
秘鲁外海茎柔鱼角质颚的形态特征及其与个体大小和日龄的关系
2017-09-04 01:28胡贯宇陈新军
海洋渔业 2017年4期
胡贯宇,金 岳,陈新军,2,3
秘鲁外海茎柔鱼角质颚的形态特征及其与个体大小和日龄的关系
胡贯宇1,金 岳1,陈新军1,2,3
(1.上海海洋大学海洋科学学院,上海 201306;2.国家远洋渔业工程技术研究中心,上海 201306;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306)
角质颚是头足类重要的硬组织,被用于估算其个体大小及其生物量等。根据2009~2014年中国鱿钓船在秘鲁外海(79°22′W~84°30′W、10°00′S~18°16′S)采集的茎柔鱼(Dosidicus gigas)样本,提取出1 346 ind茎柔鱼的角质颚,并对角质颚的形态进行了分析,建立了角质颚的形态特征参数与胴长、体重和日龄之间的关系。结果显示,雌性个体角质颚各形态参数的平均值均大于雄性个体,在角质颚12个形态参数中,上喙长(UHL)、上脊突长(UCL)、上侧壁长(ULWL)、下脊突长(LCL)、下侧壁长(LLWL)、下翼长(LWL)与胴长、体重、日龄之间的相关系数较高。胴长与角质颚各形态参数的关系在雌、雄个体间的差异性均显著(P<0.05),体重、年龄与角质颚各形态参数的关系在雌、雄个体间的差异性均不显著(P>0.05)。角质颚各形态参数与胴长、日龄之间呈线性关系;除上侧壁长外,角质颚的其它形态参数与体重之间呈幂指数关系。主成分分析认为,第一主成分解释上、下角质颚形态的贡献率分别为95.69%和95.11%,在上角质颚和下角质颚的形态参数中,载荷系数最大的形态参数分别为上脊突长和下侧壁长。研究认为,上述关系式的建立可以用于茎柔鱼的个体大小、生物量和日龄的估算。
茎柔鱼;角质颚;形态特征;个体大小;日龄
茎柔鱼(Dosidicus gigas)隶属柔鱼科(Ommastrephidae)茎柔鱼属,是柔鱼科中个体最大、资源量最为丰富的种类之一[1-2]。茎柔鱼广泛分布于东太平洋,自加利福尼亚(37°N)至智利的南部(47°S),从南、北美洲的沿岸一直延伸到125°W[1]。而且,其种群结构复杂,资源丰度易受环境的影响[3]。据FAO统计,2000年以来其产量呈现上升的趋势,2012年达到历史最高产量,为95.1×104t,2013年总产量为84.7×104t。
在海洋生态系统中,头足类是甲壳类、鱼类以及软体动物的捕食者[1-4],同时又是鲸鱼、海豚、金枪鱼、海鸟等顶级捕食者的重要食物来源[5]。角质颚是头足类的主要摄食器官,具有稳定的形态特征、良好的信息储存以及耐腐蚀等特点[6],被广泛应用于种群鉴定[7-8]、分类[9]、日龄鉴定[10-11]等研究。角质颚主要由几丁质和蛋白质组成[12],由于其不易被消化而经常会出现在捕食者的胃中,因此,在已有的研究中,角质颚被广泛地应用于估算被捕食头足类种类的个体大小和生物量[5,13-14]。为此,本研究对秘鲁外海茎柔鱼角质颚的形态特性进行了分析,建立角质颚形态参数与胴长、体重和日龄之间的关系,为利用角质颚估算茎柔鱼的个体大小、生物量和日龄提供基础。
