
秦岭箭竹生物量分配规律
2017-01-18 07:27朱慧男何强利范晓东
绿色科技 2016年18期
朱慧男, 何强利, 范晓东
(陕西太白山国家级自然保护区管理局, 陕西 杨凌 712100)
秦岭箭竹生物量分配规律
朱慧男, 何强利, 范晓东
(陕西太白山国家级自然保护区管理局, 陕西 杨凌 712100)
为探明秦岭箭竹生物量分布规律,以太白山保护区厚畛子保护站管区后沟、云雾沟地区为研究区域,对秦岭箭竹进行了调查,探讨了秦岭箭竹生物量与海拔高度之间的关系,并分析了秦岭箭竹单株生物量在地上和地下的分配格局,结果表明:秦岭箭竹个体水平上在同一海拔高度不同个体间呈现数值变异。随着海拔的升高,同一海拔内个体差异降低。不同海拔秦岭箭竹生物量差异显著,随着海拔的升高,秦岭箭竹个体生物量降低;秦岭箭竹在个体水平,地上生物量、地下生物量和总生物量的变异系数均大于1,表现出较大的个体变异,地下生物量平均值为4.01小于地上生物量6.91。个体水平上的R/S值在0.2~1,8之间,平均值为0.57,大部分小于1,0.2~0.7之间分布最多;比较选出几个关系模型,建立的模型效果良好。
秦岭箭竹;生物量;规律
1 引言
生物量是指单位面积上所有生物有机体的干重。森林生物量是森林生态系统结构和功能的最基本特征之一,生态系统的能量和营养循环的研究首先依赖于生物量的数据,在研究森林生长和森林生态系统动态变化过程中,生物量的研究和测定显得极为重要,世界各国广泛开展了对森林生物量的监测[1,2]。国外就已开始了不同类型森林生物量的研究[3]。国内目前在个体、种群、群落、生态系统、区域、全国尺度、生物圈等多尺度上已开展了相应的生物量研究[4~10]。研究主要集中于油松[11~14]落叶松[15~18]、马尾松[19~20]、山杨、栎等树种的生物量分配以及模型构建方面,关于秦岭林区竹类的研究主要集中在主食竹种的资源调查、分类与分布、生物生态学特性等方面,对秦岭箭竹的单株生物量研究较少。
秦岭箭竹(Fargesiaqinlingensis)属禾本科竹亚科箭竹属,在太白山自然保护区内,秦岭箭竹主要分布于海拔1500~2900 m的针阔混交林林内,常为单一竹种在林冠下形成优势层片。对于高山地区的水土保持和环境防护具有重要的生态意义[21~26]。但由于人为打笋、采药等持续的干扰破坏及竹子开花、森林火灾等自然灾害的影响,天然秦岭箭竹林正面临着面积减少、生长衰退等问题的困扰。研究太白山保护区的秦岭箭竹单株生物量大小及其在地上和地下的分配格局,不仅对探索秦岭箭竹生物量与大熊猫栖息地选择关系具有重要意义,而且对评价秦岭大熊猫森林群落质量和对大熊猫栖息地森林群落进行科学管理和经营具有重要作用,对于进一步研究该区的碳储量和碳循环都有着非常重要的价值。
本研究以太白山保护区厚畛子保护站管区后沟、云雾沟地区为研究区域,对秦岭箭竹进行调查分析,探明秦岭箭竹生物量与海拔高度之间的关系,揭示秦岭箭竹分布规律,探讨秦岭箭竹单株生物量在地上和地下的分配格局,为确定该地区秦岭箭竹的生物量变化规律提供参考,进而为研究森林生产力和碳储量及碳汇功能的评价提供定量数据基础,为森林经营抚育管理提供一定的理论依据。
