
鸡mtDNA D-loop区单倍型簇频率与中国家鸡多母系起源研究
2017-01-12 04:52崔建勋吕雪梅张细权
畜牧兽医学报 2016年12期
梁 勇,康 乐,崔建勋,吕雪梅,杨 宁,张细权
(1.广东省家禽科学研究所,广州 510520; 2.华南农业大学动物科学学院,广州 510642; 3.中国农业大学动物科技学院,北京 100193; 4.广东省农业科学院农业经济与农村发展研究所,广州 510640; 5.中国科学院北京基因组研究所,北京 100101)
鸡mtDNA D-loop区单倍型簇频率与中国家鸡多母系起源研究
梁 勇1,2,3,康 乐2,4,崔建勋2,4,吕雪梅5,杨 宁3,张细权2*
(1.广东省家禽科学研究所,广州 510520; 2.华南农业大学动物科学学院,广州 510642; 3.中国农业大学动物科技学院,北京 100193; 4.广东省农业科学院农业经济与农村发展研究所,广州 510640; 5.中国科学院北京基因组研究所,北京 100101)
本研究旨在通过分析家鸡和原鸡D-loop区单倍型簇的区域性分布和频率揭示中国家鸡多母系起源的区域和传播途径。通过对1 347条鸡D-loop序列进行完全对比排序,共发现198种单倍型,聚成12个单倍型簇。分析结果显示,海岛型红色原鸡与家鸡和大陆型红色原鸡有明显差异;苏门答腊大陆型红色原鸡作为独立分支,也体现出与东南亚大陆型红色原鸡的区别。单倍型簇频率分析显示,Clade C代表的母系可能更早在东南亚及云南等地驯化并对我国南方家鸡形成产生影响,但并非直接由云南引入我国,应该另有途径;Clade D代表的母系可能更早在山东及河南一带驯化繁衍,后引入四川,并在四川开始大规模繁育饲养,此后向周边区域传播扩散,成为中国家鸡重要母系来源。本研究结果进一步支持了我国家鸡多母系起源的论断,并指出东南亚是我国南方家鸡重要的母系来源;同时,河南和山东一带也可能是我国家鸡起源的区域之一。
家鸡;mtDNA D-loop区;单倍型;聚类分析;频率;起源
以往的研究曾推测出家鸡起源于大陆型红色原鸡亚种(G.g.gallus),并认为家鸡单一起源于东南亚地区[1-2]。然而,中国考古发现却表明中国存在家鸡独立起源的社会物质文化基础[3-5]。H.Xiang等的研究认为,中国古代遗址出土的39个鸡骨残骸均为家鸡骨骼残骸,最古老的残骸距今约1.1万年[6],从而推测中国家鸡应该可能存在独立起源,但由于该研究缺乏动物形态学方面的证据,其研究结论在考古学领域可能会引起较大的争议。
采用分子生物学手段开展的中国家鸡起源研究显示,中国家鸡可能具有多母系起源。Y.Liu等对家鸡和红色原鸡D-loop序列进行了分析,发现了9个高度遗传分化的单倍型簇,并且各单倍型分布具有区域特异性[7]。包文斌等对家鸡和红色原鸡分析结果也表明,一些中国家鸡群体起源于泰国红色原鸡亚种(G.g.gallus),另一些中国家鸡群体起源于中国红色原鸡亚种(G.g.spadiceus),在一些中国地方鸡种还同时具有这2种红色原鸡的遗传贡献;认为中国家鸡只起源于泰国或单纯起源于中国的观点都是不全面的[8]。国内学者对我国地方鸡种线粒体DNA的遗传多样性分析,均得出了我国家鸡存在多母系来源的结论[9-15]。
尽管前期研究均得出了一致结论,但所涉区域大多为华南、西南部等地,所涉及的北方品种较少,因此得出结论无法完整的说明北方家鸡起源及传播路径;同时,所涉及的红色原鸡样品均缺乏中国广西和海南岛地区的样品,无法对云南、广西及海南岛的红色原鸡亚群之间的遗传关系及对我国家鸡起源的影响进行全面阐述。本研究在NCBI已有数据基础上增加了广西和海南红色原鸡及华中、华北和西北地区家鸡品种,旨在通过分析各单倍型簇的区域性分布和频率较全面揭示我国家鸡多母系起源的区域及其传播路径。
1 材料与方法
1.1 样品来源
红色原鸡亚种(G.g.jabouillei)的两个亚群共43个个体,分别采自广西(28)和海南(15)。本研究采集的16个地方鸡种样本包括辽宁大骨鸡(10)、山东寿光鸡(17)、河南固始鸡(10)、河南斗鸡(10)、江西白耳黄鸡(16)、余干乌骨鸡(22)、广东清远麻鸡(13)、杏花鸡(12)、阳山鸡(15)、海南省地方鸡(16)、广西灵山鸡(10)、云南武定鸡(17)、茶花鸡(15)、西藏藏鸡(12)、青海海东鸡(24)、陕西太白鸡(19)。藏鸡基因组DNA由中国农业大学动物科技学院提供,其余样本均由华南农业大学动物科学学院保存。