1 材料和方法
1.1 样品采集
样品采集的时间为2009~2014年,作业的海域为79°22′W~84°30′W、10°00′S~18°16′S。样本委托中国远洋专业鱿钓船在生产期间采集,每一采集站点的样本从渔获物中随机获得,每次采集30~40 ind,采集的样本冷冻后运回实验室,样本总数为1 346 ind,其中雌性个体955 ind,雄性个体391 ind。
1.2 研究方法
1.2.1 生物学测定与角质颚提取
将茎柔鱼样本带回实验室并解冻,测量茎柔鱼的胴长(mm)、体重(g),对其性别进行鉴别。角质颚位于头部口器中,将角质颚取出后,先用水进行清洗,尽量去除附在表面的有机物质,并将其保存在70%的乙醇溶液中。在本研究中,取出了1 253个完整的上角质颚和1 301个完整的下角质颚,并对其进行形态学测量。
1.2.2 角质颚的形态测量
将角质颚清洗干净后,用游标卡尺对其进行测量。首先沿水平方向和垂直方向进行校准,然后对角质颚的外部形态进行测量,测量结果精确至0.01 mm。形态参数包括12个(图1),分别为上头盖长(Upper Hood length,UHL);上脊突长(Upper crest length,UCL);上喙长(Upper rostrum length,URL);上喙宽(Upper rostrum width,URW);上侧壁长(Upper lateral wall length,ULWL);上翼长(Upper wing length,UWL);下头盖长(Lower hood length,LHL);下脊突长(Lower crest length,LCL);下喙长(Lower rostrum length,LRL);下喙宽(Lower rostrum width,LRW);下侧壁长(Lower lateral wall length,LLWL);下翼长(Lower wing length,LWL)。
1.2.3 角质颚研磨与轮纹计数
角质颚微结构被认为可以用来估算茎柔鱼的日龄[16],本研究通过分析上角质颚喙部矢状切面上的生长纹对茎柔鱼进行日龄估算(图2),通过参考PERALES-RAYA等[17]的方法来制作角质颚的切片。每一个角质颚的生长纹由两个观察者分别计数一次,两者计数的轮纹数与均值的差值低于10%,则认为准确,否则计数无效[18]。

图1 角质颚外部形态测量示意图(引自FANG等[15])Fig.1 Scheme of themeasurement for the external morphology of beak(cited from FANG et al[15])
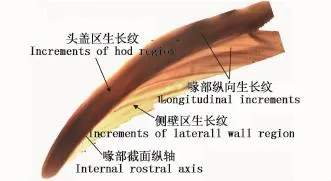
图2 秘鲁外海茎柔鱼上角质颚微结构Fig.2 Upper beak m icrostructure of D.gigas caught off the Peruvian EEZ
1.2.4 数据处理
(1)利用方差分析(ANOVA)法检验茎柔鱼的胴长、体重、日龄在雌、雄个体间的差异性。