2 材料与方法
2.1 研究区概况
研究区域为太白山保护区厚畛子保护站后沟-云雾沟区域,地理位置为东经107°22 ′25″~107°51′30″和北纬33°49′30″~34°05′35 ″之间,海拔1598~2480 m;该区气候为暖温带大陆性气候,年均气温7.6 ℃,年均降水量751.8 mm,无霜期158 d,最高气温32.8 ℃,最低气温-25.5 ℃。最冷月为1月,平均气温为-5.0 ℃,最热月为7月,平均气温为19.1 ℃。2002~2007年间,区域内秦岭箭竹相继开花死亡,此后监测结果显示该区域内大熊猫分布急剧减少,区域内秦岭箭竹现处于自然更新阶段。各样地基本情况如下表:
2.2 研究方法
2.2.1 样地调查
2014年3月~2014年5月,采用样线调查和典型样地调查法,以太白山自然保护区管理局的巡护样线16A为调查样线,沿样线海拔每上升100~200 m设20 m×20 m的样地,调查内容包括GPS定位点、海拔、坡度、坡向、坡位、林冠层树种组成、植被类型、林冠郁闭度和竹子总体盖度。在每个样地内对角和中心位置布设2 m×2 m的小样方,调查竹子年龄、高度、基径、数量、密度、盖度等。
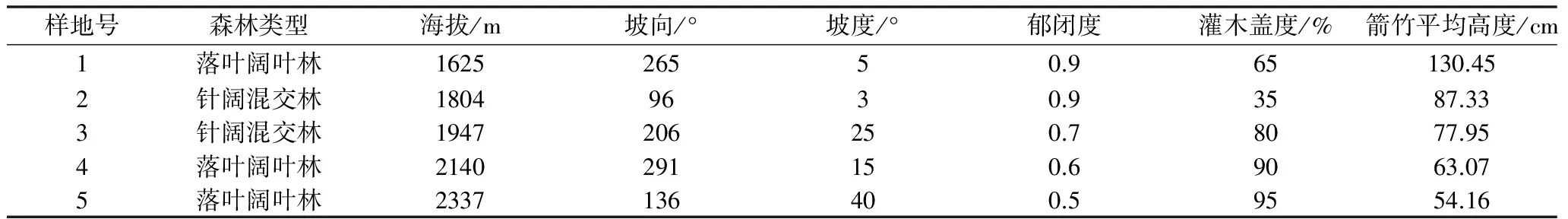
表1 各样地基本情况
2.2.2 生物量测定
竹子生物量测定采用收获法,在调查样地内随机选择10棵秦岭箭竹,按器官分别称取其鲜重并取200g鲜样,样品带回室内在105 ℃条件下烘干至恒重,计算各器官含水率,根据含水率计算生物量。
3 结果与分析
3.1 不同海拔单株生物量变化
从图1可以看出,在秦岭箭竹个体水平上,同一海拔高度不同个体间呈现数值变异。随着海拔的升高,同一海拔内个体差异降低。不同海拔秦岭箭竹生物量差异显著,随着海拔的升高,秦岭箭竹个体生物量降低。
由于低海拔地区光照、温度、土壤等环境条件优于高海拔地区,秦岭箭竹的生长较好,低海拔内秦岭箭竹个体生物量积累远高于高海拔地区。在同一海拔高度内,外界条件的小范围变化引起秦岭箭竹个体生长差异,低海拔地区秦岭箭竹生长对环境变化更敏感,而高海拔地区,环境条件较差,个体间差异不明显。
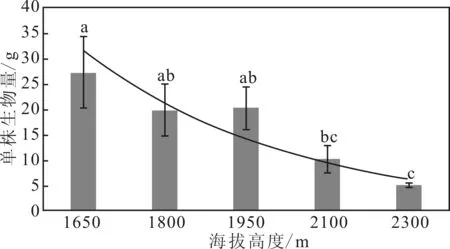
图1 不同海拔单株生物量
3.2 秦岭箭竹个体水平上生物量分配情况
由表2可以看出,秦岭箭竹在个体水平,地上生物量、地下生物量和总生物量的变异系数均大于1,表现出较大的个体变异,地下生物量平均值为4.01 g小于地上生物量6.91 g。
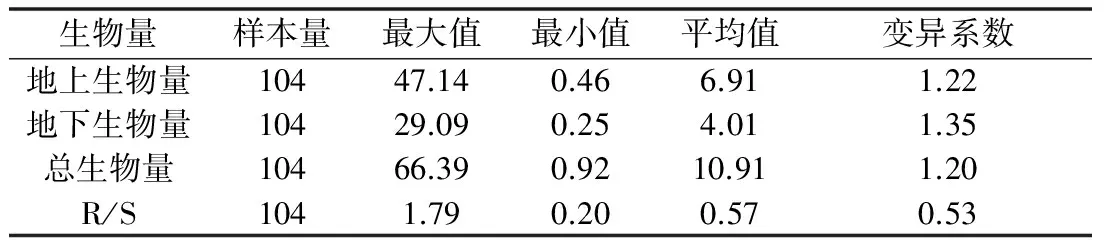
表2 秦岭箭竹个体水平上生物量分配
结合图2可知,个体水平上的R/S值在0.2~1.8之间,平均值为0.57,大部分小于1,0.2~0.7之间分布最多。
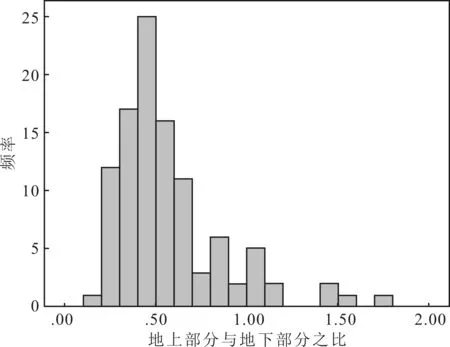
图2 个体水平上生物量地上与地下部分之比(R/S)频数分布
3.3 地上和地下生物量的相关生长关系