1.2 线粒体D-loop序列扩增及测序
1.2.1 引物设计与合成 利用DNASART软件,根据家鸡线粒体D-loop序列(序列号:AB009433)设计引物;其中,上游引物L16750与Y.Liu等[7]的相同,H476为下游引物。引物由上海博亚生物技术有限公司合成。预期PCR产物长度约为795 bp。
上游引物L16750:5′-AGGACTACGGCTTGAAAAGC-3′;下游引物H476: 5′-ATGTGCCTGACCGAGGAACCAG-3′。
1.2.2 PCR扩增和测序 PCR扩增体系总体积为50 μL:2.5×buffer (含Mg2+),dNTP各0.5 mmol· L-1,引物L16750、H476 各5 μmol· L-1,LATaq酶2 U,基因组DNA约50 ng(LATap酶及10×buffer (含Mg2+)购于大连宝生物技术有限公司,dNTP购于上海生物工程有限公司),ddH2O补足50 μL。PCR扩增程序:94 ℃预变性2 min;32个循环中,94 ℃变性50 s,60 ℃退火45 s,72 ℃延伸30 s;最后72 ℃充分延伸2 min。反应完毕后,用凝胶成像系统观察并记录结果。与设计长度吻合的PCR产物委托上海博亚生物技术有限公司提纯测序。
1.3 数据处理和分析
除自测序列外,其他序列均源自GenBank (http://www.ncbi.nlm.nih.gov),同源序列所属品种及编号见表1。利用DNASART软件对测序获得的家鸡及原鸡正反向序列进行拼接编辑,并参照测序图核实每一个变异位点。利用ClustalX1.81软件[16]对被测序列和GenBank下载序列进行完全比对排序。利用MEGA 6.0软件,依据Neighbour-Joining法[17]构建不同单倍型的分子系统树。
表1 GenBank同源序列情况表
Table 1 Homologous sequences from GenBank

地 区Region品种和序列编号BreedsandsequenceID广 东GuangdongHuxu(AF512215⁃20),QingyuanPatridge(AF512255⁃60,AY465964⁃67)广 西GuangxiLuke(AF512237⁃45),WangfengWugu(AF512283⁃91)贵 州GuizhouDwarfWugu(AF512117⁃27),GuizhouMountainWugu(AF512128⁃38),WumengWugu(AY46600⁃03)河 南HenanGushi(AF512139⁃50),HenanGame(AY465968⁃71)湖 北HubeiHubeiBlackSilky(AF512200⁃9),HubeiSilky(AF512189⁃99),HybridSilky(AF512234⁃36),YunxianWugu(AF512307⁃16)江 苏JiangsuJiangsuSilky(AF512221⁃33),Langshan(AY466004⁃07)江 西JiangxiTaiheSilky(AF512273⁃82,AY465988⁃91),YuganBlack(AY465992⁃95)山 东ShandongJiningBairi(DQ462524⁃33,DQ462540,DQ462542),LaiwuBlack(DQ462563⁃70),Langya(DQ462534⁃39,DQ462544⁃45,DQ462550,DQ462556),Luxi(AY588621⁃28,DQ462521⁃25,DQ462547⁃49,DQ462551⁃52),Souguang(AY465984⁃87,DQ462541,DQ462543,DQ462546,DQ462553⁃55,AY465984⁃87),Wenshang(DQ462557⁃62)四 川SichuanCaoke(AF512091⁃107),ChengduBlackSilky(AF512067⁃75),ChengduSilky(AF512060,AF512062⁃66),HeikangLayer(AF512210⁃14),SichuanMountainWugu(AF512261⁃72),Ya’anNon⁃wugu(AF512317⁃23),Ya’anWugu(AF512328⁃37)西 藏TibetTibet(AY465960⁃63,DQ648776)云 