
草地植物群落组成对鹅绒委陵菜无性系拓展能力及生物量配置的影响
2017-03-02 08:33刘金平宗人旭
草业科学 2017年1期
樊 星,蔡 捡,刘金平,伍 德,宗人旭
(西华师范大学 西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009)
草地植物群落组成对鹅绒委陵菜无性系拓展能力及生物量配置的影响
樊 星,蔡 捡,刘金平,伍 德,宗人旭
(西华师范大学 西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009)
以不同优势种的6个草地植物群落中鹅绒委陵菜(Potentillaanserina)为材料,通过测定基株、母株、无性系3个等级的构件数量性状及生物量结构,分析鹅绒委陵菜在不同群落中的空间拓展及生物量分配差异,探讨其应对草地群落组成差异的生长策略。结果表明,1)群落组成对母株根系有极显著影响(P<0.01),6个群落中母株根长和根数均显著下降(P<0.05),基株的子株数及子株根长也显著降低,但子株根数未受显著影响。2) 群落组成对无性系匍匐茎茎长、茎直径及间隔子长有显著影响(P<0.05),群落中鹅绒委陵菜通过降低茎直径、增大间隔子长,增加基株的空间拓展能力。3)群落组成对基株纵向、横向拓展参数有显著影响(P<0.05),群落中鹅绒委陵菜显著增加基株拓展面积和拓展空间,降低拓展熵和拓展效益,以游击型生长方式,觅养或逃离环境资源。4)群落组成显著降低了基株、母株、无性系生物量累积(P<0.05),群落中基株生物量优先向母株分配,母株生物量优先向叶分配,群落中无性系生物量分配比虽下降,但匍匐茎分配比呈增加趋势。5) 群落组成对基株生物量配置有显著影响(P<0.05),多数群落中显著增大了基株根茎比、叶茎比。综上所述,鹅绒委陵菜通过形态可塑性,在6个群落中均可存活和生长,其基株拓展能力和生物量配置除受群落盖度影响外,或许还受群落物种的生物学特点和无性系子株间竞争的影响,有待深入研究。
鹅绒委陵菜;群落组成;无性系;生长策略;群落竞争
鹅绒委陵菜(Potentillaanserina)属蔷薇科(Rosacrae)委陵菜属(Potentilla),又名曲尖委陵菜、人参果、延寿草、蕨麻[1],是典型的高寒地区多年生匍匐茎型莲座状草本植物[2-3],具有游击型(guerilla)克隆生长特点[4],呈粗网状紧贴地表的匍匐生长习性,使其常处于植物群落的底层。植物群落的种类组成、多度、密度、盖度等数量特征,外表形态或相貌特征及层片、垂直、水平、时间等结构特征,使鹅绒委陵菜生存空间基本资源异质化,必将导致其在不同群落中的存在度(presence)、恒有度(constancy)、确限度(fidelity)、优势度(dominance)和重要值(importance value)差异。近年来,对鹅绒委陵菜药用价值、食用价值[5]、饲用价值[6]及景观价值[7-8]进行了大量研究,尤其对克隆生长特性[9]、空间拓展性[10]、生物学特性[11]、无性系结构[2,12]、繁殖分配[13]、匍匐茎扩散模式[12]、克隆分株的风险分摊和存活调控[14]进行了较为深入研究。对光照[15]、土壤养分[10]、水分胁迫[16]等单一因子胁迫研究表明,鹅绒委陵菜具有极强的形态可塑性和生理整合能力,植物群落中个体间互惠、竞争关系,使处于群落底层的委陵菜面临尤为复杂的综合性胁迫。目前未见草地植物群落组成对委陵菜生长、生理及生殖过程的形态塑性及生理整合影响的报道。
