste receptor T1R3[J]. Science, 2003, 301: 850-853. DOI:10.1126/science.1087155.
[12] HUANG Y J, MARUYAMA Y, LU K S, et al. Mouse taste buds use serotonin as a neurotransmitter[J]. The Journal of Neuroscience, 2005, 25(4): 843-847. DOI:10.1523/JNEUROSCI.4446-04.2005.
[13] JIANG P H, JOSUE J, LI X, et al. Major taste loss in carnivorous mammals[J]. Proceedings of the National Academy of Sciences, 2012, 109(13): 4956-4961. DOI:10.1073/pnas.1118360109.
[14] LI X, STASZEWSKI L, XU H, et al. Human receptors for sweet and umami taste[J]. Proceedings of the National Academy of Sciences, 2002, 99(7): 4692-4696. DOI:10.1073/pnas.072090199.
[15] NELSON G, CHANDRASHEKAR J, HOON M A, et al. An aminoacid taste receptor[J]. Nature, 2002, 416: 199-202.
[16] RALIOU M, GRAUSO M, HOFFMANN B, et al. Human genetic polymorphisms in T1R1 and T1R3 taste receptor subunits affect their function[J]. Chemical Senses, 2011, 36(6): 527-537. DOI:10.1093/ chemse/bjr014.
[17] YASUMATSU K, HORIO N, MURATA Y, et al. Multiple receptors underlie glutamate taste responses in mice[J]. The American Journal of Clinical Nutrition, 2009, 90(Suppl 3): 747-752. DOI:10.3945/ ajcn.2009.27462J.
[18] CHAUDHARI N, ROPER S D. The cell biology of taste[J]. The Journal of Cell Biology, 2010, 190(3): 285-296. DOI:10.1083/ jcb.201003144.
[19] BACHMANOV A A, BOSAK N P, CAILU L, et al. Genetics of taste receptors[J]. Current Pharmaceutical Design, 2014, 20(16): 2669-2683. DOI:10.2174/13816128113199990566.
[20] MEYERHOF W, BATRAM C, KUHN C, et al. The molecular receptive ranges of human TAS2R bitter taste receptors[J]. Chemical Senses, 2010, 35(2): 157-170. DOI:10.1093/chemse/bjp092.
[21] ADLER J E. T2R taste receptors and genes encoding same: 8334367[P]. 2012-12-18.
[22] DRAYNA D. Human taste genetics[J]. Annual Review of Genomics and Human Genetic, 2005, 6: 217-235. DOI:10.1146/annurev. genom.6.080604.162340.
[23] CLARK A A, LIGGETT S B, MUNGER S D. Extraoral bitter taste receptors as mediators of off-target drug effects[J]. The FASEB Journal, 2012, 26(12): 4827-4831. DOI:10.1096/fj.12-215087.
[24] MARGOLSKEE R F. Molecular mechanisms of bitter and sweet taste transduction[J]. Journal of Biological Chemistry, 2002, 277(1): 1-4. DOI:10.1074/jbc.R100054200.
[25] WONG G T, GANNON K S, MARGOLSKEE R F. Transduction of bitter and sweet taste by gustducin[J]. Nature, 1996, 381: 796-800. DOI:10.1038/381796a0.
[26] CHANDRASHEKAR J, HOON M A, RYBA N J, et al. The receptors and cells for mammalian taste[J]. Nature, 2006, 444: 288-294. DOI:10.1038/nature05401.
[27] LI Feng, ZHOU Minliang. Depletion of bitter taste transduction leads to massive spermatid loss in transgenic mice[J]. Molecular Human Reproduction, 2012, 18(6): 289-297. DOI:10.1093/molehr/gas005.
[28] DOTSON C D, ROPER S D, SPECTOR A C. PLCβ2-independent behavioral avoidance of prototypical bitter-tasting ligands[J]. Chemical Senses, 2005, 30(7): 593-600. DOI:10.1093/chemse/bji053.
[29] HUANG Y A, ROPER S D. Intracellular Ca2+and TRPM5-mediated membrane depolarization produce ATP secretion from taste receptor cells[J]. The Journal of Physiology, 2010, 588(13): 2343-2350. DOI:10.1113/jphysiol.2010.191106.