南YunnanChahua(AF512076⁃90,AY392377⁃407,AY465972⁃75),Chigulu(AY392172⁃204),Douji(AF512108⁃16),Jiangbian(AY392205⁃32),Lv’erwu(AY392233⁃61),Nixi(AF512246⁃54,AY392262⁃95),Shenggou(D06(1),J01(2)),Tengchongxue(AF512057⁃59),Wenshanshandi(AY392296⁃340),Wuding(AY392341⁃76),Xishuangbanna(AB098664⁃66,AY588629⁃35,AY728085),Yanjing(AF512324⁃27,AY465996⁃99,AY465996⁃99)浙 江ZhejiangBaiyiner(AF128320⁃24),Lingkun(AF128325⁃29),WhiteLeghorn(AF128330⁃34),Xianju(AF128315⁃19),ZhejiangWugu(AF128340⁃44)印 度IndiaDomestic(DQ525610,DQ525603⁃08,DQ835958⁃66,DQ531490⁃99,DQ531500⁃07,DQ525611⁃13,DQ525609,DQ835967⁃72,AY704701⁃19,AY644966⁃73,DQ531508⁃14,DQ531515⁃19,DQ525614⁃19,DQ531527⁃31,DQ531520⁃26,DQ835973⁃80)欧 洲EuropeBarredPlymouthRock(AB007719,D82920),Domestic(AY644974⁃5020,AY704720⁃24),NewHampshireRed(AY235570⁃71),WhiteLleghorn(AB007723,D82923,D82925,AP003317,X52392,NC_001323,AP003580)

表1(续)
2 结 果
2.1 线粒体D-loop区域序列完全对比排序
通过对1 347条序列进行完全对比排序,共检测到101个变异位点,主要分布在400 bp的范围内,共发现198种单倍型(图1),聚类成12个单倍型簇。苏门答腊(Sumatra)的红色原鸡大陆型亚种(G.g.gallus)与东南亚大陆亚种之间存在差异,成为一个独立分支(Clade Q)。海岛型红色原鸡(G.g.bankiva)与其他单倍型有显著差异,成为一个独立分支(Clade R)。藏鸡个体的单倍型P01和P02聚为一支(Clade P)。分别来自越南和印度尼西亚的家鸡个体与东南亚红色原鸡(G.g.gallus)聚为一支(Clade N)。单倍型I01和I02包含了海南红色原鸡亚群(G.g.jabouillee),成为一个独立分支(Clade I)。大部分的家鸡与东南亚红色原鸡序列集中于7个大单倍型簇,分别为Clade A(36)、Clade C(31)、Clade D(40)、Clade E(18)、Clade H(17)、Clade J(17)和Clade L(23)。
2.2 利用Neighbour-joining法构建分子系统树
前期研究显示,海岛型红色原鸡(G.g.bankiva)与家鸡和东南亚大陆型红色原鸡具有较远的遗传距离[1-2]。排序的结果也得出了一致的结论。本研究不再对Clade R所代表的海岛型红色原鸡(G.g.bankiva)数据进行分子系统发育分析。剔除了Clade R单倍型数据后,分子进化树显示,单倍型聚合成8个较大的单倍型簇,其中,苏门答腊岛的大陆型红色原鸡(Clade Q)所在的单倍型簇也体现出与其他单倍型簇较为明显的遗传差异(图2)。
2.3 各单倍型簇在不同地区鸡品种中的分布
本研究将一个单倍型簇视为单一的母系血缘,并统计其在不同区域家鸡和原鸡中的出现频率,将频率数据按区域的空间距离进行排列,不同的单倍型簇呈现出了不同的分布模式。
2.3.1 Clade A单倍型簇在不同品种和区域的分布特点 如图3所示,Clade A在广西原鸡中的出现频率最高(0.857),并在云南原鸡(0.400)和东南亚原鸡(0.323)中保持一定频率,在海南原鸡中也有发现(0.151)。然而该类单倍型在我国内陆家鸡中出现的频率较低,只在山东(0.026)、浙江(0.036)、云南(0.027)等地的家鸡群体中以低频率出现。该单倍型簇所代表的母系血缘并没有表现出以高频率区(广西)为中心向比邻区域传播扩散的趋势,说明该母系血缘对我国内陆家鸡形成没有产生显著影响,然而该母系血缘可能与印度尼西亚家鸡(0.529)、藏鸡(0.529)和印度家鸡(0.385)形成有关。