克隆植物具有分株、克隆片段和基株3个层次的等级结构系统特征[17],基因型相同的母株与分株之间通过匍匐茎或根茎相连,形成了有机的时空分布整体[18-19],通过不断更新与增加构件,从垂直和水平方向拓展空间, 以游击型(guerilla)克隆生长方式,觅养或逃离环境资源,提高对资源的利用效率[20]。尤其在辐射强、年积温低、生长期短的高寒地区,只有能够及时、准确、系统地依据生境胁迫进行生长策略调整的物种,才能在自然选择中得以存活和延续。鹅绒委陵菜是高寒草地的主要伴生种,同时也是草地退化演替进程中的主要指示种,不同草地群落中其地位和意义有所不同,可能是无性系扩展能力分异所致,因而研究不同植物群落内鹅绒委陵菜无性系的拓展能力,将有助于明确该种群在不同草地植物中的繁殖策略。因而本研究以6个不同草地植物群落中的鹅绒委陵菜为研究对象,通过测定基株、母株、无性系3个等级的数量性状及生物量结构差异,分析鹅绒委陵菜在不同群落中的空间拓展及生物量分配的调节能力差异,研究母株、无性系、基株不同层次应对或适应生境胁迫的生长策略。旨在探讨鹅绒委陵菜在不同植物群落中拓展能力、形态塑性及繁殖策略的差异,为鹅绒委陵菜的人工栽培时生境设置及养护技术奠定基础,为野生植物资源的开发、利用及保护提供依据。
1 材料与方法
1.1 试验地概况
试验于阿坝州红原县邛溪镇草原科学院试验基地内进行,该基地为大陆性高原温带季风气候[21],地理位置为102°32′ E、32°46′ N、海拔3 497 m,年均温1.1 ℃,极端高温23.5 ℃,极端低温-33.8 ℃,年降水量738 mm,相对湿度71%,≥10 ℃·d年积温865 ℃·d,年辐射量20.93~29.30×106KJ·m-2。土壤为草甸土,0-20 cm土壤的有效氮、有效磷、有效钾含量分别为276.0、10.2、131.0 mg·kg-1,有机质含量5.87%,pH 5.91。
1.2 试验材料
于2015年8月,以传播于6个废弃种子试验地自然形成的草地植物群落中的当年生野生鹅绒委陵菜为材料,群落组成及基本情况见表1,群落组成比例最高的为建群种和优势种,均处在生殖生长阶段。群落密度为单位面积枝条数,优势种高度为自然高度,盖度为群落植物地上部分的投影面积占地面的比率。
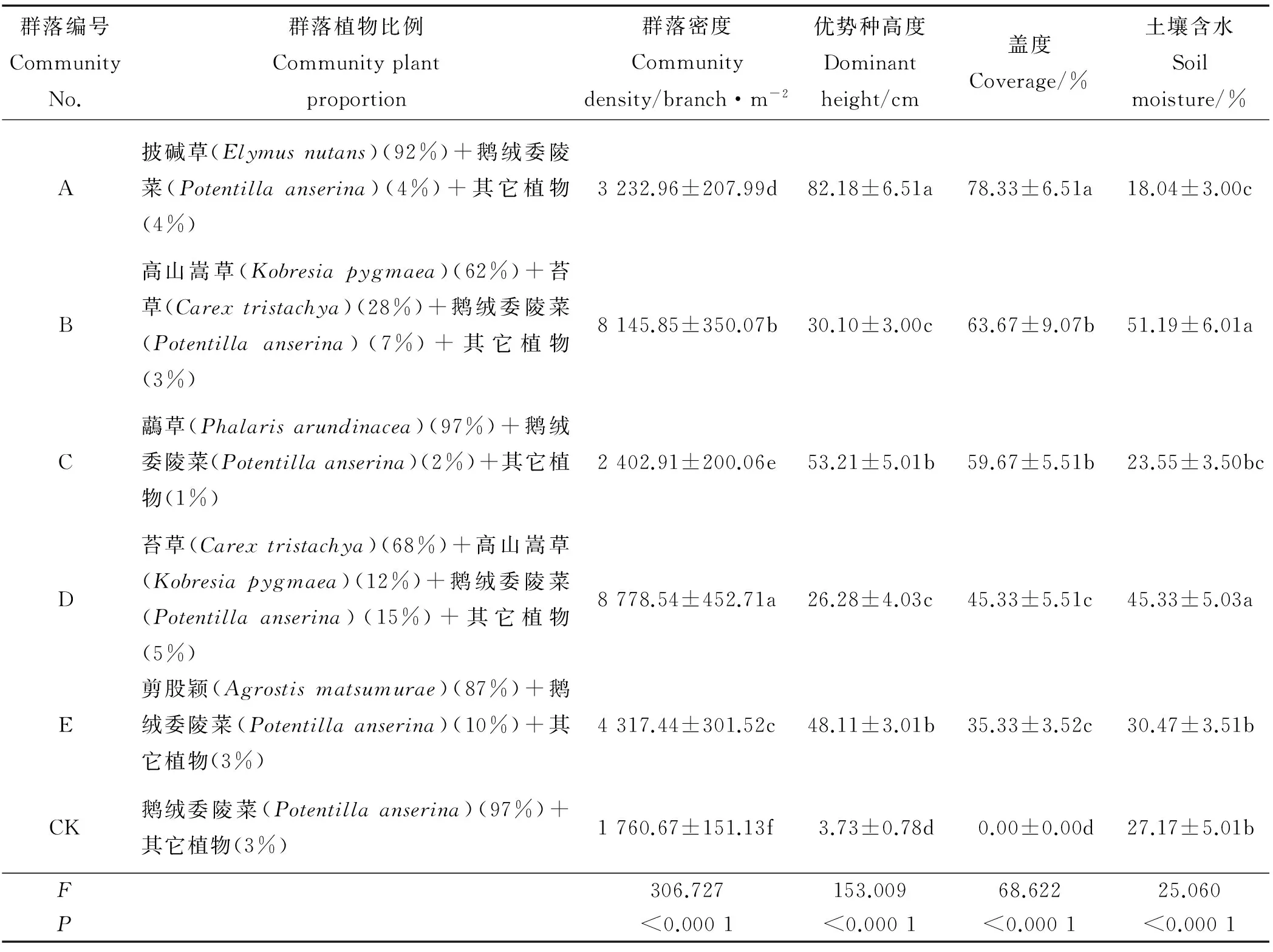
表1 群落组成及基本情况Table 1 Community composition and characterstics
注:同列不同小写字母间差异显著(P<0.05)。下同。
Note: Values with in the same column with different lower case letters are significant difference at 0.05 level. The same below.
1.3 试验设计
在每种群落内随机划定大小1 m×1 m样方,每样方按五点取样法设0.2 m×0.2 m的5个样点,测定每样点中鹅绒委陵菜的各个参数。设3次重复。
1.4 测定项目与方法
以基株为单位,8月中旬,当优势种处于盛花期,测定鹅绒委陵菜的下列项目:
株高:样点内随机选3基株,测定母株的自然高度。
无性系性状(匍匐茎和子株):样点内随机选3基株,测定基株的最大匍匐茎茎粗与茎长,茎数与总茎长,间隔子长,单茎子株数及子株总数。
根:挖取、清洗、拭干后,测定母株(或子株)的根长(80%根的长度),最长根长为最大根长,根系支数为根数。
生物量:将基株分为母株叶、子株叶、茎和根4部分,各自称鲜重后,装袋在105 ℃下烘至恒重,称干重为生物量。
生物量分配=某部分生物量/总生物量×100%;
