
统一进化论
2017-06-14 10:29迈克尔·斯金纳
飞碟探索 2017年6期
迈克尔·斯金纳
译/晨飞
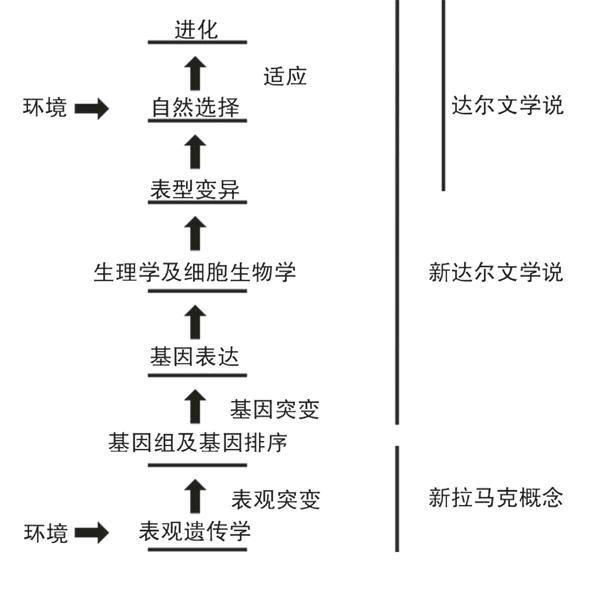
现代生物学的统一主题大体上建立在查尔斯·达尔文的进化论上,即自然选择的过程。自然会选择最健康、适应力最强的生物体,让它们繁殖、生存。这个过程也被称为适应,很可能有助于个体生存的性状被当作自适性状。随着生物体变化,新变体茁壮成长,物种出现、进化。19世纪50年代,达尔文描述了自然选择的这种机制,当时人们还不了解潜在的分子机制。但是,在过去的一个世纪,遗传学和分子生物学的发展勾画出一个现代版的新达尔文进化学说:基因序列任意突变,具有特定序列的生物体最能适应环境,继而繁殖并胜出。它们在环境发生变化、进化机

制再次生发之前最适应自己的那块地盘。
可是,对于进化的这种解释原来并不完整,其他分子机制在物种进化中也扮演着一定的角色。达尔文的学说存在一个问题,即:种族的确进化出更具适应能力的性状(生物学家称其为表型),基因序列任意突变的速度太慢,无法解释已观察到的许多变化。科学家很清楚这个问题,提出各种不同的遗传机制作为补充,这里只举两例:遗传漂变,即小群体的个体发生剧烈变化;或异位显性,即一组基因抑制另一组基因。
然而,即使有这样的机制在起作用,像人类这样的复杂有机体的基因突变率也比一些性状——从调节新陈代谢到抵抗疾病——的变化频率要慢很多,仅仅通过经典遗传学和新达尔文学说仍旧难以解释性状多样化的迅速出现。引用著名进化生物学家乔纳森.B.L·巴德意译T.S·艾略特的诗句来说,就是“在表型和基因型之间落下阴影”。
达尔文学说的问题超出了进化科学,进入生物学的其他领域和生物医学。例如,如果基因遗传决定我们的特性,那么具有相同基因的双胞胎为什么会得不同的疾病?为什么患多种特殊疾病的人只有少数(经常不到1%)具有共同的基因突变?如果突变率随意、稳定,那么为什么许多疾病在一二十年间频率就增长了10倍?为什么几百种环境污染物能够改变疾病攻击,而不能改变基因序列?在进化论和生物医学里,表型性状离异率比遗传变化和突变快得多,又是为什么呢?
