
环境温度对中华蟾蜍间脑神经元电活动的影响
2017-07-01 08:46邓昕旸晁贺王勇王丽文梁传成
湖南师范大学学报·自然科学版 2017年3期
关键词:环境温度
邓昕旸+晁贺+王勇+王丽文+梁传成
摘 要 应用微电极电生理技术记录不同环境温度下中华蟾蜍间脑神经元电活动,观察环境温度对中华蟾蜍间脑神经元电活动的影响,探讨生理性体温调节在中华蟾蜍体温调节中的作用.结果,中华蟾蜍间脑松果体和下丘脑视前区前部神经元的放电频率和放电幅度随环境温度的升高而升高,呈兴奋性变化,相关显著,表明松果体和下丘脑视前区前部神经元对高温环境较敏感,参与了降温过程的体温调节,但下丘脑视前区前部较松果体变化明显;下丘脑视前区后部神经元的放电频率和放电幅度随温度的升高而降低,呈抑制性变化,相关显著,表明下丘脑视前区后部神经元对低温敏感,参与了升温过程的体温调节.
关键词 环境温度;中华蟾蜍;间脑;电活动;体温调节
中图分类号 Q42 文献标识码 A 文章编号 1000-2537(2017)03-0027-05
Impact of Ambient Temperature on Electrical Activities of
Bufo Gargarizans Diencephalon Neurons
DENG Xin-yanga, CHAO Hea, WANG Yongb, WANG Li-wena, LIANG Chuan-chenga*
(a.College of Life Science, b.Experiment Center, Shenyang Normal University, Shenyang 110034, China)
Abstract The objective of this study is to probe the impact of ambient temperature on physiological thermaregulation in Bufo gargarizans. The microelectrode technique was employed to record extracellularly spontaneous electrical activities of neurons in diencephalon of Bufo gargarizans at different ambient temperatures. Our results show that, as ambient temperature increases, the firing frequency, amplitude of neurons of anterior hypothalamus and pineal gland were also found increased. Significant correlations among them were observed, indicating that neurons in anterior hypothalamus and pineal gland are sensitive to the high temperature environment, and they are all involved in body temperature adjustment. We also found that changes in the anterior hypothalamus were more pronounced than those in pineal gland. The firing frequency and amplitude of neurons in posterior preoptic areas were seen decreased as the ambient temperature increased, suggesting that there was an inhibition process going on in posterior preoptic areas. This mechanism indicates that neurons in the posterior preoptic area are more sensitive to low temperatures and they play important roles in the process of fervescence.
Key words ambient temperature; Bufo gargarizans; diencephalon; electrical activities; thermoregulation
動物感知外界温度,躲避对自身有害的气候环境,调节体温,维持内环境的稳定[1].恒温动物通过下丘脑体温调定点的调节机制引起产热和散热活动,还可改变代谢速率,以维持体温的相对稳定.变温动物则通过行为性体温调节方式来维持自身的优选最适温度[2-5],它们的体温在一定范围内随环境温度的变化而变化[6].研究表明,变温动物的体温与环境温度呈正相关,但在环境温度较低时体温略高于环境温度,环境温度较高时体温又略低于环境温度,说明变温动物除具有行为性体温调节能力外,还具有一定的生理性体温调节能力[7-8].
大量的哺乳动物体温与体温调节机制研究表明其体温调节中枢位于下丘脑视前区[9-16].而有关变温动物体温的生理性调节研究相对较少,Charles等在对蜥蜴的实验研究中发现,松果复合体对环境温度的变化很敏感,可能具有温度调节作用[17].Kenia 和Lui在对两栖类和爬行类动物的实验中发现,下丘脑被损毁的牛蛙(Rana catesbeiana)其正常的体温调节行为受到干扰;注射脂多糖并不能使下丘脑视前区损毁的蟾蜍其体温发生改变[18].尹书明等人研究发现,荒漠沙蜥具有一定程度的生理性调节体温能力,下丘脑视前区参与了荒漠沙蜥对体温的调节整合过程[19].王喆等人发现荒漠沙蜥松果复合体通过调节褪黑激素的分泌影响产热和体温变化,以此来影响和参与体温调节[20].由此可见,下丘脑作为变温动物的体温调节中枢已经得到初步证实,间脑的松果体也参与了变温动物的生理性体温调节,但还缺乏足够证据.为此,作者以中华蟾蜍Bufo gargarizans为实验对象,运用微电极电生理技术,记录中华蟾蜍间脑松果体、下丘脑视前区前部和后部神经元在不同环境温度下的电活动,观察环境温度对其神经元电活动的影响,探讨其在体温调节中的作用,丰富变温动物生理性体温调节的资料.