图3 单倍型簇A在不同地区的分布及频率Fig.3 Distribution and frequencies of Clade A in different regions
2.3.2 Clade C单倍型簇在不同品种和区域的分布特点 如图4所示,Clade C频率在浙江、江西、湖北及广东地区的家鸡群体中较高,分别为0.536、0.542、0.500、0.491,并呈现出该母系血缘以高频区为中心向周边区域扩散的趋势。同样的趋势也出现在了云南、东南亚及周边区域,说明该母系在上述地区均发生了群体扩张,暗示该母系起源与这些区域有关,并对当地家鸡形成产生影响。
2.3.3 Clade D单倍型簇在不同品种和区域的分布特点 单倍型簇Clade D主要分布于四川、云南、河南及山东等地区。7个单倍型中也在日本家鸡中被发现,显示日本家鸡受到该母系血缘影响。如图5所示,该类单倍型分布呈现出以四川(0.661)和青海(0.708)为中心向周边区域扩散的趋势。类似的趋势也出现在河南(0.523)和山东(0.442)为中心的区域,暗示该母系的起源与这些区域有着密切联系。该单倍型簇在广西(0.036)、海南(0.133)和云南(0.033)红色原鸡样品中的频率较低,而在东南亚原鸡中没有检测到。

图4 单倍型簇C在不同地区的分布及频率Fig.4 Distribution and frequencies of Clade C in different regions

图5 单倍型簇D在不同地区的分布及频率Fig.5 Distribution and frequencies of Clade D in different regions
2.3.4 Clade E单倍型簇在不同品种和区域的分布特点 Clade E包含了18种单倍型。其中,E01和E02主要分布在日本、韩国、印度尼西亚,我国的江西、广东、贵州、云南、四川等省及欧洲品种中也有少量分布。如图6所示,该单倍型簇所代表的母系血缘在东南亚红色原鸡(0.032)和海南红色原鸡(0.067)中以低频率存在,但在印度(0.577)和欧洲(0.903)的家鸡中以高频率被检测到,说明该母系与印度和欧洲家鸡形成有关,也对韩国(0.363)及周边地区家鸡形成产生影响。
2.3.5 Clade H单倍型簇在不同品种和区域的分布特点 如图7所示,Clade H包含了17条单倍型,主要分布在我国和日本。其中日本的频率较高,达0.317;我国的广东(0.316)、青海(0.292)、湖北(0.265)及江苏(0.222)的家鸡群体中的分布也以一定的频率出现。该类单倍型的频率展现出离散型分布模式,提示日本家鸡与我国家鸡具有一定的联系。
2.3.6 Clade J和Clade L单倍型簇在不同品种和区域的分布特点 单倍型簇Clade J和单倍型簇Clade L分别包含了17条和23条单倍型。除了少量分布在日本、广东、广西和青海等地区外,该2个单倍型簇均为云南地方家鸡及周边红色原鸡(云南红色原鸡和东南亚红色原鸡)所专有,体现了显著的区域特异性,并暗示云南家鸡形成与当地红色原鸡有着密切的关系,但这2个单倍型簇所代表的母系并没有展现出向我国内陆家鸡扩散传播的趋势。

图6 单倍型簇E在不同地区的分布及频率Fig.6 Distribution and frequencies of Clade E in different regions
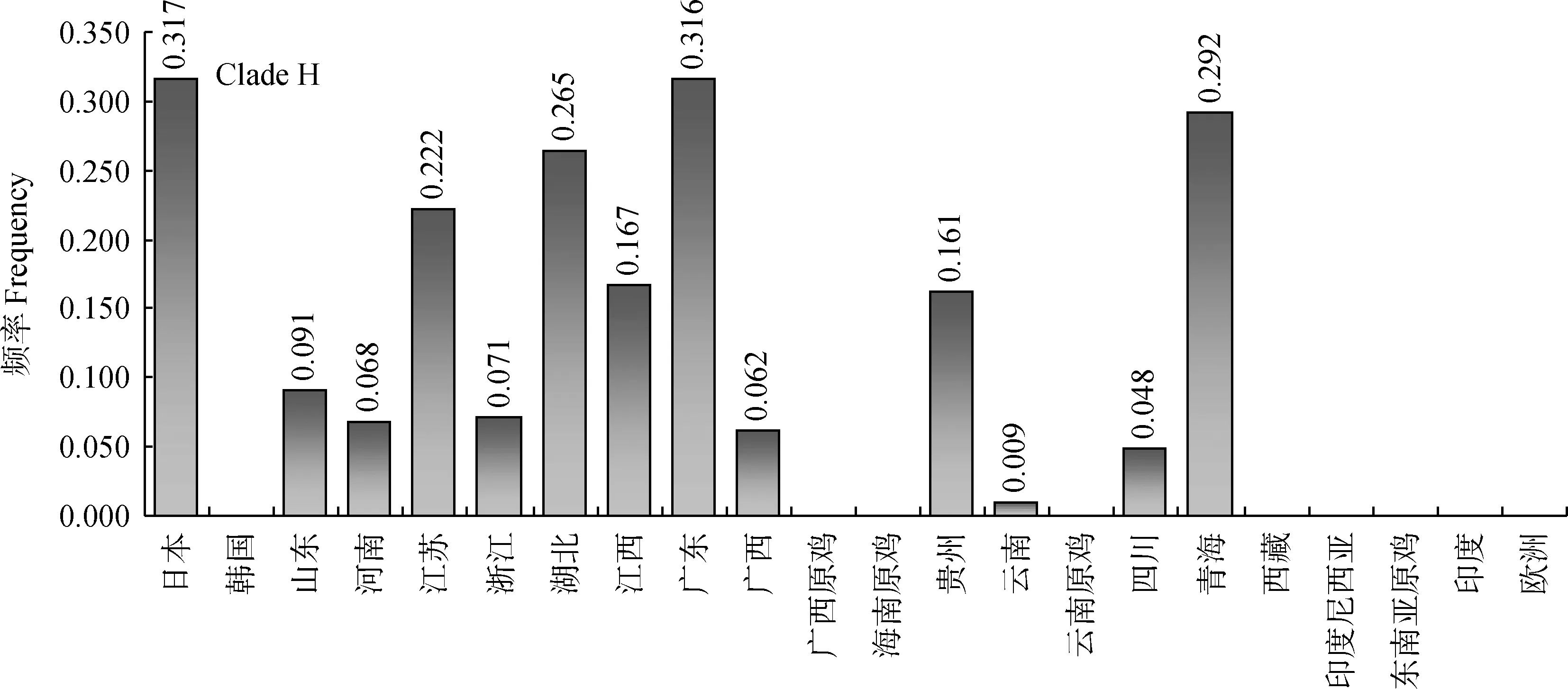
图7 单倍型簇H在不同地区的分布及频率Fig.7 Distribution and frequencies of Clade H in different regions
3 讨 论
3.1 红色原鸡不同群体间的遗传差异及分布规律
红原鸡(Gallusgallus),又名红色原鸡(Red jungle fowl),共有5个亚种[16-18]。Y.Liu等的研究显示,海岛型红色原鸡(G.g.bankiva)在D-loop区域单倍型变异上与大陆型红色原鸡有明显差异[7]。本研究的结果核实了Y.Liu等[7]的结论,但发现在苏门答腊岛的红色原鸡大陆型亚种(G.g.spdiceus)与东南亚大陆型红色原鸡各亚种之间亦存在明显差异(图1)。分子系统树也显示,苏门答腊岛的大陆型红色原鸡亚种(G.g.gallus)聚合成独立分支(Clade Q),显现出明显的区域特异性,这可能是地理生殖隔离所造成遗传分化。