地上生物量比率=地上生物量/总生物量×100%;
根茎比=地下生物量/地上生物量×100%;
茎叶比=茎生物量/叶生物量。
拓展指标[22]:纵向、横向拓展参数分别为母株自然高度、匍匐茎长度。
植株形态指数=横向拓展参数(匍匐茎长度)/纵向拓展参数(自然高度);
单株拓展面积=3.14×(横向拓展参数)2;
单株拓展空间=单株拓展面积×纵向拓展参数;
拓展熵=单株匍匐茎数量/单株拓展空间;
拓展效益=空间内生物量/单株拓展空间。
1.5 数据处理
用SPSS 19.0对鹅绒委陵菜各部分拓展能力,生物量和分配比进行多重比较和方差分析。
2 结果与分析
2.1 草地群落组成对根系拓展能力的影响
群落组成对母株的80%根长有显著影响(P<0.05)(表2),对最大根长及根数均有极显著的影响(P<0.01), CK根性状参数均显著高于其它群落(P<0.05),与其它植物共生竞争显著降低了鹅绒委陵菜母株的根拓展能力。5种群落组成间母株80%根长、最大根长均无差异(P>0.05),群落E的根数显著高于其它群落(P<0.05)。
群落组成对子株根数无显著影响(P>0.05),对子株80%根长和最大根长有极显著影响(P<0.01)。子株CK的80%根长显著大于其它群落(P<0.05),A群落次之,B、D、E群落间无显著差异,C显著小于CK和A群落。CK和A子株的最大根长显著高于B、C、D群落(P<0.05)。可见,在群落植物竞争压力下,鹅绒委陵菜母株与子株根系呈现出浅表化趋势,尤其B群落使母株、子株根拓展能力下降了60%左右。
2.2 草地群落组成对无性系拓展能力的影响
群落组成对匍匐茎茎直径有极显著影响(P<0.01)(表3),CK茎直径高于其它群落,但与C、D、E无显著差异(P>0.05),B茎直径最小仅为CK的1/2,A、C、D、E间无显著差异。群落组成对总茎长有显著影响(P<0.05),CK总茎长最大,B、D、E总茎长显著小于CK(P<0.05),仅为CK的40%左右。群落组成对间隔子长有极显著影响(P<0.01),C、D间隔子长显著高于CK和E(P<0.05),其它群落间无显著差异。群落组成对无性系单茎子株数有显著影响(P<0.05),仅B、E显著小于CK。群落组成对基株子株总数有极显著影响(P<0.01),共生植物影响使群落中鹅绒委陵菜基株子株总数均显著下降(P<0.05)。可见,群落组成对无性系拓展能力均具有显著影响,尤其对茎直径、间隔子长和基株子株总数影响极显著(P<0.01)。
2.3 草地群落组成对基株拓展指标的影响
群落组成对基株横向参数有显著影响(P<0.05)(表4),对纵向参数有极显著影响(P<0.01) ,C群落横向参数略高于其它群落,A群落显著低于C群落。A、B、D基株纵向参数显著高于E群落,A、B、C、D纵向参数显著高于CK近2.5倍。群落组成对植株形态指数无显著影响(P>0.05),但显著影响拓展面积(P<0.05),C的拓展面积显著高于A、E和CK。群落组成对基株拓展空间、拓展熵及拓展效益有极显著影响(P<0.01),C、D拓展空间显著高于A、E与CK。群落植物显著降低了鹅绒委陵菜基株的拓展熵及拓展效益,拓展熵是CK的14.28%~52.42%,拓展效益为CK的18.43%~35.16%。由F值可见,群落组成对基株拓展有显著影响,影响大小为拓展效益>纵向参数>拓展熵>拓展空间>横向参数>拓展面积>植株形态指数。
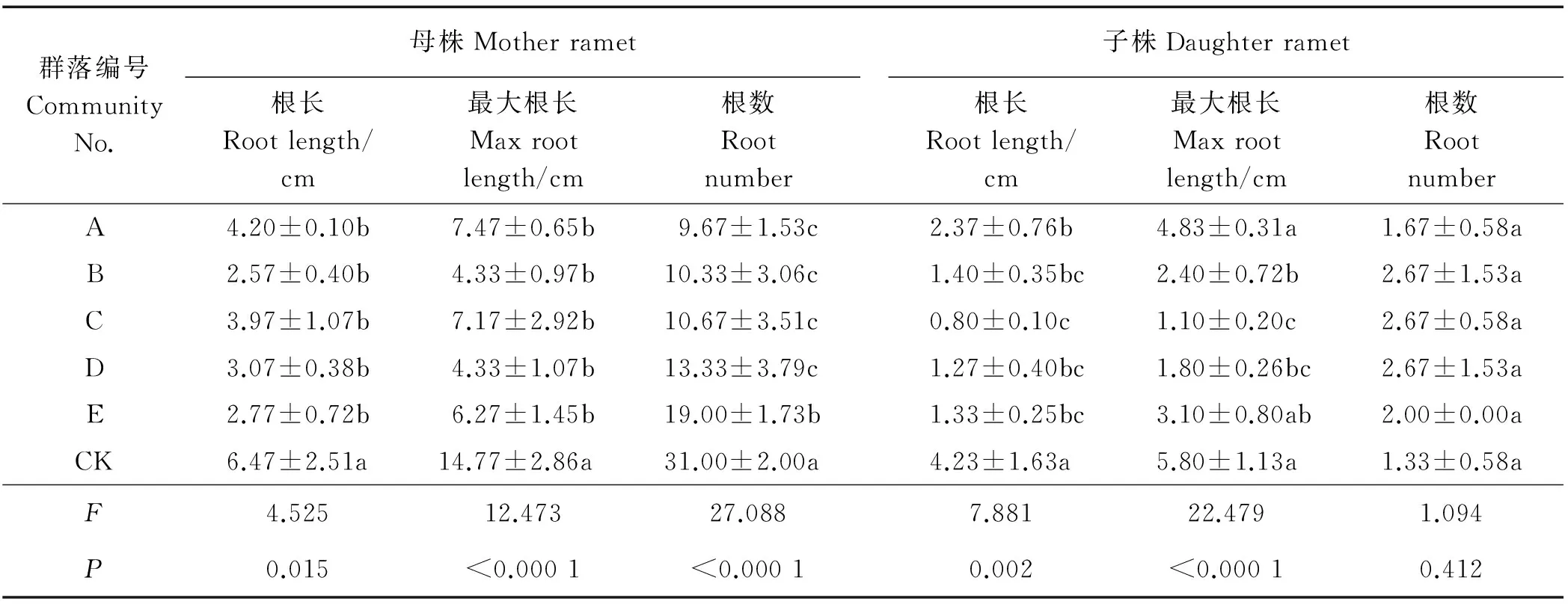