1 材料和方法
1.1 实验动物
实验用中华蟾蜍购自沈阳医学院,为体质量35~40 g的雄性成体.室内养殖,养殖温度为(22±2) ℃,箱内水深2 cm,自然光照.隔日向養殖箱内投放面包虫.实验中每个温度梯度各用中华蟾蜍5只.实验后,实验动物上交实验中心统一处理,实验过程符合相关法律法规规定及道德伦理要求.
1.2 实验方法
1.2.1 温度控制装置 控温系统为自制的控温装置[21].该装置为一个长40 cm,宽20 cm,高15 cm,壁厚1.8 cm具有保温性的聚苯乙烯盒.盒盖上有2个圆形小孔,一个用于固定温度计实时监测装置内温度,另一个小孔用于玻璃微电极及引导电极导线进出口.盒内温度由放置不等量的冰块来控制,盒内温度能达到实验要求温度±1 ℃,每个温度梯度可维持6~9 h.
1.2.2 电生理记录 对中华蟾蜍间脑松果体、视前区前部、视前区后部3个部位神经元应用微电极电生理技术在6个不同环境温度 (5,10,15,20,25和30 ℃)的放电活动进行在体胞外记录.将用盐酸普鲁卡因局麻后的中华蟾蜍固定于解剖板上,常规手术方法快速暴露间脑.滴加任氏液后置于已调好温度的温度控制装置内,驯化1 h后进行电生理实验记录放电活动.用WF-2型微电极推进器(西北光学仪器厂)将内充3 mol/L KCl尖端直径小于1 μm、阻抗5~20 MΩ的玻璃微电极分别插入松果体、视前区前部、视前区后部3个部位.电极记录位置以松果体横向和纵向中轴线为0,松果体:0前300 μm,0后500 μm,左右300 μm,深度为0~200 μm;下丘脑视前区前部:0前200 μm,左右130 μm,深度为1 000~1 500 μm;下丘脑视前区后部:0后100 μm,左右130 μm,深度为1 050~1 450 μm.放电信号经由WF-IB型高阻微电极放大器(成都仪器厂)放大后进入RM6240型(成都仪器厂)信号采集处理系统,在计算机上观察并记录3个部位神经元的电活动.每个温度梯度记录3 h.实验后统计间脑不同部位神经元在不同温度下的放电单位数量、放电频率和放电幅度,并对环境温度与间脑神经元的电活动进行相关性分析.
1.2.3 数据分析 实验结果用Office 2010 Excel绘图,SPSS 16.0进行统计分析.数据用平均值±标准差(X±SD)表示.用单因素方差分析(one-way ANOVA)分析组间差异,多重比较采用Tukey检验.P<0.05即认为差异极显著.
2 结果
2.1 环境温度对中华蟾蜍间脑神经元电活动的影响
实验在间脑3个部位共记录了982个放电单位,其中松果体305个,下丘脑视前区前部317个,下丘脑视前区后部360个.结果显示(图1)松果体和视前区前部的变化一致,其放电活动随着环境温度的升高而增加.视前区后部神经元放电活动随环境温度的升高而降低.