决定系数R2的大小决定了相关的密切程度。当R2越接近1时,表示相关的方程式参考价值越高;相反,越接近0时,表示参考价值越低。F值表示回归模型的方差与残差的比值(残差就是总方差减去回归模型的方差),理论上来说F值越大,通过检验的可能性就越大。可以想象F越大,表示残差越小,模拟的精度越高。P值是统计学上结果真实程度,p值越大,越不能认为样本中变量的关联是总体中各变量关联的可靠指标。在许多研究领域,0.05的p值通常被认为是可接受错误的边界水平。
对地上生物量与地下生物量关系进行常见的关系拟合,可以看出R2最大的是逻辑斯蒂模型,值为0.6992,最小的是指数函数模型,值为0.4328,剩余模型R2 均大于0.6,说明模型具有参考价值。P值均为0.0001,认为样本中变量的关联是总体中各变量关联的可靠指标。F值均较大,通过检验的可能性就较大,几个关系模型效果良好。在野外调查中,为了方便,可以根据实际情况,测定地上生物量或者地下生物量推算整体生物量,一定程度上节约了劳动时间和劳动量。

表3 地上和地下生物量的相关生长关系模型
注:X1地上生物量X2地下生物量
4 结语
由于水灾等自然因素,部分预设样地未发现秦岭箭竹分布,后重新布设5个样地,共调查样地5个。调查结果表明,秦岭箭竹个体水平上在同一海拔高度不同个体间呈现数值变异。随着海拔的升高,同一海拔内个体差异降低。不同海拔秦岭箭竹生物量差异显著,随着海拔的升高,秦岭箭竹个体生物量降低。由于低海拔地区光照、温度、土壤等环境条件优于高海拔地区,秦岭箭竹的生长较好,低海拔内秦岭箭竹个体生物量积累远高于高海拔地区。在同一海拔高度内,外界条件的小范围变化引起秦岭箭竹个体生长差异,低海拔地区秦岭箭竹生长对环境变化更敏感,秦岭箭竹个体生长差异很大,而高海拔地区,环境条件较差,所有个体间均生长不好,个体间差异不明显。秦岭箭竹是秦岭大熊猫食物的主要来源,其空间分布和数量多少在很大程度上决定了大熊猫的生境分布状况,在海拔1900~2600m秦岭箭竹生长最为茂盛的地段,为有效保护大熊猫及其生境,建议保护区在管护和森林群落经营管理方面,应尽量避免在秦岭箭竹适宜生长的地段海拔1900~2600m范围内林冠郁闭度0.6以下的落叶阔叶和针阔混交林中进行人为生产及其他干扰活动。
秦岭箭竹在个体水平,地上生物量、地下生物量和总生物量的变异系数均大于1,表现出较大的个体变异,地下生物量平均值为4.01g小于地上生物量6.91g。个体水平上的R/S值在0.2~1,8之间,平均值为0.57,大部分小于1,0.2~0.7之间分布最多。
通过数据分析,比较选出几个关系模型,建立的模型效果良好。在野外调查中,为了方便,可以根据实际情况,测定地上生物量或者地下生物量推算整体生物量,一定程度上节约了劳动时间和劳动量。
本文仅对太白山自然保护区后沟—云雾沟区域秦岭箭竹单株生物量进行了研究,不能代表太白山南坡秦岭箭竹生物量分布规律,应继续扩大研究区域,探明太白山南坡秦岭箭竹生物量以及其分布规律。
[1]罗 辑,杨 忠,杨清伟. 贡嘎山森林生物量和生产力的研究[J]. 植物生态学报,2000(2):191~196.
[2]刘志刚,马钦彦,潘向丽. 兴安落叶松天然林生物量及生产力的研究[J]. 植物生态学报,1994(4):328~337.