表2 不同草地植物群落对鹅绒委陵菜根系拓展能力的影响Table 2 Effects of different grassland plant communities on the root expansion capacity of P. anserina
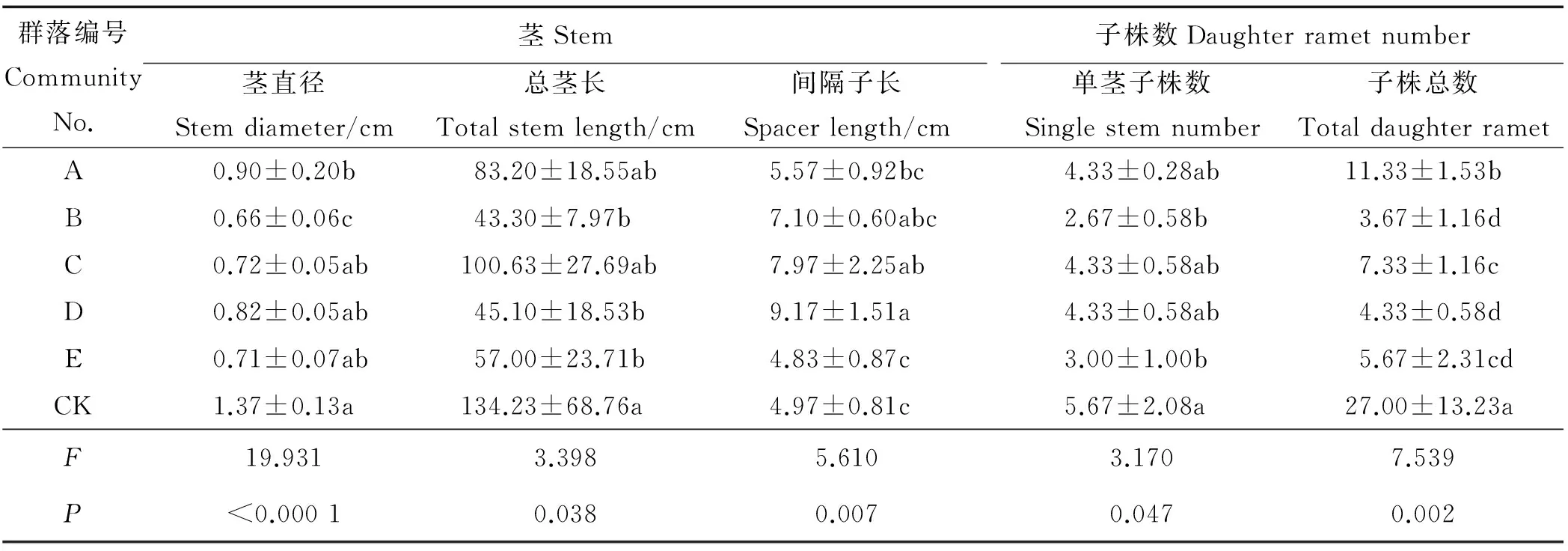
表3 不同草地植物群落对鹅绒委陵菜对无性系拓展能力的影响Table 3 Effects of different grassland plant communities on clone expansion capacity of P. anserina
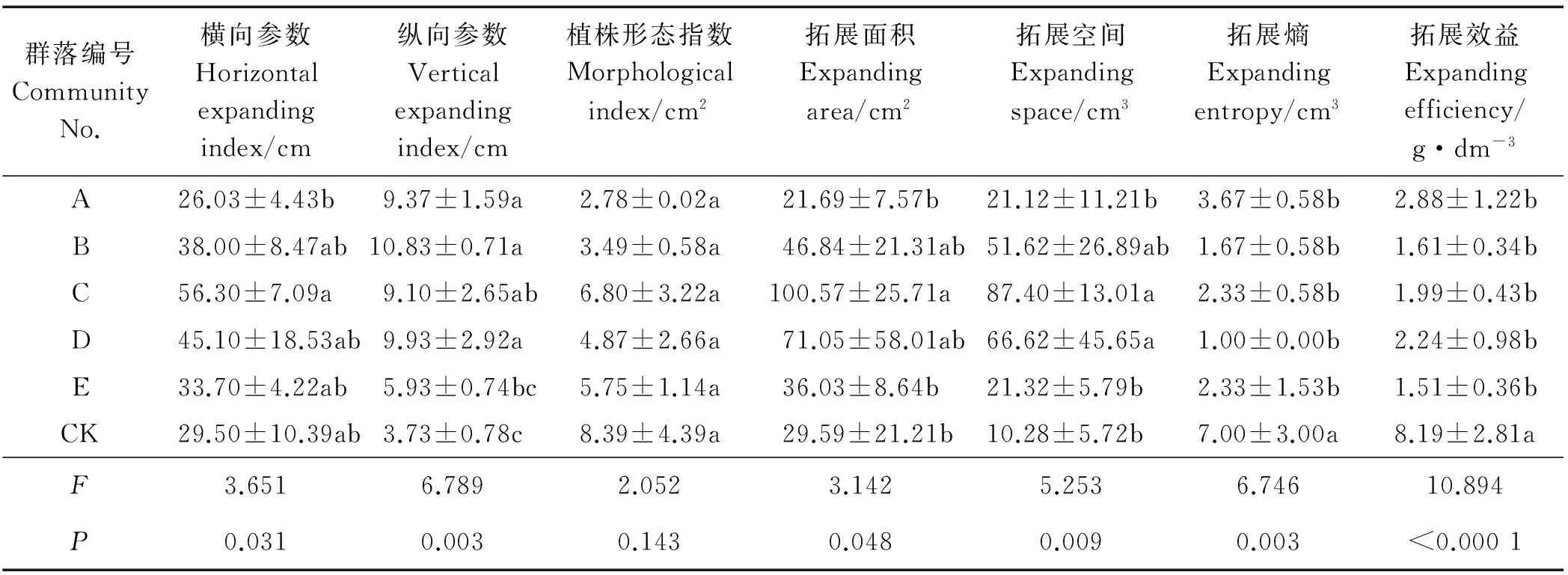
表4 不同草地植物群落对鹅绒委陵菜基株拓展指标的影响Table 4 Effects of different grassland plant communities on the expansion index of P. anserina genet
2.4 草地群落组成对生物量的影响
2.4.1 对生物量积累的影响 群落组成对子株叶生物量有显著影响(P<0.01)(表5),5个群落的子株叶生物量积累显著低于CK(P<0.05)。群落组成对无性系匍匐茎生物量无显著影响(P>0.05),对无性系总生物量有显著影响(P<0.05),CK无性系总生物量显著高于其它群落,群落间无差异。群落组成对母株叶生物量及总生物量积累有极显著影响(P<0.01),对根生物量有显著影响(P<0.05),5个群落母株叶、根和总生物量均显著低于CK(P<0.05),群落间均无差异。群落组成对基株生物量有极显著影响(P<0.01),5个群落基株生物量显著低于CK(P<0.05),群落间均无差异。由此可见,鹅绒委陵菜母株和无性系生物量均受植物群落组成影响而降低,对叶的影响大于茎和根。
2.4.2 对生物量分配的影响 群落组成对各构件分配比分配有显著影响(P<0.05) (表6),A和CK的无性系总分配显著大于其它群落,B和E母株总分配显著大于其它群落(P<0.05)。
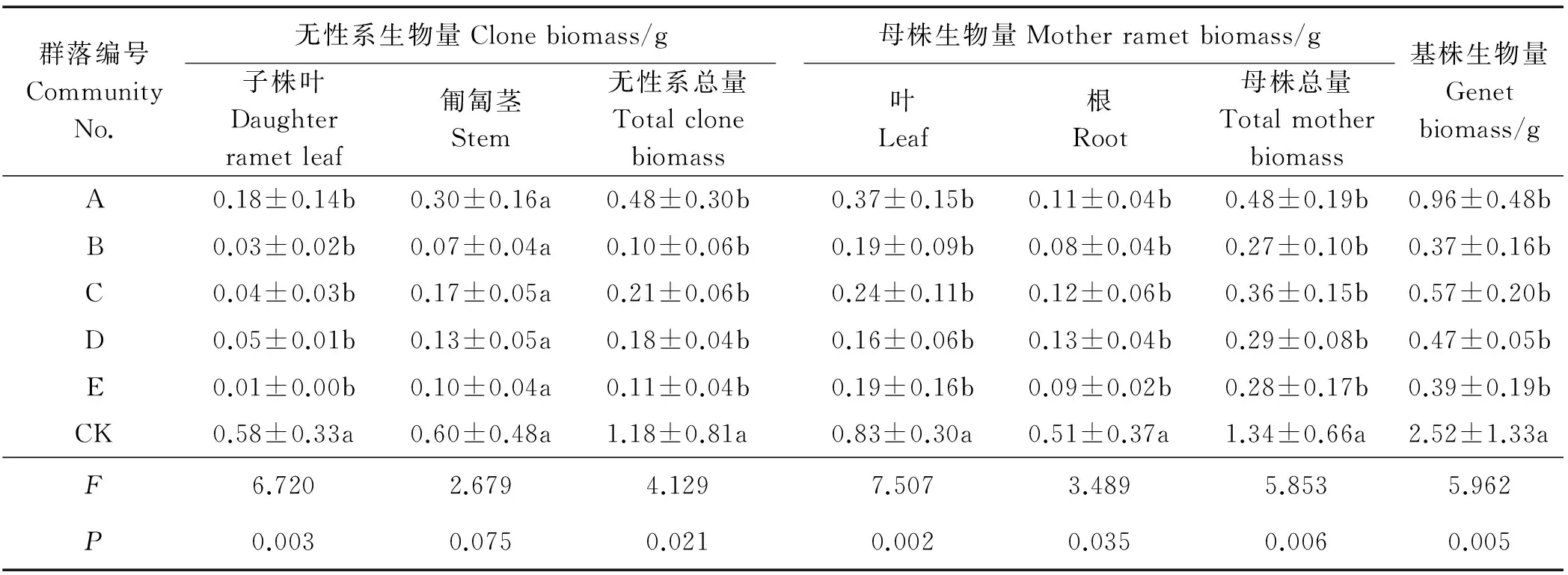
表5 不同草地植物群落对鹅绒委陵菜各构件生物量的影响Table 5 Effects of different grassland plant communities on various component biomass of P. anserina
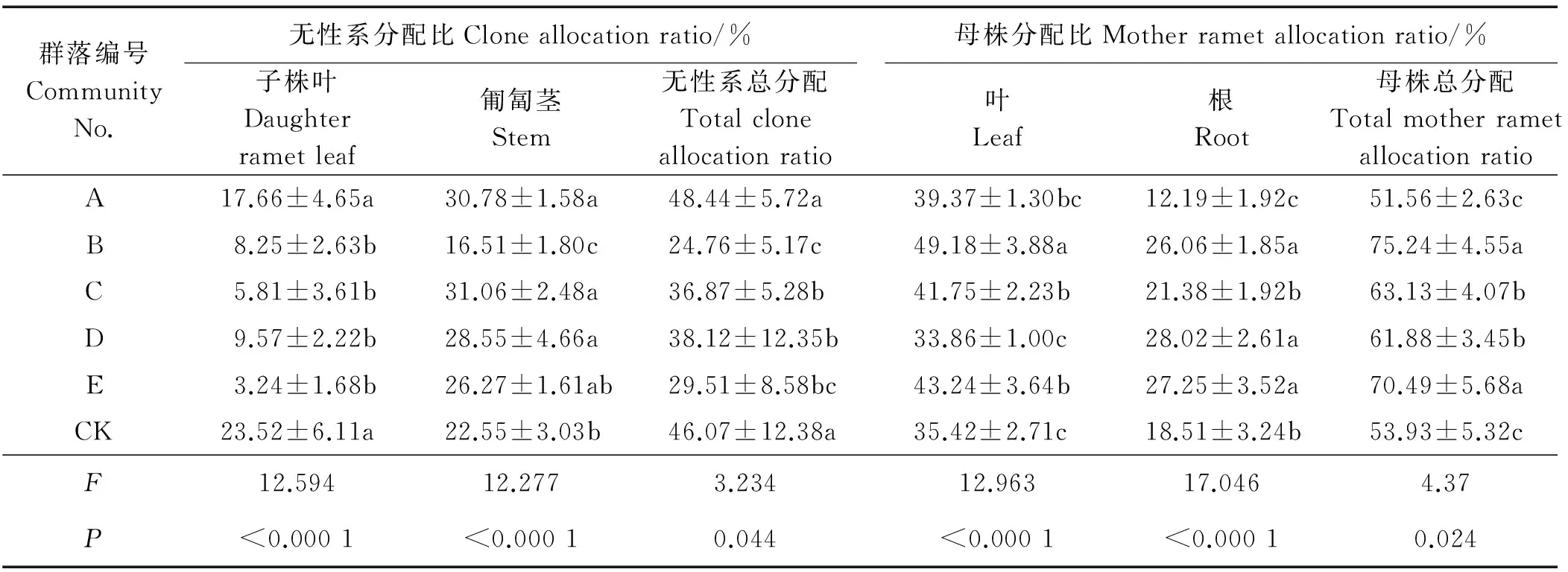
表6 不同草地植物群落对鹅绒委陵菜各构件分配比的影响Table 6 Effects of different grassland plant communities on various component allocation ratio of P. anserina
不同群落组成对无性系子株叶、匍匐茎分配均有极显著影响(P<0.01),CK和A子株叶分配显著高于其它群落(P<0.05),E子株叶分配仅为CK的1/8,而B、C、D与E间无显著差异。B群落匍匐茎分配比最低,A、C、D匍匐茎分配显著高于CK。群落组成对母株叶、根分配比均有极显著影响(P<0.01),B母株叶分配比最大,C和E次之, D和CK最低。B、D、E母株根分配比显著大于CK,A母株根分配比最低。可见,群落组成不仅影响鹅绒委陵菜母株、子株和无性系间生物量分配,对母株或子株的根、茎、叶等构件分配比也有显著影响。