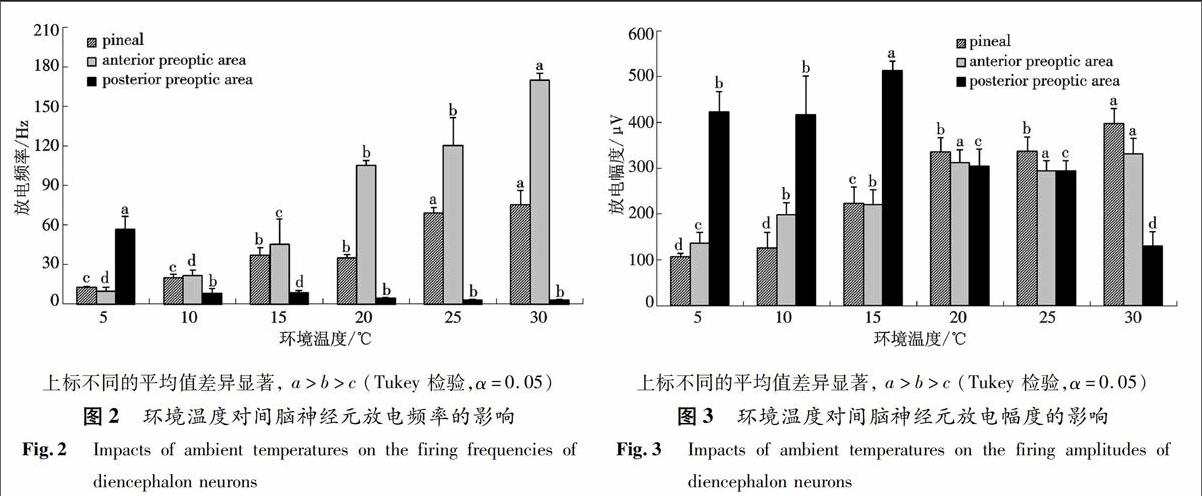
2.2 环境温度对中华蟾蜍间脑神经元放电频率和放电幅度的影响
不同环境温度下间脑3个部位神经元放电幅度和放电频率的变化见图2和图3.松果体和视前区前部神经元放电频率的变化趋势一样,随着环境温度的升高而加大.但视前区前部神经元的放电频率在环境温度达20 ℃时明显高于松果体.视前区后部神经元的放电频率则随环境温度的增高而降低.不同环境温度下3个部位神经元放电幅度的变化与放电频率的变化相似,松果体和视前区前部神经元的放电幅度随环境温度的升高而增大,视前区后部神经元的放电幅度在环境温度15 ℃时有所增大,此后则随温度的升高而降低,在30 ℃时降低显著.
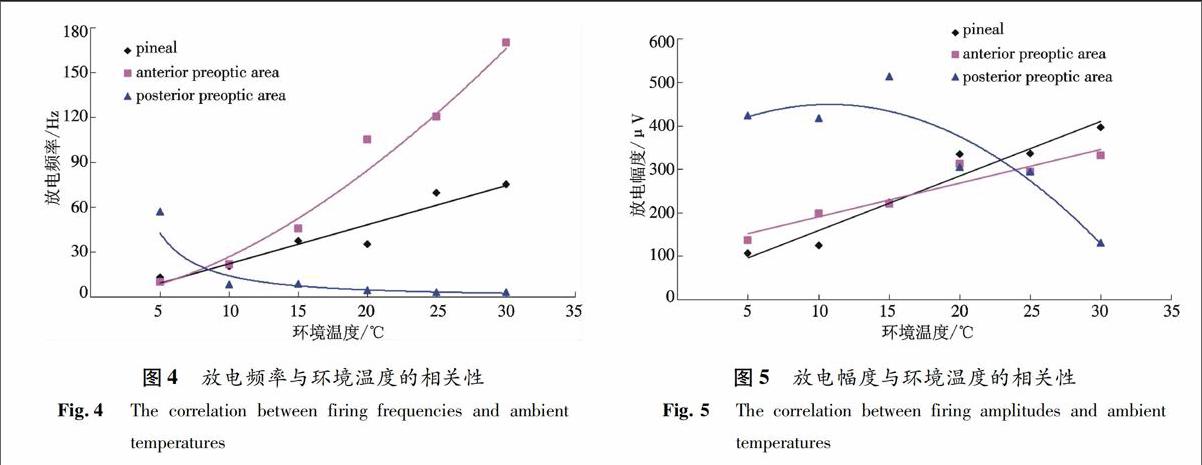
2.3 环境温度与间脑神经元电活动的相关性分析
3个部位神经元的电活动与环境温度都具有相关性,且相关显著(P<0.05).放电频率与环境温度的相关性见图4.松果体神经元的放电频率与环境温度呈直线相关,相关方程y=2.615x-4.105 6,相关系数r=0.96;视前区前部和视前区后部神经元的放电频率与环境温度均呈幂相关,相关方程和相关系数分别是y=0.589 2x1.658 7,r=0.99;y=568.2x-1.608,r=0.99;放电幅度与环境温度的相关性见图5.松果体和视前区前部神经元的放电幅度与环境温度均呈线性相关,相关方程和相关系数分别为y=12.544x+34.413,r=0.97;y=7.724 6x+113.79,r=0.95;视前区后部神经元的放电幅度与环境温度呈多项式相关,相关方程y=-0.865 6x2+18.623x+349.46,相关系数r=0.92.