[3]Leith H, Whittaker R H. Primary Productivity of the Biosphere[M].New York: Springer Verlag. 1975.
[4]冯宗炜,陈楚莹,张家武,等. 湖南会同地区马尾松林生物量的测定[J]. 林业科学,1982(2):127~134.
[5]马钦彦. 中国油松生物量的研究[J]. 北京林业大学学报,1989(4):1~10.
[6]Gallis C. Probabilistic assessment of forest biomass storage times and its effect on cost: A beech biomass case study. Forest Products Journal, 2003 ,53 (10): 44~54.
[7]Fang J Y, Chen A P, Zhao S Q,et al. Estimating biomass carbon china 's forests: Supplementary notes on report published in science(291 : 2320-2322) by Fang et al[J]. Acta Phytoecologica Sinca, 2002, 26(2): 243 ~249.
[8]吴 刚,冯宗炜. 中国油松林群落特征及生物量的研究[J]. 生态学报,1994(4):415~422.
[9]方精云,刘国华,徐嵩龄. 我国森林植被的生物量和净生产量[J]. 生态学报,1996(5):497~508.
[10]刘玉萃,吴明作,郭宗民,等. 内乡宝天曼自然保护区锐齿栎林生物量和净生产力研究[J]. 生态学报,2001(9):1450~1456.
[11]秦仲焘,李志坚. 浅析天津市油松生物量测定方法[J]. 天津农林科技,2013(1):1~2.
[12]王 宁,王百田,王瑞君,等. 晋西山杨和油松生物量分配格局及异速生长模型研究[J]. 水土保持通报,2013(2):151~155+159.
[13]王 明. 森林油松生物量测定方法[J]. 农民致富之友,2013(13):142.
[14]张志明,常帅,张亚珍. 不同类型油松林生物量碳密度的研究[J]. 山西林业科技,2012(3):11~13+24.
[15]郭金堂,姜 鹏,郭凯,谷建才. 针阔混交林华北落叶松生物量模型及碳储量研究[J]. 西北林学院学报,2016(2):45~48.
[16]宋相禄. 不同营林措施对日本落叶松生物量的影响[J]. 林业科技,2003(2):4~6.
[17]闫涛,朱教君,杨凯,等. 辽东山区落叶松人工林地上生物量和养分元素分配格局[J]. 应用生态学报,2014(10):2772~2778.
[18]符利勇,唐守正,张会儒,等. 东北地区两个主要树种地上生物量通用方程构建[J]. 生态学报,2015(1):150~157.
[19]符利勇,曾伟生,唐守正. 利用混合模型分析地域对国内马尾松生物量的影响[J]. 生态学报,2011(19):5797~5808.
[20]樊后保,李燕燕,苏兵强,等. 马尾松-阔叶树混交异龄林生物量与生产力分配格局[J]. 生态学报,2006(8):2463~2473.
[21]王 瑞,周建云,曹 庆,等. 不同生境秦岭箭竹人工种群的生长特性研究[J]. 西北农林科技大学学报(自然科学版),2011(5):110~118.
[22]杜焰玲,张 莹,董国华,等. 秦岭山地秦岭箭竹群落物种多样性研究[J]. 西北农林科技大学学报(自然科学版),2013(10):59~66.
[23]吴高潮,李景侠,张国桢,等. 秦岭箭竹种群生物量研究[J]. 西北林学院学报,2014(1):46~50.
[24]马青青,刘建军,余 鸽,等. 佛坪国家级自然保护区秦岭箭竹克隆结构的SSR分析[J]. 生态学报,2016(20):1~10.
[25]王小红,周建云,张文辉,等. 大熊猫栖息地秦岭箭竹无性系构件形态研究[J]. 西北农业学报,2010(11):93~99.
[26]任 毅,刘明时,等.太白山自然保护区生物多样性研究与管理[M].北京:中国林业出版社,2006.
2016-08-10
秦岭大熊猫及其栖息地保护网络巡护员能力建设小额基金项目
朱慧男(1989—),男,助理工程师,主要从事森林保护与森林植物群落研究工作。
S795
A
1674-9944(2016)18-0016-03
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
军事文摘(2022年14期)2022-08-26
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
天天爱科学(2022年4期)2022-05-23
当代陕西(2022年5期)2022-04-19
散文诗(2021年22期)2022-01-12
安徽农学通报(2020年7期)2020-05-26
环球时报(2019-05-23)2019-05-23
当代陕西(2019年9期)2019-05-20