(2)利用典型相关分析,分析胴长、体重、日龄与角质颚各形态参数之间的相关性。
(3)对上、下角质颚的形态参数分别进行主成分分析,探讨不同形态参数表征角质颚形态的差异。
(4)利用协方差分析,以雌、雄组为协变量,分析角质颚各形态参数与胴长、体重、日龄的关系在雌、雄个体间的差异性。
(5)利用线性、指数、幂指数、Logistic模型来拟合角质颚各形态参数与胴长、体重、日龄的关系[19-20],并利用R2和AIC(Akaike information criterion)值选择最佳模型,其中R2最大和AIC值最小的模型为最佳模型[21-22]。
2 结果与分析
2.1 胴长、体重和日龄组成
雌、雄个体的胴长范围分别为202~545 mm和192~534 mm,平均胴长分别为(331.55± 76.48)mm和(298.43±67.60)mm。雌、雄个体的体重范围分别为207~5 104 g和204~4 278 g,平均体重分别为(1 282.60±987.32)g和(882.02±746.41)g。雌、雄个体的日龄范围分别为123~402 d和106~369 d,平均日龄分别为(229.78±59.03)d和(198.20±47.77)d。利用ANOVA对雌、雄个体的胴长、体重以及日龄进行差异性分析,结果显示,胴长、体重和日龄在雌、雄个体间的差异性均极显著(P<0.01),且雌性个体胴长、体重和日龄的平均值均大于雄性个体。
2.2 典型相关分析
采用典型相关分析法,分析胴长、体重、日龄与角质颚各形态参数的相关性。结果显示,胴长、体重、日龄与角质颚各形态参数的相关性均均达到了极显著水平(P<0.01),而且,胴长和体重与角质颚各形态参数的相关系数均达到0.9以上(表1)。在上角质颚6个形态参数中,上喙长(UHL)、上脊突长(UCL)、上侧壁长(ULWL)与胴长、体重、日龄的相关系数均较高;在下角质颚6个形态参数中,下脊突长(LCL)、下侧壁长(LLWL)、下翼长(LWL)与胴长、体重、日龄的相关系数均较高(表1)。
2.3 主成分分析
对上角质颚和下角质颚的形态参数分别进行主成分分析,结果显示,第一主成分解释上、下角质颚形态的贡献率分别为95.69%和95.11%(表2~3),因此,第一主成分即可代表上、下角质颚的外部形态特征。上、下角质颚第一主成分与角质颚各形态参数均存在较大的相关性,载荷系数均在0.96~0.99之间。在上角质颚各形态参数中,上脊突长(UCL)的载荷系数最大,达到0.988 4;在下角质颚各形态参数中,下侧壁长(LLWL)的载荷系数最大,达到0.987 3。

表1 秘鲁外海茎柔鱼角质颚各形态参数与胴长、体重、日龄的Pearson相关系数Tab.1 Pearson correlation coefficient between themorphological param eters of beak and mantle length,body weight and daily age for D.gigas caught off the Peruvian EEZ
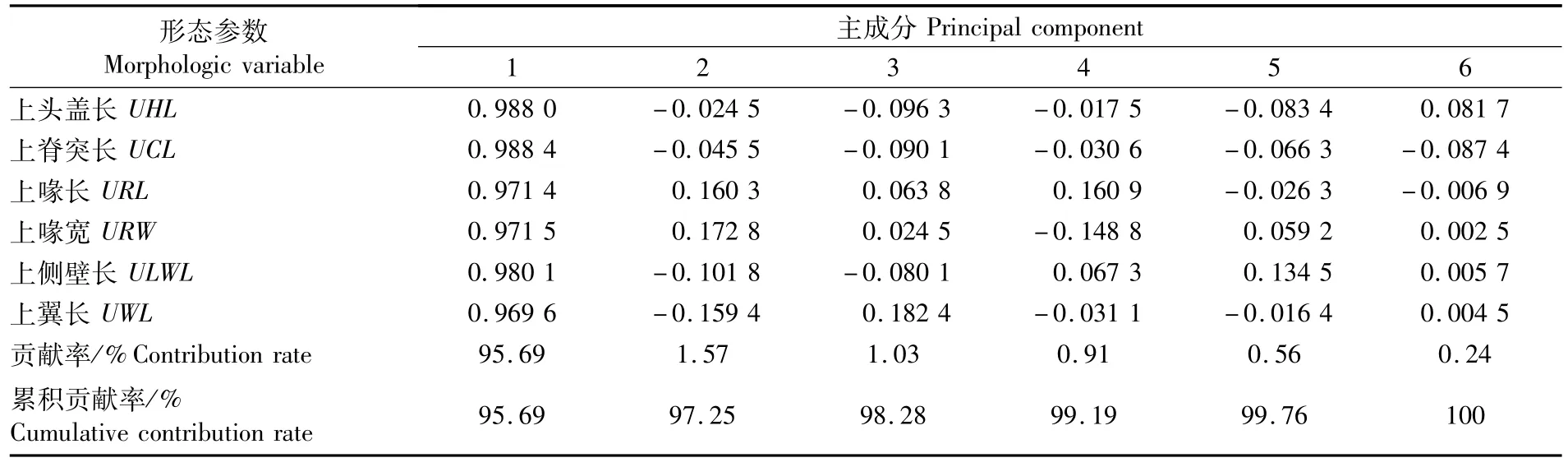