中国国内有2个红色原鸡亚种,分别为滇南亚种(G.g.spdiceus)和海南亚种(G.g.jabouille)。

图8 单倍型簇J在不同地区的分布及频率Fig.8 Distribution and frequencies of Clade J in different regions

图9 单倍型簇L在不同地区的分布及频率Fig.9 Distribution and frequencies of Clade L in different regions
滇南亚种分布在与泰国交界的云南西部、西南部及南部的西双版纳。海南亚种终年留居云南东南部、广西西南、广东西南及海南岛。本研究检测到海南岛红色原鸡所特有的单倍型簇(Clade I),显示了其与广西红色原鸡及东南亚红色原鸡之间的遗传差异。在Clade A(0.151)、Clade D(0.133)、Clade E(0.067)中均检测到海南红色原鸡所具有的单倍型,但频率均较低。广西红色原鸡主要分布在Clade A(0.857)中,其中单倍型A10-12、A33等4个单倍型为广西红色原鸡所独有,同时云南红色原鸡和东南亚红色原鸡在该单倍型簇中均以较高频率被检测到,说明东南亚红色原鸡与广西红色原鸡具有共同的母性起源。云南红色原鸡主要分布于Clade A(0.400)和Clade C(0.433)中,在Clade D(0.033)、 Clade J(0.133)中频率较低,这一分布模式与东南亚红色原鸡的分布规律基本一致[18]。海南红色原鸡和广西红色原鸡尽管和地方家鸡间存在一定的基因交流,但并没显示对周边家鸡形成产生显著影响。
3.2 单倍型簇的地域性分布与我国家鸡形成的关系
刘坤凡等对中国红原鸡滇南亚种和海南亚种与我国茶花鸡、泰和鸡和寿光鸡等地方鸡种以及芦花鸡、洛岛红等外国鸡种进行了血型、蛋白质(酶)多态和DNA指纹分析,结果显示出“红原鸡-茶花鸡-泰和鸡、寿光鸡或芦花鸡-洛岛红”的进化阶梯[19]。同时,由于我国的4个鸡种系统聚为一个类群,而国外的原鸡聚为另3个类群,推测中国地方鸡种可能有独立的血缘来源[19]。贾晓旭等通过对藏鸡mtDNA的D-loop区全序列比对,发现藏鸡单倍型可以分为A、B、E 3个分支,推测A分支可能起源于红色原鸡的印度亚种分支,B和E与红色原鸡滇南亚种(G.g.spadiceus)聚为一类,推测这两个分支可能起源于云南、缅甸附近地区的红色原鸡滇南亚种[20]。本研究虽然不能完全核实贾晓旭等的结论,但可以认定的是藏鸡的地理地缘位置独特,应该有别于我国内陆家鸡的独特起源。本研究所涉及的家鸡和原鸡大都聚集到7个大的单倍型簇中,在此,本研究将依据各单倍型簇的频率分布规律,对不同母系起源及传播路径进行分析。
3.2.1 云南及与之相邻的东南亚地区与我国家鸡形成的关系 Clade A作为一个母系单位,其单倍型为日本斗鸡、印度尼西亚家鸡、广西红色原鸡和东南亚红色原鸡所共享。然而,Clade A所代表的母系血缘从频率分布上并没有向周边家鸡群体扩散的迹象。Clade L和Clade J尽管在云南家鸡或原鸡中也以一定频率出现,但也没有显示出向周边区域的扩散,这可能与地理生殖隔离有关。据此推断,中国家鸡的形成没有受到这一母系血缘的直接影响。同时,这也意味着东南亚作为家鸡重要的起源地,并不是通过与之接壤的云南对我国内陆家鸡产生影响的,可能另有通道。这在Clade C的区域性分布中可见端倪。Clade C的分布模式显示出以云南为中心向周边区域(四川和贵州)扩散的趋势。然而,Clade C的频率却在浙江、江西和广东家鸡群体中最高(湖南、陕西及海南地区家鸡个体数太少没有被考虑),并展现出以此为中心向周边地区逐渐降低的趋势。说明该血缘在上述地区发生了群体扩张。武艳平等对江西鸡系统发育的研究也显示整个江西家鸡群体曾出现了群体扩张事件[21]。据此推测,Clade C所代表的母系并非起源于中国,而是早期由东南亚引入中国浙江、江西及广东一带,然后进行大规模的扩群,并向周边地区传播。