部分解释可以从让-巴普蒂斯特·拉马克的一些概念里找到。拉马克提出这些概念之后50年,达尔文才发表了他的著作。拉马克的学说早已被扔到了科学的垃圾箱里。拉马克认为,“环境能够直接改变性状,然后遗传给未来若干代”等。拉马克当年是位于巴黎的国家自然历史博物馆无脊椎动物学教授,18世纪晚期至19世纪早期研究包括昆虫和蠕虫在内的生物体。他把“生物学”和“无脊椎动物”这些名词引入科学词汇,撰写了关于生物学、无脊椎动物和进化论等的著作。尽管有这样重要的学术生涯,拉马克却以他亵渎性的进化观念跟许多同时代的人以及历史上200多年的科学家相对抗。
最先,人们可能将他视为一名异教徒(但是在现代这是科学正统),尤其是当时达尔文不可批评的进化论,使得他的名字成了人们的笑柄。然而,在结束自己的职业时,达尔文终于醒悟:即使没有分子生物学的发现,他仍然能够看到随意变化没有迅速到足以充分支持他的学说。\
问题在这里:如果不只是自然选择在对基因突变起作用,那么是什么分子力产生自然选择所需的那一整套不同性状,以便完成这项工作呢?在达尔文提出他的学说将近一个世纪之后的1953年,詹姆斯·沃森和弗朗西斯·克里克正在试图揭开DNA和双螺旋之谜,这时出现了一条线索。那一年,爱丁堡大学的发育生物学家康拉德·瓦丁顿报告,果蝇在胚胎发育期间受到外界化学刺
激或温度变化时可能促使它发育出不同的翼结构,并且被它的后代一路遗传下去。瓦丁顿杜撰了一个现代术语“表观遗传学”来描述这种迅速变化的现象。显然,在沃森和克里克还没有揭示出DNA结构的时候,瓦丁顿已经认识到他的发现对进化论将产生潜在的影响:果蝇翼的一代变化支持异教徒拉马克的独到见解。看来环境能够直接影响性状。
生物学的规则绝不会是一个“仅遗传过程”,也不是一个“僅表观遗传的过程”。两者要完全结合。
瓦丁顿虽然描述了表观遗传的一般作用,但是他对分子元素或机制的了解并不比拉马克和达尔文多。但是,分子生物学越是解码生命机制,瓦丁顿的概念——还有拉马克的概念——就越能讲得通。虽然绝大多数环境因子不能直接改变DNA 的分子序列,但是的确控制着一批表观遗传机制,调节DNA的功能(将基因表达上调或下调),或命令我们的基因产品——蛋白质——在细胞里如何表达。
今天,这就是表观遗传学的精确定义:不管DNA序列本身,调节DNA 功能以及基因开关的分子因子。表观遗传有若干个分子过程极大地影响基因组活动,而无须改变基因本身的DNA序列。
最常见的此类过程之一是“DNA甲基化”:被称为甲基原子团(由甲烷组成)的分子成分依附于DNA,将基因打开或关闭,调节基因表达的水平。研究显示,诸如温度或情绪压力之类的环境因子可以改变DNA的甲基化,这些变化可以永久进入程序并一代代地遗传下去。这个过程就叫作表观基因代际遗传。
最近几年发现的另一个重要的表观基因过程是“组蛋白修饰”。组蛋白是依附并改变DNA结构的蛋白质,但DNA可以反过来把组蛋白绕起来,就像一串珠子那样。DNA与组蛋白的结合被称为“染色质结构”。染色质结构中回应环境压力的螺旋、圈环和扭曲也能永久改变基因表达。
最近,研究人员记录了“RNA甲基化”,甲基原子团依附于遗传辅助分子,在此过程中改变基因表达以及其后代代相传的蛋白质生产。同样,所谓“非编码RNA”的活动捆绑于RNA小分子、RNA和蛋白质,也独立于DNA序列改变基因表达。
所有这些表观基因机制都很关键,在分子调节DNA功能中起到独一无二的作用。如此看来,生物学的规则绝不会是一个“仅遗传过程”,也不是一个“仅表观遗传的过程”。相反,表观遗传学过程与遗传学完全结合。无此即无彼。
要想让表观遗传对进化论产生重大影响,它的变化必须被后代遗传,就像DNA序列和基因突变一样。可是,表观基因遗传并不遵守许多适用于经典遗传学和新达尔文进化论的孟德尔法则。这些法则将基因序列和基因功能离散,像粒子一样;在繁殖时,每个母体的“粒子”与另一母体相匹配的一对任意结合,产生一个新的基因序列和新的遗传性状表达。
相比之下,表观基因代际遗传是在生殖细胞系(精子或卵子)代际传输表观遗传信息时发生,即使没有环境暴露的持续直接影响也可以。环境压力和暴露在生殖细胞系发育期间的影响尤其大。的确,关键时期环境暴露能够通过DNA甲基化、组蛋白修饰和非编码RNA的改变引发永久性表观基因变化。