表2 秘鲁外海茎柔鱼上角质颚形态参数的主成分分析Tab.2 Principal component analysis for themorphological parameters of upper beak of D.gigas caught off the Peruvian EEZ
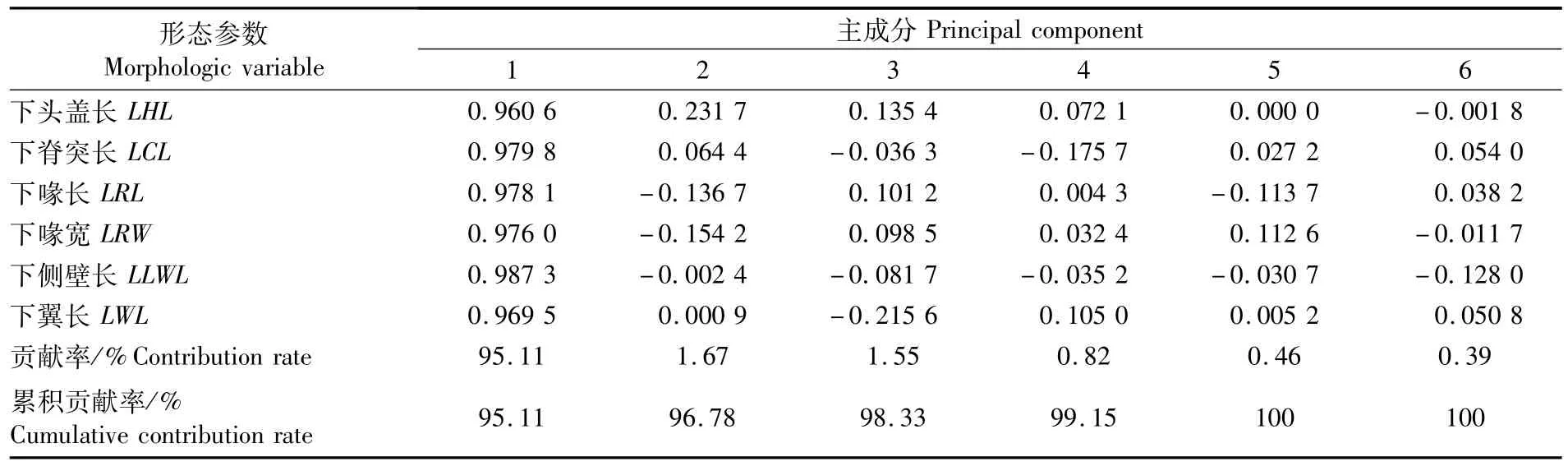
表3 秘鲁外海茎柔鱼下角质颚形态参数的主成分分析Tab.3 Principal component analysis for themorphologic parameters of lower beak of D.gigas caught off the Peruvian EEZ
2.4 角质颚外部形态参数与胴长、体重、年龄的关系
综合考虑典型相关分析和主成分分析的结果,选取上喙长(UHC)、上脊突长(UCL)、上侧壁长(ULWL)、下脊突长(LCL)、下侧壁长(LLWL)和下翼长(LWL)与胴长、体重、年龄进行关系建立。利用协方差分析法,检验胴长、体重、年龄与角质颚各形态参数的关系在雌、雄个体间的差异性。结果显示,胴长与角质颚各形态参数的关系在雌、雄个体间的差异性均显著(P<0.05),因此,将雌、雄个体的数据分开进行分析。然而,体重、年龄与角质颚各形态参数的关系在雌、雄个体间的差异性均不显著(P>0.05),因此,可将雌、雄个体的数据合并起来。
根据R2和AIC值对不同方程进行选择,角质颚各形态参数与胴长、日龄的关系均较为复合线性相关关系(图3A-F,M-R);上侧壁长与体重较为复合指数关系,角质颚其它形态参数与体重的关系均较为复合幂指数关系(图3G-I)。
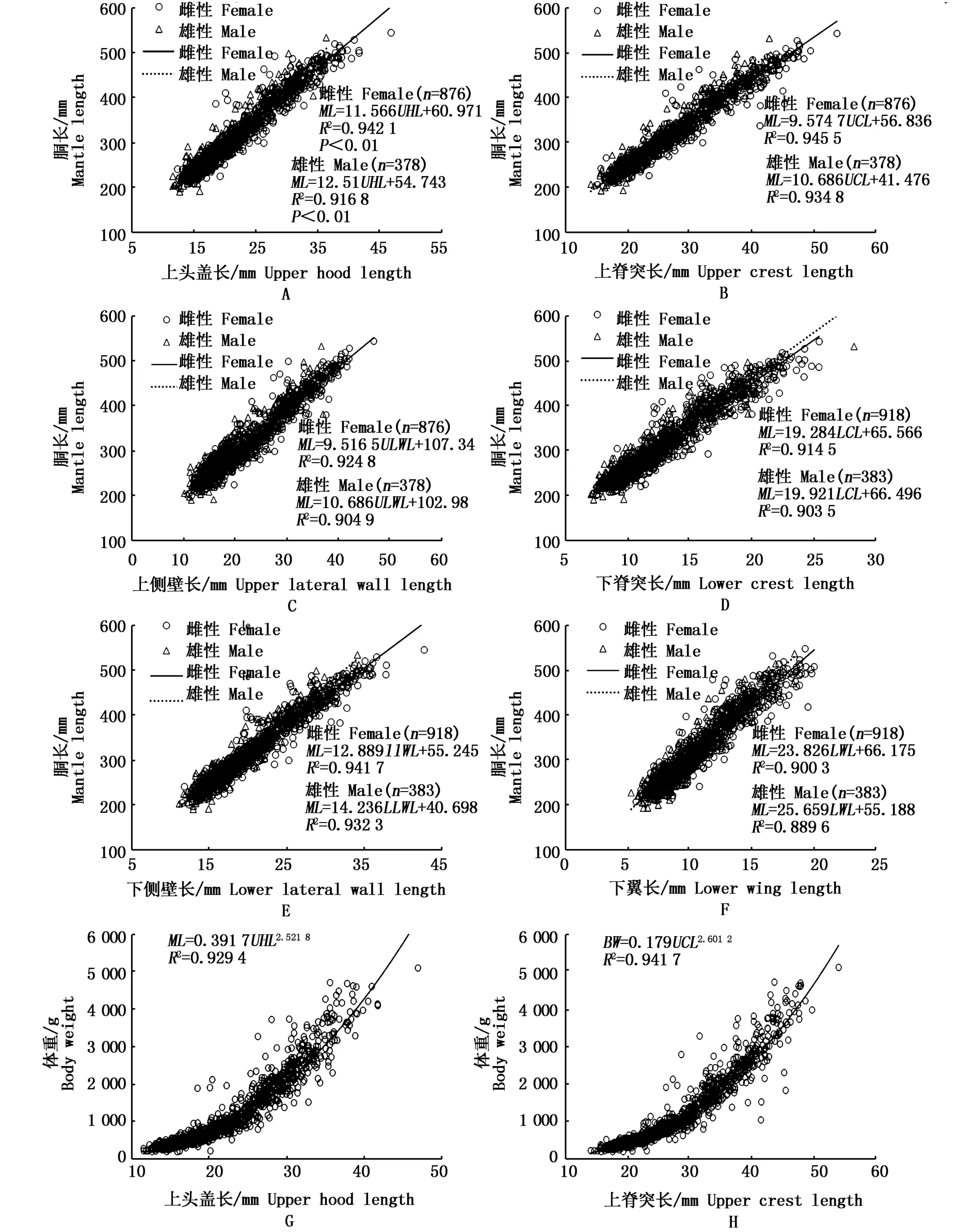

图3 秘鲁外海茎柔鱼角质颚形态参数与胴长、体重、年龄的关系Fig.3 Relationship between the beak morphological parameters and mantle length,body weight,and daily age of D.gigas caught off the Peruvian EEZ
3 讨论
3.1 样本的胴长、体重和日龄组成
一般认为,茎柔鱼的生命周期约为1年,然而,大个体群体中的一些个体大的茎柔鱼(ML>750 mm),生命周期可达1.5~2年[1]。本研究通过分析上角质颚微结构生长纹估算秘鲁外海茎柔鱼的年龄,结果表明,雌、雄个体的最大日龄分别为402 d和369 d,所对应的胴长分别为485 mm和428 mm,所对应的体重分别为3 877.1 g和2 710.9 g。MEJIA-REBOLLO等[23]通过分析耳石微结构对下加利福尼亚西部沿岸海域茎柔鱼的日龄进行了估算,发现雌、雄个体的最大日龄分别为433 d和391 d,所对应的胴长分别为700 mm和690 mm。CHEN等[24]利用耳石微结构估算了哥斯达黎加外海茎柔鱼的日龄,研究发现,雌、雄个体的最大日龄分别为289 d和240 d,所对应的胴长分别为429 mm和352 mm。