3.2.2 关于我国家鸡独立起源的推测 据古文化遗址中所发现的家鸡骨骸,推测中国家鸡可能起源于7 500年以前[3-4]。其中,磁山家鸡跗骨长度稍大于现代原鸡,而明显小于现代家鸡,由此推断磁山家鸡可能是处于驯养阶段的早期品种[3-4]。山东大汶口遗址(距今6 500年)中出土的文字图案也揭示中国较早就开始了家鸡的驯化[5]。H.Xiang等2014年对中国北方4个古代遗址出土的39个鸡骨残骸进行了DNA分析,发现在距今约1万年的新石器时代,黄河流域中游地区的古代鸡中就出现了现代家鸡的几个主要单倍型,揭示出黄河流域的原始种群是现代家鸡的祖先之一,并推测中国北方地区曾经存在早期家鸡驯化事件[6]。本研究中,Clade D包含的单倍型主要分布于四川、云南、河南及山东等地区的家鸡群体中。其中,山东的鲁西斗鸡和河南斗鸡作为古老的品种出现在该单倍型簇中,暗示山东和河南地区与该母系家鸡最初的形成有关。然而,本研究表明中国家鸡独立驯化区域可能位于山东和河南一带,与H.Xiang等[6]所确定以中国河北省徐水县为中心的区域并不一致。考虑到以往关于我国古代家鸡的认定多有值得商榷之处,目前相对可靠的家鸡证据来自于距今3 300年左右的河南省安阳市殷墟遗址,这与本研究的结果更为接近。此外,Clade D在四川和青海(海东鸡)的家鸡群体中频率最高,分别为0.661和0.708,并展现出以此为中心向周边区域扩散的趋势,提示四川及周边地区可能为该母系家鸡重要的繁育中心。结合之前考古学的研究结果和Xiang H.等的结论,可推测Clade D所代表的母系可能更早在山东及河南一带驯化,之后才被引入四川。四川历史上曾经历了3次农业经济高度发展和人口快速膨胀的发展高峰期,外来人口的大量迁入带来了先进的农业技术,极大地推进了当地农业包括畜牧业的发展,这能够解释Clade D所代表的家鸡血缘为何在这一地区出现频率高峰。此后,随着四川经济对周边地区的影响不断扩大,家鸡品种也向周边区域传播扩散,成为中国家鸡重要的母系来源。
3.2.3 Clade H和Clade E的分布模式 Clade H所包含的17种单倍型广泛分布在我国各个省份,并呈现出离散型分布。该离散型分布模式既体现出区域内的群体扩张,但又存在区域间隔离。此外,该单倍型簇中以日本的单倍型种类居多,提示该血缘日本家鸡与我国家鸡间的遗传关系密切。从古至今,家禽品种的传播与人类社会的变迁密切相关,而Clade H所代表的家鸡血缘传播方式可能体现了人类社会一种特殊的生活和迁徙模式,这需要进一步的研究探讨。Clade E没有包含现有的我国及东南亚红色原鸡。印度和欧洲的家鸡群体中Clade E频率最高,并以低频率分布在日本、韩国和中国的一些省份,显示在印度和欧洲出现该母系家鸡的群体扩张,该母系血缘应为现代欧洲家鸡的母系来源。
4 结 论
本研究通过对鸡线粒体D-loop序列单倍型进行聚类和剖分,得出,我国的山东、河南一带可能是中国最先驯化和饲养家鸡的地区,并对四川家鸡的形成产生重要影响;云南及与之相连的东南亚地区作为家鸡起源较早区域,对中国南方家鸡的形成也具有深远的影响,但之间的遗传联系并非以陆地作为通路而建立的。本研究没有检测到广西和海南红色原鸡与我国地方鸡种显著的遗传相关。总而言之,不同地区、不同时期起源的家鸡群体,伴随着人类的社会活动,不断融合渗透,经历杂交选育的复杂历程,最终形成了现今具有不同地方特色的家鸡品种。
致 谢 感谢中国农业科学院北京畜牧兽医研究所韩建林研究员在本项目数据分析中给予的帮助;感谢青海大学农牧学院张玲勤教授、西宁市野生动物园王民园长、青海省海东市畜牧兽医站李有山同志对本研究的大力支持。
[1] FUMIHITO A, MIYAKE T, SUMI S, et al. One subspecies of the red junglefowl (Gallusgallusgallus) suffices as the matriarchic ancestor of all domestic breeds [J].ProcNatlAcadSciUSA, 1994, 91 (26): 12505-12509.
[2] FUMIHITO A, MIYAKE T, TAKADA M, et al. Monophyletic origin and unique dispersal patterns of domestic fowls [J].ProcNatlAcadSciUSA, 1996, 93(13): 6792-6795.
[3] CHOW B. The animal remains discovered at Baiying site, Tongyin, Henan Province [J].ChineseArchaeol, 1983, 4: 339-348.