[30] JANG H J, KOKRASHVILI Z, THEODORAKIS M J, et al. Gutexpressed gustducin and taste receptors regulate secretion of glucagonlike peptide-1[J]. Proceedings of the National Academy of Sciences, 2007, 104(38): 15069-15074. DOI:10.1073/pnas.0706890104.
[31] CORNISH J, MACGIBBON A, LIN J M, et al. Modulation of osteoclastogenesis by fatty acids[J]. Endocrinology, 2008, 149(11): 5688-5695. DOI:10.1210/en.2008-0111.
[32] HIRASAWA A, TSUMAYA K, AWAJI T, et al. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120[J]. Nature Medicine, 2005, 11(1): 90-94. DOI:10.1038/nm1168.
[33] CARTONI C, YASUMATSU K, OHKURI T, et al. Taste preference for fatty acids is mediated by GPR40 and GPR120[J]. The Journal of Neuroscience, 2010, 30(25): 8376-8382. DOI:10.1523/ JNEUROSCI.0496-10.2010.
[34] HARA T, HIRASAWA A, SUN Q, et al. Novel selective ligands for free fatty acid receptors GPR120 and GPR40[J]. Naunyn-Schmiedeberg’s Archives of Pharmacology, 2009, 380(3): 247-255. DOI:10.1007/s00210-009-0425-9.
[35] GILBERTSON T A, DAMAK S, MARGOLSKEE R F. The molecular physiology of taste transduction[J]. Current Opinion in Neurobiology, 2000, 10(4): 519-527. DOI:10.1016/S0959-4388(00)00118-5.
[36] TARUNO A, VINGTDEUX V, OHMOTO M, et al. CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes[J]. Nature, 2013, 495: 223-226. DOI:10.1038/ nature11906.
[37] HUANG Y A, MARUYAMA Y, ROPER S D. Norepinephrine is coreleased with serotonin in mouse taste buds[J]. The Journal of Neuroscience, 2008, 28(49): 13088-13093. DOI:10.1523/ JNEUROSCI.4187-08.2008.
[38] HUANG Y A, DANDO R, ROPER S D. Autocrine and paracrine roles for ATP and serotonin in mouse taste buds[J]. The Journal of Neuroscience, 2009, 29(44): 13909-13918. DOI:10.1523/ JNEUROSCI.2351-09.2009.
[39] ROPER S D. Parallel processing in mammalian taste buds?[J]. Physiology & Behavior, 2009, 97(5): 604-608. DOI:10.1016/ j.physbeh.2009.04.003.
[40] HUANG A L, CHEN X, HOON M A, et al. The cells and logic for mammalian sour taste detection[J]. Nature, 2006, 442: 934-938. DOI:10.1038/nature05084.
[41] FRANK M E, LUNDY R F, CONTRERAS R J. Cracking taste codes by tapping into sensory neuron impulse traffic[J]. Progress in Neurobiology, 2008, 86(3): 245-263. DOI:10.1016/ j.pneurobio.2008.09.003.
[42] BREZA J M, NIKONOV A A, CONTRERAS R J. Response latency to lingual taste stimulation distinguishes neuron types within the geniculate ganglion[J]. Journal of Neurophysiology, 2010, 103(4): 1771-1784. DOI:10.1152/jn.00785.2009.
[43] YOSHIDA R, MIYAUCHI A, YASUO T, et al. Discrimination of taste qualities among mouse fungiform taste bud cells[J]. The Journal of Physiology, 2009, 587(18): 4425-4439. DOI:10.1113/ jphysiol.2009.175075.
[44] KASHIMA H, HAMADA Y, HAYASHI N. Palatability of tastes is associated with facial circulatory responses[J]. Chemical Senses, 2014, 39(3): 243-248. DOI:10.1093/chemse/bjt074.
[45] BASSON M D, BARTOSHUK L M, DICHELLO S Z, et al. Association between 6-N-propylthiouracil (PROP) bitterness and colonic neoplasms[J]. Digestive Diseases and Sciences, 2005, 50(3): 483-489. DOI:10.1113/jphysiol.2009.175075.