2.4.3 对基株生物量结构的影响 群落组成对基株生物量结构有显著影响(P<0.05)(表7),A地上生物量分配比显著高于其它群落,D显著低于CK,其它群落间地上生物量比无差异。进一步分析表明,不同群落间根茎比存在极显著差异(P<0.01),A基株根茎比显著低于其它群落,B、C、D、E根茎比显著高于CK,共生植物使基株生物量结构发生了显著变化。对地上生物量进一步分析表明,群落组成对基株茎叶比有极显著影响(P<0.01),C、D、E基株茎叶比显著高于CK,A、B与CK无显著差异(P>0.05)。可见,群落组成不仅对基株地下、地上生物量结构有显著影响,对基株根茎比和茎叶比也有极显著影响。
3 讨论与结论
植物群落是生活在一定区域内通过互惠、竞争等相互作用,巧妙组合形成的植物集合,是群落植物适应共同生存环境的结果[22]。克隆植物具有强大的无性繁殖能力和空间拓展能力,使其在群落竞争中具有一定的优势[23],物种类型和种群密度是影响物种竞争能力的重要因素[24-25]。竞争物种的高度影响群落底层克隆植物的光照环境,光不仅为光合作用提供能量,同时也为植物光形态建成提供信号,克隆植物可通过增加间隔子长、根茎长和分枝强度逃离弱光环境[26],也可增强分株(枝)能力、提高空间拓展能力适应高光环境[27]。鹅绒委陵菜拓展能力易受邻体影响[28],在矮嵩草(Kobresiahumilis)内其无性系的间隔子长度、粗度、总长、干重都大于金露梅(Potentillafruticosa)灌丛内[29]。本研究分析了鹅绒委陵菜在不同草地群落中的拓展能力,表明草地群落组成对鹅绒委陵菜地下根系延伸、地上无性系拓展都有影响。植物群落中委陵菜母株的根数、根长均低于CK,说明种间竞争降低了母株根的拓展能力。不同群落植物组成使群落密度、高度、光照及土壤含水量均有差异,导致对母株根拓展力的影响不同,可见群落中植物生物学特点、生态学特性对委陵菜母株根系竞争力有显著影响。土壤理化性质及含水量对植物根系数量性状有显著影响[30],根系拓展深度、拓展范围及分布密度决定根系吸收水、肥的能力,对植株地上部分生长发育及抗逆性有重要作用。不同群落中委陵菜根系拓展能力差异,直接导致地上无性系的拓展能力不同。基株的适应性及生存能力由母株与无性系拓展能力决定。本研究结果表明,群落组成对鹅绒委陵菜植株形态影响极小,对基株株高、间隔子长、总茎长、茎粗及子株数有显著影响,对基株纵向拓展的影响大于横向。或许群落中优势种高度和叶片开展方向,决定底层委陵菜承受的郁闭度,对光源“觅养”是导致纵向拓展差异的主要原因,或许还受优势种茎秆密度与强度的“挟持”作用。
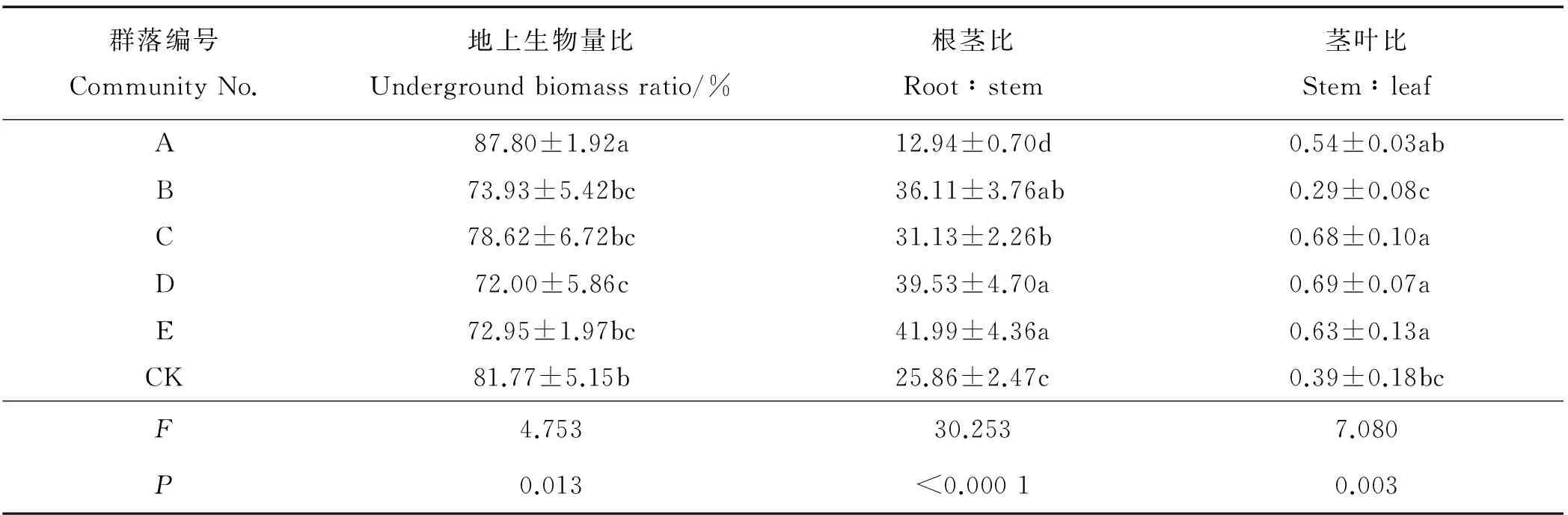
表7 不同草地植物群落对鹅绒委陵菜基株生物量结构的影响Table 7 Effects of different grassland plant communities on various components of the biomass structure of P. anserina
生物量是能量接受、同化、贮藏的基本形式,常用生物量累积量和生物量分配比,研究植物对异质生境的适应能力和植物各项功能间的权衡关系。局部特化的生物量分配格局是植物克服环境异质性的重要途径[31-32],通过适当改变生物量分配最大效率地利用资源,来提高其适合度、竞争力和存活几率[32-34],克隆植物生物量配置变化反映其对生境资源的生态适应对策[34]。群落盖度影响鹅绒委陵菜可利用的光照强度,鹅绒委陵菜母株与无性系生物量分配,不仅受群落物种对光资源竞争强度(盖度)的限制,还受其它资源竞争和物种生长习性的影响。如土壤含水量不仅影响营养生长阶段构件组成及拓展能力,对生殖投入及生殖效益也产生影响[35]。