3 讨论
在哺乳动物中,下丘脑被认为是体温调节的高级中枢,其中下丘脑视前区前部对机体的体温起着主导作用[22].下丘脑同样是低等脊椎动物的体温调节中枢,对维持动物体温的相对稳定起到了一定的作用[7,23].笔者在对中华蟾蜍下丘脑视前区前部及松果体神经元电生理学特性的研究中发现,随着环境温度的升高,这两个部位神经元的放电频率、放电幅度均逐渐升高,但松果体的放电幅度明显低于视前区前部.推测下丘脑视前区前部及松果体的神经元主要对高温环境较敏感,参与了降温过程中的体温调节.下丘脑视前区后部神经元的电生理特性正好与视前区前部相反,其放电幅度、放电频率均随温度的升高而降低,推测视前区后部对低温环境较为敏感,主要参与了升温过程的体温调节.这与变温动物的体温在环境温度较低时体温略高于环境温度,环境温度较高时体温又略低于环境温度相一致[6-8].且环境温度的变化与中华蟾蜍下丘脑视前区神经元的电活动相关显著,与尹书明和刘重斌的研究荒漠沙蜥的下丘脑视前区参与了荒漠沙蜥对体温的调节整合过程相一致[5,19],进一步说明变温动物一定的生理性体温调节和哺乳动物一样来自下丘脑[5,16].此外,中华蟾蜍松果体神经元的电活动与环境温度也呈显著相关,与王喆等人在荒漠沙蜥松果复合体的研究结果相似[20],表明松果体也参与了变温动物的体温调节.
此外,下丘脑视前区前部神经元可以合成、分泌和释放多种促激素、神经肤和神经递质,同时与中枢神经系统的其他部位有广泛的纤维联系[28],接受多种来自其他部位的信息分子,参与哺乳动物的体温调节[24-27].下一步工作将研究这些激素等物质是否参与了变温动物的体溫调节.
参考文献:
[1] 许云华.体温调节机制研究进展[J].连云港师范高等专科学校学报, 2010,27(2):96-98.
[2] COWLES R B, BOGERT C M. A preliminary study of the thermal requirements of desert reptiles[J].Bull Am Mus Nat Hist, 1944,83:263-296.
[3] BARTHOLOMEW G A. Physiological control of body temperature[J]. Harvey Pough Bio Reptilia, 1982,12:167-211.
[4] HUEY R B. Temperature, physiology, and the ecology of reptiles[J]. Bio Reptilia,1982,12:25-91.
[5] 刘重斌.前列腺激素和下丘脑在荒漠沙蜥体温调节中的作用[D]. 兰州:兰州大学,2006.
[6] 王培潮,陆厚基,祝龙彪,等.大蟾蜍生理生态研究Ⅰ.体温调节与热量代谢[J].两栖爬行动物学报,1984,3(1):47-53.