3.2 角质颚的生长与摄食
在海洋生态系统中,茎柔鱼是主动的捕食者,主要捕食的种类为浮游动物、甲壳类、头足类和鱼类[1]。而且,茎柔鱼嗜食同类的现象是普遍存在的,在加利福尼亚海湾,如果被捕食者很稀少,茎柔鱼所嗜食同类的重量可达到其食物总重量的30%[25]。头足类在生长的过程中会发生食性的变化,而角质颚是头足类的重要摄食器官,因此,角质颚形态的变化代表着被捕食者的大小和种类的转变[26-27]。本研究发现,随着个体的生长,茎柔鱼的角质颚也不断地变大,为茎柔鱼能够摄食更大个体的食物提供保障。此外,随着角质颚的色素沉着的加深,角质颚变得更为坚硬,从而使头足类捕食更大、更强壮、更坚硬的食物,角质颚色素沉着的程度被认为是反映食性转变的重要特征[28-29]。
从本研究主成分分析的结果来看,第一主成分认为是角质颚各形态特征的代表,在上角质颚各形态参数中,上头盖长、上脊突长和上侧壁长的载荷系数均较大,其中以上脊突长为最大;在下角质颚各形态参数中,下脊突长、下喙长和下侧壁长的载荷系数较大,其中以下侧壁长为最大。因此,可以认为茎柔鱼角质颚的生长主要在脊突和侧壁,这与方舟等[30]的研究结果相似。随着茎柔鱼的不断生长,其角质颚也不断地变大,所捕食的食物也变得更大更坚硬,因此,需要其具有强大的咬合力才能捕获并且撕碎猎物[30]。角质颚的脊突和侧壁被肌肉所覆盖,随着脊突和侧壁的不断生长,肌肉与其接触面积不断增大,使茎柔鱼的咬合力逐渐加强,从而保证茎柔鱼能够捕获更大、更坚硬的食物。
3.3 角质颚形态参数与胴长、体重、日龄的关系
对茎柔鱼角质颚的形态参数进行比较,发现雌性个体角质颚各形态参数的均值均大于雄性,除下脊突长外,雌性个体角质颚的其它形态参数的最大值也均大于雄性[31],在本研究中,茎柔鱼雌性个体的平均胴长和平均体重均大于雄性个体,因此,这可能是由于茎柔鱼角质颚的生长与其个体的生长呈正相关关系(图3),即较大个体的茎柔鱼具有较大的角质颚。
本研究发现,茎柔鱼角质颚各形态参数与体重和日龄的关系在雌、雄个体间的差异性均不显著(P>0.05),然而,角质颚各形态参数与胴长的关系在雌、雄个体间的差异性均显著(P<0.05)。陆化杰等[32]研究认为,阿根廷滑柔鱼(Illex argentinus)角质颚的形态参数与日龄的关系在雌、雄个体间的差异性不显著。JACKSON等[33]研究了巴塔哥尼亚大陆架福克兰群岛海域的强壮桑椹乌贼(Moroteuthis ingens),发现其角质颚的形态参数与胴长和体重在雌、雄个体间的差异性均不显著。BOLSTAD[20]研究发现,强壮桑葚乌贼角质颚的生长在雌、雄个体间具有差异性。胡贯宇等[31]研究发现,茎柔鱼在不同生长阶段之间,雌、雄个体角质颚的生长存在差异,这种差异可能是由于雌、雄个体在生长发育过程中摄食习性的不同所造成的。