本研究中,群落物种极显著降低了基株生物量累积,增加母株分配而降低无性系分配,或许首先保证母株需求是委陵菜生存、生长、生殖的基本策略。母株生物量优先向叶分配,在所有群落中母株生物量分配大于或接近CK,确保不同盖度下,母株能固定、合成更多物质为无性系拓展或逃离环境提供能量,无性系生物量更多向可移动的匍匐茎分配,通过显著降低子株数、子株叶分配、匍匐茎粗,增加间隔子长度,尽可能提高匍匐茎生物量分配,使鹅绒委陵菜在地上生物量分配降低的情况下,增加了根茎比和茎叶比,体现了鹅绒委陵菜对群落组成的适应策略。研究结果与遮阴下喜旱莲子草(Alternantheraphiloxeroides)[36]、过路黄(Lysimachiachristinae)[37]的总生物量及根、茎、叶生物量结果一致,与遮阴环境下矮竹(Sasapalmata)的分株生物量明显下降[38],结缕草(Zoysiajaponica)总分株生物量、分株叶和根生物量随郁闭度增加显著减小[39]的研究结果相似。鹅绒委陵菜生物量分配除受群落物种的盖度影响外,或许还受邻体植物生物学特点的影响,关于邻体植物竞争对基株、母株、分株生物量分配的影响有待于进一步研究。
形态可塑性是植物适应环境、提高生态幅和耐受性的重要方式[40-43],是植物变异来源之一,对增加植物生存、竞争能力有很重要的作用[44],常用形态研究光照、水、肥、气、热等资源受限环境下植物生长策略。克隆植物是由母株和无性系构成,具有分株、克隆片段和基株3个层次的等级结构系统特征[17]。诸多研究表明,克隆植物具有极强的表型塑性和生理整合潜力[20]。克隆植物面对生境胁迫时,水分、光合产物、生理活性物质在分株、克隆片段和基株3个等级结构系统中的生理整合表现出明显的差异[20],致使其适应性、抗逆性及生存能力不同。鹅绒委陵菜基株、母株和分株的形态可塑性和生物量配置对生境资源有高度适应性[10],其游击型(guerilla)生长习性,分散分布分株提高了觅养或逃离环境资源的效率[20]。群落组成对鹅绒委陵菜拓展能力和生物量配置有显著影响,母株与无性系在不同群落中的营养器官和生物量的空间分布格局,是鹅绒委陵菜基株适应环境生长策略的外在表现。鹅绒委陵菜与群落物种竞争时,基株空间拓展能力与生物量累积受到了抑制,但通过形态实现了基株对所处群落的最适生长格局。本研究中群落组成对鹅绒委陵菜拓展能力和生物量分配有影响,在高密度群落中生物量优先向地上部分分配,尤其优先向母株叶和无性系匍匐茎分配,说明鹅绒委陵菜主要利用母株捕获能量,通过匍匐茎逃离胁迫环境。整合作用使克隆分株对生境产生局部反应和非局部反应,两种反应共同影响着克隆植物的生长和繁殖,进而影响基株和分株的适合度[45]。但很多研究表明,生理整合对克隆植物的形态特征修饰效果有正效应,也有负效应[46]。关于鹅绒委陵菜生理整合能力,以及生理整合对形态可塑性、构件等级结构、种群调节能力及适应能力的影响,有待于进一步深入研究。
References:
[1] 张彦芬.鹅绒委陵菜的生物学特性.西北农业学报,2011,20(12):126-129. Zhang Y F.Study on biological characteristics ofPotentillaanserine.Acta Agriculturae Boreali-Occidengtalis Sinica,2011,20(12):126-129.(in Chinese)
[2] Eriksson O.Mobility and space capture in the stoloniferous plantPotentillaanserine.Oikos,1986,46:82-87.