[7] BARTHOLOMEW G A, TUCKER V A. Control of changes in body temperature, metabolism, and circulation by the agamid lizard, amphibolurus barbatus[J]. Physiol Biochem Zool, 1963,36(3):199-218.
[8] 王丽文,梁传成,黄 薇,等.环境温度对爪鲵体温及能量代谢的影响[J].动物学报, 2008,54(4):640-644.
[9] BLIGH J. The central neurology of mammalian thermoregulation[J]. Neuroscience, 1979,4(9):1213-1236.
[10] BOULANT J A, DEAN J B. Temperature receptors in the central nervous system[J]. Ann Rev Physiol, 1986,48(1):639-654.
[12] BOULANT J A. Hypothalamic neurons: mechanisms of sensitivity to temperature a[J]. Ann New York Acad Sci, 1998,856(1):108-115.
[13] SAARELA S, REITER R J. Function of melatonin in thermoregulatory processes[J]. Life Sci, 1994,54(5):295-311.
[14] LIPTON J M, CLARK W G. Neurotransmitters in temperature control[J]. Ann Rev Physiol, 1986,48(1):613-623.
[15] HORI T, NAKASHIMA T, KOGA H, et al. Convergence of thermal, osmotic and cardiovascular signals on preoptic and anterior hypothalamic neurons in the rat[J]. Brain Rew Bull, 1988,20(6):879-885.
[16] BOULANT J A. Role of the preoptic-anterior hypothalamus in thermoregulation and fever[J]. Clin Infect Dis, 2000,31(S5):438-442.
[17] CHARLES L R, BRUCE T F, GERN W A, et al. The pineal complex and thermoregulation[J]. Biol Rev Cambridge Phil Soc, 1979,54(1):41-72.
[18] BICEGO K C, BRANCO L G. Discrete electrolytic lesion of the preoptic area prevents LPS-induced behavioral fever in toads[J]. J Exp Biol, 2002,205(22):3513-3518.
[19] 尹书明.下丘脑在荒漠沙蜥体温调节中的作用及其机制初探[D]. 兰州:兰州大学,2004.
[20] 王 喆.荒漠沙蜥松果复合体影响体温调节的生理生化机制研究[D]. 兰州:兰州大学,2004.
[21] 郭丽杰.爪鲵体温调节中枢组织学及电生理学特性的研究[D].沈阳:沈阳师范大学,2011.
[22] 廖仕元.体温调节机理的某些进展[J].中国医师进修杂志,1983(10):18-21.
[23] 马 爽, 谢惠春, 李晓晨. 两栖类和爬行类体温调节的神经生物学研究进展[J].四川动物,2008,27(3):464-467.
[24] 何子安.与体温调节有关的多肽[J].生理科学进展,1985(1):36-40.
[25] SAGONAS K, MEIRI S, VALAKOS E D, et al. The effect of body size on the thermoregulation of lizards on hot, dry Mediterranean islands[J]. J Therm Biol, 2013,38(2):92-97.
[26] HEPPNER K M, MLLER T D, KIRCHNER H, et al. The role of ghrelin-octanoyl-acyl-transferase in thermoregulation[J]. J Endocrinol Invest, 2013,36(3):180-418.
[27] HODGES M R, RICHERSON G B. The role of medullary serotonin (5-HT) neurons in respiratory control: contributions to eupneic ventilation, CO2 chemoreception, and thermoregulation[J]. J Appl Physiol, 2010,108(5):1425-32.
[28] 王 维,王欢欢,邓昕旸,等.爪鲵端脑胚后发育的组织学研究[J]. 湖南师范大学自然科学学报, 2016,39(3):18-21.
猜你喜欢
科学与财富(2021年33期)2021-05-10
山东工业技术(2017年15期)2017-09-05
科技与创新(2017年12期)2017-06-30
科技与创新(2017年10期)2017-06-07
科技创新导报(2016年36期)2017-04-20
江苏农业科学(2016年4期)2016-06-14
科技与创新(2015年21期)2015-12-01
中国高新技术企业(2015年17期)2015-05-18
科技与创新(2015年7期)2015-05-05
祝您健康(1993年1期)1993-12-28