LEFKADITOU等[34]对色雷斯海的尖盘爱尔斗蛸(Eledone cirrhosa)角质颚的形态进行了研究,认为角质颚各形态参数与胴长、体重呈幂指数关系。JACKSON等[33]和KASHIWADA等[35]分别研究了强壮桑椹乌贼和乳光枪乌贼(Loligo opalescens)的角质颚形态特征,发现角质颚各形态参数与胴长、体重均较为复合线性关系。BOLSTAD[20]研究认为,强壮桑椹乌贼角质颚的形态参数与胴长呈线性关系,与体重呈幂指数关系。LU等[5]研究了75种头足类种类的角质颚,并根据角质颚的形态和色素沉着特征,建立了头足类角质颚鉴定检索表,同时对角质颚形态参数与胴长、体重进行了相关性分析。WOLFF[36]对太平洋18种头足类的角质颚进行了种类鉴定,同样分析了角质颚形态参数与胴长、体重之间的关系。PERALES-RAYA等[17]研究发现,真蛸(Octopus vulgaris)角质颚微结构的生长纹数与上头盖长呈幂指数关系。本研究认为,茎柔鱼角质颚各形态参数与胴长、日龄之间为线性相关关系;除上侧壁长外,角质颚其它形态参数与体重之间为幂指数关系。因此,不同头足类种类角质颚的形态特征有所不同,角质颚的生长也不同,其生长方程也会有所差异。
头足类是许多海洋动物的重要食物来源,它们坚硬的角质颚经常会出现在捕食者的胃中,因此头足类的角质颚被应用于种类鉴定以及个体大小和生物量的估算[5,36,38]。GROÉGER等[39]通过分析角质颚的形态参数,估算了寒海乌贼(Psychroteuthis glacialis)的个体大小和资源量。LU等[5]对澳大利亚南部海域头足类的角质颚进行了种类鉴定并利用角质颚对资源量进行了估算。LALAS等[14]利用角质颚估算了毛利蛸(Macroctopusmaorum)的个体大小。JACKSON[40]利用角质颚估算了新西兰海域强壮桑葚乌贼的资源量。同样,茎柔鱼也是许多大型鱼类、海鸟以及海洋哺乳动物的重要捕食对象,在海洋生态系统中具有重要地位[1]。本研究分析了茎柔鱼角质颚的形态参数与胴长、体重以及日龄之间的关系,结果表明角质颚各形态参数与胴长、体重和日龄之间的相关系数均较高,因此,可以利用角质颚估算茎柔鱼的个体大小、日龄以及被捕食的生物量,对东南太平洋食物链和食物网的研究具有重要意义。
4 展望
本研究分析了秘鲁外海茎柔鱼角质颚的形态特征及各形态参数与胴长、体重和日龄的关系。但本研究所采集的样本并没有包含所有的生活史阶段,特别是小个体的茎柔鱼样本,因此,在今后的研究中,应利用多种渔具(如围网、脱网等)采集不同发育阶段的个体对本研究加以补充。此外,地理区域不同,茎柔鱼的生长也会不同[21,23,24,41],其角质颚的生长可能也会有所差异,因此,在今后的研究中,应采集不同地理区域的茎柔鱼,并对其角质颚生长进行比较分析,从而研究不同地理区域茎柔鱼的角质颚对食性的影响。
[1] NIGMATULLIN CM,NESISK,ARKHIPKIN A.A review of the biology of the jumbo squid Dosidicus gigas(Cephalopoda:Ommastrephidae)[J].Fisheries Research,2001,54(1):9-19.