我在华盛顿州立大学的项目组于2000年辨认出的这种非基因形式的遗传证据很具说服力。我们研究小组的发现刊登在2005年的《科学》杂志上。我们的发现表明,在没有任何持续暴露的情况下,环境化学物能够促使老鼠三代甚至更久的疾病遗传。在过去的10年里,许多实验室用不同的物种进一步观察记录了这种现象。例如,联合王国南安普顿大学的格雷厄姆·伯奇及其团队报告,如果营养过剩,会造成老鼠三代及以上表观基因诱发的代谢异常。
美国得克萨斯大学奥斯汀分校的西本松及其同事发现,干旱及温度变化可引发植物表观基因变化,导致其若干代生长和开花性状变化。最近的一些研究表明,环境压力能促使遗传给后代并诱发后代病变的表观基因变化。加拿大莱斯布里奇大学的格林德·麦茨及其同事的一项最新研究证明,抑制怀孕的老鼠或者强迫其游泳,会产生表观基因损伤,危及新生幼鼠。这种祖先压力也促使被暴露妊娠期的雌性第三代表观基因代际畸形遗传。可以看到,目前的几项研究都支持环境压力在促成表观基因代际疾病遗传中起作用的观点。
据观察,环境引发植物、昆虫、鱼类、鸟类、啮齿类、猪和人类表观基因代际遗传。因此,
这是一种高度保守的现象。研究表明,表型性状变异和疾病的表观基因代际遗传在多数生物体上至少跨越10代,多数广泛研究针对几百代的植物。这里仅举一例植物。热引发的开花性状是18世纪由卡尔·林奈首次观察到的,后来发现是因为初期植物的DNA甲基化改变引起的,并持续了100代。营养变化改变的蠕虫性状可以传播5 0多代。我们发现,对寿命较长的哺乳动物来说,毒剂引发的异常性状传播近10代。在大多数这些研究中,代际性状不退化,而是持续。就连瓦丁顿的
果蝇实验也传播了16代,被改变的性状直至今天还存在。
新拉马克概念与新达尔文概念相结合,成为一种统一理论,为研究进化机制提供了更加有效的分子基础。
正如拉马克认为的那样,环境的变化的确改变了我们的生理习性。即使没有持续暴露,被改变的生理习性(表达为性状或表现在疾病的形式上)也从一代传给下一代。
环境在进化方面起着非常重要的作用。在达尔文学说的意义上,环境决定着哪些个体和物种会在自然选择的无情机制中生存下去。但是大量环境因子也能通过表观基因手段更直接地影响进化和生理特性:性状能够通过接触温度和光来转移或回应高脂肪或热限制饮食等营养参数,植物中的许多化学物或毒素也能影响表型变异及健康。
我们的研究有一项是关于环境化学物接触对性状变异和疾病的影响。研究中,我们着手调查一种环境毒剂——乙烯菌核利(农利灵,当今农业最常用的杀菌剂) ——的能力。首先,我们让怀孕的老鼠短暂接触杀菌剂;然后,在没有持续接触的情况下,让老鼠繁殖至第三代。我们观察到,该血统几乎所有雄性的精子数量减少,活力下降,还有连带的、随着年龄增大而发生不孕的现象。我们发现,不直接接触杀菌剂的三代雄性和雌性产生其他各种疾病,包括睾丸、卵巢、肾、前列腺、乳腺和大脑的畸形和异常。精子里相应的表观基因变化涉及DNA甲基化和非编码RNA表达的变化。
我们的研究表明,祖先接觸毒剂农利灵可影响到动物相同血统三代的性选择(雌雄淘汰)。自从达尔文首次提出他的学说以来,性选择(又称为择偶偏好)被视为进化的一大动力,通过允许其他雌幼鼠选择被暴露或未被暴露的雄性进行评估。压倒多数的雌鼠挑选了缺乏代际表观基因变化和祖先未被暴露的雄性。总之,暴露于杀菌剂永久性地改变了后代的精子表观基因;反过来,这导致了性选择特点的遗传,降低了基因可能在更大群体中传播的频率,直接影响微观进化规模的演变。
在最新的另一项研究中,我们调查了宏观进化规模上的演变——物种形成。物种形成的一个经典例子是加拉帕戈斯群岛上的达尔文雀。从一个种传播出来的一群雀成为不同大小的16个种,各自有其不同性状,比如说变化了的喙结构。我们的小组及合作者着手调查其中5种的DNA。我们注意到一个种群到另一个种群的DNA序列突变,但是其DNA 甲基化(表观突变)的表观基因变化数量更大,且与种群间的系统发生(系谱)距离更加相关。虽然进化领域目前集中于新达尔文遗
传概念,但是我们的发现表明,表观遗传学在物种形成和达尔文雀的演变中也有作用。
进化中的表观基因作用得到越来越多的证据支持。一项有趣的研究比较了尼安德特人和人类的DNA,其遗传差异比表观基因差异显然要小,这其中涉及基因组中DNA甲基化的变化。简言之,新拉马克概念与新达尔文概念相结合,成为一种统一理论,为研究进化机制提供了更加有效的分子基础。