[4] CHOW B. The animal remains discovered at Cishan village, Wu’an Hebei province [J].ActaArchaeologiaSinica, 1981, 62: 339-348.
[5] 马庆民,来永青.试谈莒地大汶口文化时期的社会经济[J].临沂师专学报,1999,21(5):74-75.
MA Q M, LAI Y Q. Trying to talking about the society economy of Dawenkou culture period [J].JournalofLinyiTeachers’College, 1999, 21 (5): 74-75. (in Chinese)
[6] XIANG H, GAO J, YU B, et al. Early Holocene chicken domestication in northern China[J].ProcNatlAcadSciUSA, 2014,111(49): 17564-17569.
[7] LIU Y, WU G S, YAO Y G, et al. Multiple maternal origins of chickens: Out of the Asian jungles [J].MolPhylogenetEvol, 2006, 38(1): 12-19.
[8] 包文斌,束婧婷,王存波,等. 中国家鸡和红色原鸡mtDNA控制区遗传多态性及系统进化分析[J].畜牧兽医学报,2008,39(11):1449-1459.
BAO W B, SHU J T, WANG C B, et al. Investigation on genetic diversity and systematic evolution in Chinese domestic fowls and red jungle fowls by analyzing the mtDNA control region [J].ActaVeterinariaetZootechnicaSinica, 2008, 39(11): 1449-1459. (in Chinese)
[9] 章学东,李庆海,张成先,等. 东乡绿壳蛋鸡线粒体DNA控制区多态性及与 9种地方乌鸡的进化分析[J]. 浙江大学学报(农业与生命科学版),2014,40(1):103-110.
ZHANG X D, LI Q H, ZHANG C X, et al. Mitochondrial DNA control region polymorphisms in Dongxiang blue-eggshell chicken and their evolutionary relatedness with nine domestic Wugu breeds [J].JournalofZhejiangUniversity(AgricultureandLifeSciences), 2014, 40 (1): 103-110. (in Chinese)
[10] 李 辉,吴 婵,熊光源,等. 黔东南小香鸡mtDNA D-Loop区遗传多态性分析[J]. 广东农业科学,2012,39(11):155-157.
LI H, WU C, XIONG G Y, et al. Analysis on genetic diversity of mitochondrial DNA D-loop region of Xiaoxiang chicken in Qiandongnan state [J].GuangdongAgriculturalSciences, 2012,39(11): 155-157. (in Chinese)
[11] 廖承红,王华伟,梁 浩,等. 文昌鸡线粒体DNA控制区序列遗传多样性分析[J].广东农业科学,2012,39(12):145-147.
LIAO C H, WANG H W, LIANG H, et al. Genetic diversity of mitochondrial DNA D-loop of Hainan Wenchang chicken [J].GuangdongAgriculturalSciences, 2012,39(12): 145-147. (in Chinese)
[12] 宋春红,陈红菊,马月辉,等. 中国6个地方鸡品种的母系起源[J]. 畜牧兽医学报,2007,38(7):735-740.
SONG C H, CHEN H J, MA Y H, et al. Maternal origins of six indigenous chicken breeds in China [J].ActaVeterinariaetZootechnicaSinica, 2007, 38 (7): 735-740. (in Chinese)
[13] 贾晓旭,唐修君,陆俊贤,等. 基于线粒体DNA D-loop区全序列分析大围山微型鸡遗传多样性及其起源进化关系的研究[J].南京农业大学学报,2015,38(4):656-660.
JIA X X, TANG X J, LU J X, et al. The investigation of genetic diversity and evolution of Daweishan mini chicken based on complete mitochondrial DNA D-loop region sequence[J].JournalofNanjingAgriculturalUniversity, 2015, 38 (4): 656-660. (in Chinese)
[14] 高玉时,贾晓旭,唐修君,等. 基于线粒体控制区(D-loop)全序列分析安义瓦灰鸡遗传多样性及其起源进化关系[J]. 农业生物技术学报,2015,23(7):940-944.
GAO Y S, JIA X X, TANG X J, et al. The genetic diversity and origin analysis of Anyi Tile-like chickens (Gallusgallusdomestiaus) based on mitochondrial DNA D-loop sequence[J].JournalofAgriculturalBiotechnology, 2015, 23(7): 940-944. (in Chinese)
[15] 解晓东,高景舜,王 欢,等. 西南地区三个地方鸡种母系遗传多样性评估[J]. 中国家禽,2015,37(11):19-22.
XIE X D, GAO J S, WANG H, et al. Study on matrilineal genetic diversity of three local chicken breeds in the southwest of China [J].ChinaPoultry, 2015, 37(11): 19-22. (in Chinese)
[16] HILLEL J, GROENEN M A, TIXIER-BOICHARD M, et al. Biodiversity of 52 chicken populations assessed by microsatellite typing of DNA pools [J].GenetSelEvol, 2003, 35(5):533-557.
[17] NEI M. Estimation of average heterozygosity and genetic distance from a small number of individuals [J].Genetics, 1978, 89(3): 583-590.