[2] TAIPE A,YAMASHIRO C,MARIATEGUI L,et al.Distribution and concentrations of jumbo flying squid(Dosidicus gigas)off the Peruvian coast between 1991 and 1999[J].Fisheries Research,2001,54(1):21-32.
[3] WALUDA C M,YAMASHIRO C,RODHOUSE P G.Influence of the ENSO cycle on the light-fishery for Dosidicus gigas in the Peru Current:an analysis of remotely sensed data[J].Fisheries Research,2006,79(1):56-63.
[4] ARKHIPKIN A,BIZIKOV V.Statolith in accelerometers of squid and cuttlefish[J].Ruthenica,1998,8(1):81-84.
[5] LUC,ICKERINGILLR.Cephalopod beak identification and biomass estimation techniques:tools for dietary studies of southern Australian finfishes[M].Melbourne:Museum Victoria,2002.
[6] CLARKEM R.The identification of cephalopod beaks and the relationship between beak size and total body weight[M].London:British Museum,1962.
[7] BORGEST.Discriminantanalysisofgeographic variation in hard structures of Todarodes sagittatus from the North Atlantic[C]//ICES Marine Science Symposia:Copenhagen,Denmark:International Council for the Exploration of the Sea,1995(199):433-440.
[8] LIU B,FANG Z,CHEN X,etal.Spatial variations in beak structure to identify potentially geographic populations of Dosidicus gigas in the Eastern Pacific Ocean[J].Fisheries Research,2015(164):185-192.
[9] OGDEN R S,ALLCOCK A,WATS P,et al.The role of beak shape in octopodid taxonomy[J].South African Journal ofMarine Science,1998,20(1):29-36.