[3] 苟桂香.鹅绒委陵菜生长特征初步研究.草业与畜牧,2012(10):14-15,20. Gou G X.A preliminary study on the growth characteristics ofPotentillaanserina.Prataculture & Animal Husbandry,2012(10):14-15,20.(in Chinese)
[4] 周华坤,赵新全,周立,刘伟,韩发,古松.高寒草甸退化对鹅绒委陵菜克隆生长特征的影响.生态学报,2006,26(2):508-520. Zhou H K,Zhao X Q,Zhou L,Liu W,Han F,Gu S.Alpine meadow degradation alter the clonal growing characteristics ofPotentillaanserine.Acta Ecologica Sinica,2006,26(2):508-520.(in Chinese)
[5] 成英,宋九华,刘素君.鹅绒委陵菜多糖对荷瘤小鼠细胞因子的影响.安徽农业科学,2012,40(9):5177-5178,5180. Cheng Y,Song J H,Liu S J.Effects of polysaccharides fromPotentillaanserinaon the levels of cytokines in tumor mice.Journal of Anhui Agricultural Sciences,2012,40(9):5177-5178,5180.(in Chinese)
[6] 铁桂春,刘红献.鹅绒委陵菜的特性及应用价值.青海农牧业,2008(1):42-43. Tie G C,Liu H X.The characteristics and application value ofPotentillaanserina.Journal of Qinghai Agriculture and Animal Husbandry,2008(1):42-43.(in Chinese)
[7] 张庆良,陈秀红,张仁富,张凤霞,唐文煜.匍枝委陵菜的园林应用研究.山东林业科技,2006(2):25-26. Zhang Q L,Chen X H,Zhang R F,Zhang F X,Tang W Y.Study on landscape application ofPotentillaflagellaris.Shandong Forestry Science and Technology,2006(2):25-26.(in Chinese)
[8] 黄淑萍,杜桂娟,马凤江,杨姝,张哲.蛇莓委陵菜在园林绿化中的应用.园艺与种苗,2012(7):47-48,51. Huang S P,Du G J,Ma F J,Yang S,Zhang Z.Application ofPotentillaanserinain landscaping.Horticulture & Seed,2012(7):47-48,51.(in Chinese)
[9] 周华坤,周兴民,周立,沈振西,李英年.鹅绒委陵菜生长特征.西北植物学报,2002,22(1):9-17. Zhou H K,Zhou X M,Zhou L,Shen Z X,Li Y N.The clonal growing characteristic in the stoloniferous herb,Potentillaanserine.Acta Botanica Boreali-Occidentalia Sinica,2002,22(1):9-17.(in Chinese)
[10] 沈振西,陈佐忠,王彦荣,张镜锂,周华坤.高寒退化与未退化草甸土壤下匍匐茎鹅绒萎陵菜的克隆生长特征的比较.应用生态学报,2003,14(8):1332-1336. Shen Z X,Chen Z Z,Wang Y R,Zhang J L,Zhou H K.Clonal growth of stoloniferous herbPotentillaanserinaondegraded and non-degraded alpine meadow soil.Chinese Journal of Applied Ecology,2003,14(8):1332-1336.(in Chinese)
[11] 李军乔,王立祥.鹅绒委陵菜生物学特性的初步研究.西北农林科技大学学报:自然科学版,2003,31(3):190-192. Li J Q,Wang L X.Elementary study on the biological characteristic ofPotentillaanserinaL.Journal of Northwest A & F University:Natural Science Edition,2003,31(3):190-192.(in Chinese)
[12] Rautiainen P,Koivula K,Hyvarinen M.The effect of within-genet and between-genet competition on sexual reproduction and vegetative spread inPotentillaanserinassp.egedii.Journal of Ecology,2004,92(3):505-511.
[13] Saikkonen K,Koivnen S,Vuorisalo T,Mutikainen A P.Interactive effects of pollination and heavy metals on resource allocation inPotentillaanserinaL.Ecology,1998,79(5):1620-1629.
[14] Eriksson O.Ramet behavior and population growth in the clonal herbPotentillaanserine.Journal of Ecology,1988,76:522-536.
[15] Stuefer J F,Huber H.Differential effects of light quantity and spectral light quality on growth,morphology and development of two stoloniferousPotentillaspecies.Oecologia,1998,117:1-8.
[16] 盛海彦,李军乔,杨银柱,李红英.土壤水分对鹅绒委陵菜表型可塑性的影响.干旱地区农业研究,2004,22(3):119-122. Sheng H Y,Li J Q,Yang Y Z,Li H Y.Response of phenotypic plasticity ofPotentillaanserinaL. to soil moisture.Agricultural Research in the Arid Areas,2004,22(3):119-122.(in Chinese)
[17] 董鸣.克隆植物生态学.北京:科学出版社,2011. Dong M.Cloning of Plant Ecology.Beijing:Science Press,2011.(in Chinese)
[18] Stearns S C.The Evolution of Life Histories.Oxford:Oxford University Press,1992.
[19] Pigliucci M.Evolution of phenotypic plasticity:Where are we going now?Trends in Ecology & Evolution,2005,20:481-486.
[20] 张瑾,张明如,赵银宽.克隆植物表型可塑性研究若干进展.内蒙古农业大学学报,2008,29(4):271-275. Zhang J,Zhang M R,Zhao Y K.Advances in researches of phenotypic plasticity of the clonal plant.Journal of Inner Mongolia Agricultural University,2008,29(4):271-275.(in Chinese)
[21] 方精云,王襄平,沈泽昊,唐志尧,贺金生,于丹,江源,王志恒,郑成洋,朱江玲,郭兆迪.植物群落清查的主要内容、方法和技术规范.生物多样性,2009,17(6):533-548. Fang J Y,Wang X P,Shen Z H,Tang Z R,He J S,Yu D,Jiang Y,Wang Z H,Zhen C Y,Zhu J L,Guo Z D.Methods and protocols for plant community inventory.Biodiversity Science,2009,17(6):533-548.(in Chinese)
[22] 刘金平,张新全,游明鸿,陈永霞,王讯.扁穗牛鞭草克隆单株地上空间拓展能力与拓展效益多样性研究.中国生态农业学报,2007,15(6):12-17. Liu J P,Zhang X Q,You M H,Chen Y X,Wang X.Diversification of expansion capacity and efficiency of clonedHemarthriacompressa.Chinese Journal of Eco-Agriculture,2007,15(6):12-17.(in Chinese)
[23] 宋明华,董鸣.群落中克隆植物的重要性.生态学报,2002,22(11):1960-1967. Song M H,Dong M.Importance of clonal plants in community.Acta Ecologica Sinica,2002,22(11):1960-1967.(in Chinese)
[24] Keddy P,Nielsen K,Weiher E,Lawson R.Relative competitive performance of 63 species of terrestrial herbaceous plants.Journal of Vegetation Science,2002,13:5-16.
[25] Watkinson A R.Density dependence in single-species populations of plants.Journal of Theoretical Biology,1980,83:354-357.
[26] 冯图,黎云祥,杨子松,钱宝英.不同生境中淫羊藿克隆构型和分株种群特征.生态科学,2005,24(4):298-303. Feng T,Li Y X,Yang Z S,Qian B Y.Clonal architecture and ramet population characteristics ofEpimediumbrevicornumin different habitats.Ecologic Science,2005,24(4):298-303.(in Chinese)
[27] Yu F H,Dong M.Effect of light intensity and nutrient availability on clonal growth and clonal morphology of the stoloniferous herbHalerpestesruthenica.Acta Botanica Sinica,2003,45(4):408-416.
[28] Turkington R,Klein E.Integration among ramets ofTrifoliumrepens.Canadian Journal of Botany,1991,69:226-228.
[29] 周华坤,周立,刘伟,严作良,赵新全.两种生境下鹅绒委陵菜无性系形态的比较.西北植物学报,2002,22(6):1406-1414. Zhou H K,Zhou L,Liu W,Yan Z L,Zhao X Q.The comparison of morphological character ofPotentillanserinain two habitats.Acta Botanica Boreali-Occidentalia Sinica,2002,22(6):1406-1414.(in Chinese)
[30] 蔡捡,张小晶,李莹,曾晓琳,刘金平.基质对中华结缕草扦插成活率及幼苗生长特性的影响.草业科学,2015,32(7):1041-1046. Cai J,Zhang X J,Li Y,Zeng X L,Liu J P.Effect of soil substrate on the survival rate and component characteristics ofZoysiasinicacutting seedling.Pratacultural Science,2015,32(7):1041-1046.(in Chinese)
[31] Harper J L,Silvertown J,Franco M.Plant Life Histories:Ecology,Phylogeny and Evolution.Cambridge:Cambridge University Press,1997.
[32] Bazaz F A,Grace J.Plant Resource Allocation.New York:Academic Press,1997.