新达尔文机制和新拉马克机制共同驱动进化,它们似乎相互交错。的确是这样。因为环境表观遗传机制能够增强一个群体的性状变异,赋予自然选择比其他群体增强了适应性状。在经典的新达尔文进化学说中,基因突变和基因变异是造成变异的主要分子机制。将直接影响性状变异的环境表观遗传加入这些机制,就能增强环境协调自然选择和进化的能力。
我们实验室考虑的另一个关键因素是表观遗传改变基因组稳定性的能力,它可以引发癌症生物学中观察到的一类基因突变。我们这里发现的基因突变包括后代的拷贝数变异(重复DNA短序列)和点突变(DNA序列中单核苷酸的变化)。现已知几乎所有种类的基因突变都有前体表观基因变化增加产生那种突变的敏感性。我们观察到,第一代直接环境暴露产生表观基因变化而没有基因突变,但是从代际角度来看增加了基因突变。环境表观遗传能够促进性状变异和突变,它也加速了进化机制,从其方式上看,光有达尔文的机制还不行。
有许多人怀疑统一进化论,尤其是基于影响了生物科学100多年的基因决定论范式产生的怀疑。基因决定论把DNA视为生物学的基本构件,将基因序列视为终极分子控制。
也许遗传决定论的重要标志是人类基因组的排序,为基因居于首位提供终极证据。据预测,全组基因关联研究将为生命的正常现象和非正常现象提供生物学标志,并揭示疾病的基础。但在排序之后,基因决定论的重大预测——大多数人类生物机制和疾病能够通过遗传学的镜头得到解释——没有得到证实。
接受了遗传学教育的科学家和公众一代又一代,但是没有多少人接触过表观遗传学这个相对新的科學。事实上,将表观遗传学纳入生物学的分子元素和进化学说是遭到反对的。沃森在发现DNA结构中发挥了他的作用,弗朗西斯·柯林斯在人类基因组排序中发挥了他的重大作用(他是美国国立卫生研究院的院长),这两位都曾严重怀疑表观遗传学的重要性,但是现在持更加积极的支持态度。基因决定论延续了100年以后,范式转移受到强烈抵抗不足为奇。
托马斯·库恩提出,当目前的范式表现出异常时,就需要考虑新的科学——科学革命因此诞生。
在我提出统一进化论一个月后,我的观点发表在2015年的《基因组生物学与进化学说》上。新西兰梅西大学的大卫·佩尼认为,表观遗传机制在很大程度上是遗传学的一个祖先特征,纯粹是遗传学的一个组成部分。其他最新论著,包括澳大利亚拉筹伯大学艾玛·怀特洛发表的一篇,对拉马克哺乳动物表观基因遗传的概念提出了不同的看法。
尽管产生了这些阻力,我还是深信我们已经到了范式转移的那个点。接受表观遗传学在进化学说中扮演的角色并不颠覆遗传科学,拥护新拉马克观念并没有向经典的新达尔文学说提出挑战。被接受的科学至关重要、精确,但也是一个更大的、更具细微差别的故事的一部分,它扩展了我们的理解视野,把我们所有的观察变成一个紧密结合的统一体。统一理论解释了环境如何直接影响表型变异,又如何直接促进自然选择。
随着越来越多的进化生物学者对表观遗传功能发生兴趣,现在已经有数学模型将遗传学与表观遗传学联合成一个系统,这项研究工作初见成效:将表观遗传视为另外一种分子机制有助于我们理解遗传漂变;基因同化(回应环境所产生的性状最终在基因里编码);甚至还有中性进化学说,通过中性进化的多数变化不是对自然选择的反应,而是偶然发生的。新模型为生物学家的观察提供了扩展的分子机制,为一般进化学说绘制了一个更深层次、具有更多细微区别和更加精确
的路线图。
将这些发现结合起来就要求我们搬出基因决定论这个旧标准以便发现缺口。正如托马斯·库恩于1962年提出的,当目前的范式表现出异常时,就需要考虑新的科学,科学革命也因此诞生。
统一进化论应该联合新拉马克学说和新达尔文学说,以便理解环境如何影响进化。拉马克200多年前做出的贡献不能因为达尔文而打折扣,相反,应该将两者结合起来形成一个更具影响力和远见的理论。同样,遗传学和表观遗传学不应被视为两个相互冲突的领域,相反,两者应该联合起来,为解释生命的控制机制提供更为全面的分子因子。
猜你喜欢
河北果树(2021年4期)2021-12-02
上海公路(2019年3期)2019-11-25
福建基础教育研究(2019年10期)2019-05-28
小学生必读(高年级版)(2018年12期)2018-04-12
求学·理科版(2017年3期)2017-04-27
中国中医药现代远程教育(2014年23期)2014-03-01
遗传(2014年3期)2014-02-28
遗传(2014年3期)2014-02-28