[18] PAUL A, JOHNSGARD P, JOHNSGARD J. Pheasants of the World: Biology and Natural History [M]. Edition 2. Washington: Smithsonian Books, 1999.
[19] 刘坤凡,程光潮. 中国红原鸡与家鸡血浆蛋白质(酶)多态性分析[J]. 畜牧兽医学报,1997,28(4): 295-299.
LIU K F, CHENG G C. Analysis of plasma protein polymorphism in red jungle fowl (Gallusgallus) and domestic fowl (Gallusdomesticus) of China[J].ActaVeterinariaetZootechnicaSinica, 1997, 28(4): 295-299. (in Chinese)
[20] 贾晓旭,唐修君,陆俊贤,等. 基于线粒体DNA D-loop区全序列分析藏鸡遗传多样性及其起源进化关系的研究[J]. 东北农业大学学报,2015,46(2):32-36.
JIA X X, TANG X J, LU J X, et al. Investigation of genetic diversity and evolution of Tibetan chicken based on complete mitochondrial DNA D-loop region sequence [J].JournalofNortheastAgriculturalUniversity, 2015, 46(2): 32-36. (in Chinese)
[21] 武艳平,霍俊宏,刘林秀,等. 江西地方鸡的系统进化及遗传多样性研究[J]. 江西农业大学学报,2011,33(6):1160-1163.
WU Y P, HUO J H, LIU L X, et al. Diversity and phylogenetic relationships of Jiangxi chicken breeds [J].ActaAgriculturaeUniversitatisJiangxiensis, 2011, 33(6): 1160-1163. (in Chinese)
(编辑 郭云雁)
The Frequencies of Chicken mtDNA D-loop Haplotype Clades and Multiple Maternal Origins of Chinese Domestic Chicken
LIANG Yong1, 2, 3, KANG Le2, 4, CUI Jian-xun2, 4, LÜ Xue-mei5, YANG Ning3, ZHANG Xi-quan2*
(1. Poultry Science Research Institute of Guangdong Province, Guangzhou 510520, China;2. College ofAnimalScienceofSouthChinaAgriculturalUniversity,Guangzhou510642,China;3.CollegeofAnimalScienceandTechnology,ChinaAgriculturalUniversity,Beijing100193,China; 4.InstituteofAgriculturalEconomicsandRuralDevelopment,GuangdongAcademyofAgriculturalSciences,Guangzhou510640,China;5.BeijingInstituteofGenomics,ChineseAcademyofSciences,Beijing100101,China)
The present study aimed to explain the domestication regions of Chinese chicken and its genetic propagation by analyzing the distribution and frequencies of D-loop haplotype clades of domestic chicken and wild red jungle fowls. The results showed that 198 haplotypes gained by sequencing 1 347 mtDNA D-loop fragments were clustered into 12 clades. Cluster analysis indicated that the island-type red jungle fowls (G.g.bankiva) were significantly different from domestic chicken and continental red jungle fowls (G.g.gallouse) of Southeast Asian, and that continental red jungle fowls (G.g.spdiceus) in Sumatra Island were independently branched from those of Southeast Asian. The distribution and frequencies analysis of haplotype clades indicated that the early domestic chicken with maternal blood presented by Clade C could origin from Southeast Asian and its surroundings(including Yunnan), but was not directly introduced into China via Yunnan. The maternal blood presented by Clade D could be domesticated in Shandong, Henan and its surroundings at early age, and then was introduced into Sichuan. From then on, the maternal blood began to reproduce on a large scale, propagated to surroundings, and became an important genetic source of Chinese domestic chicken. The present study can be an additional evidence to support the inference that Chinese domestic chicken have multiple maternal origins, and also indicate that Southeast Asian is an important original region of chickens in South China. In addition, Henan, Shandong and its surroundings are also important domestication regions of Chinese chicken.
chicken; mtDNA D-loop; haplotype; cluster analysis; frequency; origin
10.11843/j.issn.0366-6964.2016.12.003
2015-12-14
国家肉鸡产业技术体系分子育种岗位(CARS-42-G05)
梁 勇(1972-),男,广东阳江人,博士,主要从事动物遗传育种与繁殖研究,E-mail:lypromise@aliyun.com
*通信作者:张细权,博士生导师,教授,主要从事动物遗传育种与繁殖研究,E-mail:xqzhang@scau.edu.cn
S831.2
A
0366-6964(2016)12-2342-12
猜你喜欢
热带亚热带植物学报(2022年4期)2022-08-18
中国听力语言康复科学杂志(2021年6期)2021-12-21
昆虫学报(2021年11期)2021-12-08
——和田盘羊
野生动物学报(2021年4期)2021-11-14
食品与发酵工业(2019年18期)2019-10-11
疯狂英语·新阅版(2019年4期)2019-09-10
猪业科学(2018年5期)2018-07-17
英美文学研究论丛(2017年2期)2017-03-01
知识就是力量(2017年1期)2017-01-06
知识就是力量(2017年1期)2017-01-06