[33] 张大勇.植物生活史进化与繁殖生态学.北京:科学出版社,2004. Zhang D Y.Plant Life-History Evolution and Reproductive Ecology.Beijing:Science Press,2004.(in Chinese)
[34] Weiner J.Allocation,plasticity and allometry in plants.Perspectives Plant Ecology Evolution & Systematics,2004,6(4):207-215.
[35] 范志超,王彦荣.不同生境苦豆子种群的生产性能.草业科学,2016,33(3):459-470. Fan Z Q,Wang Y R.The production characteristics ofSophoraalopecuroidespopulations in different habitats.Pratacultural Science,2016,33(3):459-470.(in Chinese)
[36] 许凯扬,叶万辉,李国民,李静.入侵种喜旱莲子草对光照强度的表型可塑性反应.武汉植物究,2005,23(6):560-563. Xu K Y,Ye W H,Li G M,Li J.Phenotypic plasticity in response to light intensity in the invasive speciesAlternantheraphiloxeroides.Journal of Wuhan Botanical Research,2005,23(6):560-563.(in Chinese)
[37] 王琼,刘霞,王爱丽,杨志勇,苏智先,王燕.过路黄克隆生长对光照强度的反应.西华师范大学学报:自然科学版,2003,24(4):390-395. Wang Q,Liu X,Wang A L,Yang Z Y,Su Z X,Wang Y.Clonal growth ofLysimachiachristinaeinresponse to light intensity.Journal of China West Normal University:Natural Sciences,2003,24(4):390-395.(in Chinese)
[38] Saitoh T,Seiwa K,Nishiwaki A.Importance of physiological integration of dwarf bamboo to persistence in forest under storey:A field experiment.Journal of Ecology,2002,90:78-85.
[39] 王浩,李德志,程立丽,何莹莹,纪倩倩,刘薇,赖苏雯,陈慧娟.光资源强度变化对结缕草植株克隆生长特征的影响.生态科学,2014,33(4):664-671. Wang H,Li D Z,Cheng L L,He Y Y,Ji Q Q,Liu W,Lai S W,Chen H J.Effect of light intensity variation on the clonal growth characteristics ofZoysiajaponica.Ecological Science,2014,33(4):664-671.(in Chinese)
[40] Sultan S E.Phenotypic plasticity and plant adaptation.Acta Botanica Neerlandica,1995,44:363-383.
[41] Jonasson S.Implications of leaf longevity,leaf nutrient re-absorption and translocation for the resource economy of five evergreen plant species.Oikos,1989,56:121-131.
[42] Callaghan T V,Jonasson S,Brooker R W.Arctic clonal plants and global change.In:de Kroon H,van Groenendael J.(eds).The Ecology and Evolution of Clonal Plants.Leiden:Backbuys Published,1997:331-357.
[43] Law R,McLellan A,Mahdi A S.Spatio-temporal processes in a calcareous grassland.Plant Species Biology,1993(8):175-193.
[44] 董鸣.资源异质性环境中的植物克隆生长:觅食行为.植物学报,1996,38(10):828-835. Dong M.Clonal growth in plants in relation to resource heterogeneity:Foraging behavior.Acta Botanica Sinica,1996,38(10):828-835.(in Chinese)
[45] Evans J P.The effect of local resource availability and clonal integration on ramet functional morphology inHydrocotylebonariensis.Oecologia,1992,86:268-275.
[46] 叶上游,潘爽,王景波,王堃.克隆植物生理整合作用研究进展.草原与草坪,2008(5):63-69. Ye S Y,Pan S,Wang J B,Wang K.Research progress on physiological integration of clonal plant.Grassland and Turf,2008(5):63-69.(in Chinese)
(责任编辑 张瑾)
Effects of grassland plant communities on ramet expansion ability and biomass allocation ofPotentillaanserina
Fan Xing, Cai Jian, Liu Jin-ping, Wu de, Zong Ren-xu
(Key Laboratory of Southwest China Wildlife Resources Conservation, Ministry of Education;China West Normal University, Nanchong 637009, China)
In this study,Potentillaanserinaplants were grown in six types of plant communities characterized by different dominant species. Quantitative traits and biomass structure at three levels (genet, mother ramet, and clone) were measured. The objective of this study was to analyse the differences in space expansion and biomass distribution ofP.anserinagrown in the different plant communities, and to determine its growth strategies in response to the different communities. The results showed: 1) Community composition had a highly significant effect on the roots of the mother ramet (P<0.01). Compared to the control, root length and root number of the mother ramet in all other communities significantly decreased (P<0.05), and the number of roots and root length of the daughter ramet also showed a decline. However, the number of roots of the daughter ramet was not significantly different in any community. 2) Community composition had a significant effect on the length and diameter of clone stolons and spacer length (P<0.05).P.anserinagrown in different communities exhibited greater expansion ability of the genet by reducing stem diameter and increasing spacer length. 3) Community composition had a significant effect on both the horizontal and vertical expansion indices of the genet (P<0.05).P.anserinagrown in different communities significantly increased expansion area and space, and decreased expansion entropy, thus increasing efficiency by using the guerrilla growth pattern to obtain needed environmental resources. 4) Community composition significantly reduced biomass accumulation in the levels of ramets (P<0.05). Biomass of the genet preferentially allocated to the mother ramet, and biomass of the mother ramet was preferentially allocated to the leaves. Compared to the control, the biomass allocation ratio of clones declined, but the allocation ratio of stolons exhibited an increasing trend. 5) Community composition had a significant impact on biomass structure of the genet (P<0.05). Compared to the control, the ratio of root to stem and leaf to stem of the genet grown in most communities significantly increased. In summary,P.anserinacan survive and grow because it has characteristics of morphological plasticity. Expansion ability and biomass allocation ofP.anserinagenets perhaps change because of the biological characteristics of the community species and competition among daughter ramets of clones. Further research is needed.
Potentillaanserina; community composition; clone; the growth strategy; community competition
Liu Jin-ping E-mail:jpgg2000@163.com
10.11829/j.issn.1001-0629.2016-0161
2016-03-30 接受日期:2016-05-24
四川省科技支撑计划(2011NZ0064)
樊星(1992-),男,四川攀枝花人,在读硕士生,主要从事园林植物栽培养护技术研究。E-mail:aqz348985051@qq.com
刘金平(1972-),男,山西临县人,教授,博士,主要从事草坪建植与养护技术研究。E-mail:jpgg2000@163.com
S812;Q945.79
A
1001-0629(2017)1-0119-10*
樊星,蔡捡,刘金平,伍德,宗人旭.草地植物群落组成对鹅绒委陵菜无性系拓展能力及生物量配置的影响.草业科学,2017,34(1):119-128.
Fan X,Cai J,Liu J P,Wu D,Zong R X.Effects of grassland plant communities on ramet expansion ability and biomass allocation ofPotentillaanserina.Pratacultural Science,2017,34(1):119-128.
猜你喜欢
纺织服装周刊(2022年48期)2023-01-10
西藏农业科技(2019年2期)2019-07-10
吉林农业(2018年18期)2018-09-25
南都娱乐周刊(2018年16期)2018-08-29
江苏农业科学(2017年3期)2017-05-02
诗林(2016年5期)2016-10-25
果树实用技术与信息(2016年8期)2016-09-21
纺织导报(2015年11期)2016-01-05
江苏农业科学(2015年5期)2015-10-20
江苏农业科学(2015年8期)2015-09-10