[46] KAMBLE P. Review on: taste masking techniques[J]. Asian Journal of Pharmaceutical Sciences, 2013, 1(2): 1-16. DOI:10.1016/j.ajps.2013.07.015.
[47] DESHPANDE D A, WANG W C, MCILMOYLE E L, et al. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction[J]. Nature Medicine, 2010, 16(11): 1299-1304. DOI:10.1038/nm.2237.
[48] ZHANG C H, CHEN C, LIFSHITZ L M, et al. Activation of BK channels may not be required for bitter tastant-induced bronchodilation[J]. Nature Medicine, 2012, 18(5): 648-650. DOI:10.1038/nm.2734.
[49] MARGOLSKEE R F, DYER J, KOKRASHVILI Z, et al. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1[J]. Proceedings of the National Academy of Sciences, 2007, 104(38): 15075-15080. DOI:10.1073/pnas.0706678104.
[50] WU S V, ROZENGURT N, YANG M, et al. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells[J]. Proceedings of the National Academy of Sciences, 2002, 99(4): 2392-2397. DOI:10.1073/pnas.042617699.
[51] JANSSEN S, DEPOORTERE I. Nutrient sensing in the gut: new roads to therapeutics?[J]. Trends in Endocrinology & Metabolism, 2013, 24(2): 92-100. DOI:10.1016/j.tem.2012.11.006.
[52] MOSINGER B, REDDING K M, PARKER M R, et al. Genetic loss or pharmacological blockade of testes-expressed taste genes causes male sterility[J]. Proceedings of the National Academy of Sciences, 2013, 110(30): 12319-12324. DOI:10.1073/pnas.1302827110.
Progress in the Mechanism and Application of Guanosine-Binding Protein Coupled Receptor (GPCR) Family Taste
Receptor Signal Transduction
GAO Qian, ZHU Zhouhai, ZENG Wanli, HUANG Haitao, ZHOU Lan, LI Xuemei, YAO Jianhua* (Technology Center of China Tobacco Yunnan Industrial Co. Ltd., Kunming 650106, China)
Taste is the sensation produced when a substance in the mouth reacts chemically with taste receptor cells. When flavor substances stimulate taste receptors, the signal is transmitted to the brain via the nervous sensory system, resulting in the sense of taste after the analysis in the central nervous system. In recent years, taste receptors, taste signal transduction and application caused great concern of the public, and a lot of cell research results showed that different tastes are produced by different taste receptors. This paper reviews the signal transduction process and corresponding application of guanosine-binding protein coupled receptor (GPCR) family taste receptors.
taste receptor; signal transduction; application
10.7506/spkx1002- 6630-201603049
Q71
A
1002-6630(2016)03-0286-06
高茜, 朱洲海, 曾婉俐, 等. G蛋白偶联受体家族味觉受体的信号转导及应用研究进展[J]. 食品科学, 2016, 37(3): 286-291. DOI:10.7506/spkx1002-6630-201603049. http://www.spkx.net.cn
GAO Qian, ZHU Zhouhai, ZENG Wanli, et al. Progress in the mechanism and application of guanosine-binding protein coupled receptor (GPCR) family taste receptor signal transduction[J]. Food Science, 2016, 37(3): 286-291. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201603049. http://www.spkx.net.cn
2015-03-05
云南中烟工业有限责任公司项目(2013JC01)
高茜(1984—),女,助理研究员,博士,研究方向为生物化学与分子生物学。E-mail:gaoqian840905@163.com
*通信作者:夭建华(1968—),女,研究员,博士,研究方向为生物学。E-mail:jhyao_2007@126.com
猜你喜欢
美食(2022年4期)2022-04-16
学苑创造·A版(2021年10期)2021-10-30
今日农业(2020年24期)2020-12-15
科学(2020年4期)2020-11-26
幼儿画刊(2019年8期)2019-11-04
中国调味品(2019年2期)2019-03-18
旅游世界(2018年7期)2018-08-07
意林·少年版(2017年4期)2017-03-07
中西医结合心脑血管病杂志(2016年20期)2016-03-01