[10] HERNÁNDEZ-LÓPEZ JL,CASTRO-HERNÁNDEZ J J,HERNÁNDEZ-GARCÍA V.Age determined from the daily deposition of concentric rings on common octopus(Octopus vulgaris)beaks[J].Fishery Bulletin-National Oceanic and Atmospheric Administration,2001,99(4):679-684.
[11] CASTANHARIG,TOMÁS A R G.Beak increment counts as a tool for growth studies of the common octopus Octopus vulgaris in southern Brazil[J].BOLETIM DO INSTITUTO DE PESCA,São Paulo,2012,38(4):323-331.
[12] MISEREZA,LIY,WAITE JH,etal.Jumbo squid beaks:Inspiration for design of robust organic composites[J].Acta Biomaterialia,2007,3(1):139-149.
[13] JACKSON G D,MCKINNON J F.Beak length analysis of arrow squid Nototodarus sloanii(Cephalopoda:Ommastrephidae)in southern New Zealand waters[J].Polar biology,1996,16(3):227-230.
[14] LALAS C.Estimates of size for the large octopus Macroctopusmaorum from measures of beaks in prey remains[J].New Zealand Journal of Marine and Freshwater Research,2009,43(2):635-642.
[15] FANG Z,LIU B,LIJ,et al.Stock identification of neon flying squid(Ommastrephes bartramii)in the North Pacific Ocean on the basis of beak and statolith morphology[J].Scientia Marina,2014,78(2):239-248.
[16] 胡贯宇,陈新军,刘必林,等.茎柔鱼耳石和角质颚微结构及轮纹判读[J].水产学报,2015,39(3):361-370.HU G Y,CHEN X J,LIU B L,et al.Microstructure of statolith and beak for Dosidicus gigas and its determination of growth increments[J].Journal of Fisheries of China,2015,39(3):361-370.
[17] PERALES-RAYA C,BARTOLOMÈA,GARCÍASANTAMARÍA M T,et al.Age estimation obtained from analysis of octopus(Octopus vulgaris Cuvier,1797)beaks:Improvements and comparisons[J].Fisheries Research,2010,106(2):171-176.
[18] YATSU A,MIDORIKAWA S,SHIMADA T,et al.Age and growth of the neon flying squid,Ommastrephes bartrami,in the North Pacific Ocean[J].Fisheries Research,1997,29(3):257-270.
[19] IVANOVIC M,BRUNETTI N.Description of Illex argentinus beaks and rostral length relationshipswith size and weight of squids[J].Revista De Investigacion Y Desarrollo Pesquero,1997(11):135-144.
[20] BOLSTAD K.Sexual dimorphism in the beaks of Moroteuthis ingens Smith,1881(Cephalopoda:Oegopsida:Onychoteuthidae)[J].New Zealand Journal of Zoology,2006,33(4):317-327.
[21] CHEN X,LU H,LIU B,et al.Age,growth and population structure of jumbo flying squid,Dosidicus gigas,based on statolith microstructure off the Exclusive Economic Zone of Chilean waters[J].Journal of the Marine Biological Association of the United Kingdom,2011,91(1):229-235.
[22] LIU B,CHEN X,CHEN Y,etal.Age,maturation,and population structure of the Humboldt squid Dosidicus gigas off the Peruvian Exclusive Economic Zones[J].Chinese Journal of Oceanology and Limnology,2013(31):81-91.
[23] MEJIA-REBOLLO A,QUIÑÓNEZ-VELÁZQUEZ C,SALINAS-ZAVALA C A,et al.Age,growth and maturity of jumbo squid(Dosidicus gigas d’Orbigny,1835)off the western coast of the Baja California Peninsula[J].California Cooperative Oceanic Fisheries Investigations,2008(49):256-262.
[24] CHEN X,LI J,LIU B,et al.Age,growth and population structure of jumbo flying squid,Dosidicus gigas,off the Costa Rica Dome[J].Journal of the Marine Biological Association of the United Kingdom,2013,93(2):567-573.
[25] EHRHARDT N M.Potential impact of a seasonal migratory jumbo squid(Dosidicus gigas)stock on a Gulf of California sardine(Sardinops sagax caerulea)population[J].Bulletin of Marine Science,1991,49(1-2):325-332.
[26] CASTRO J,HERNÁNDEZ-GARCÍA V.Ontogenetic changes inmouth structures,foraging behaviour and habitat use of Scomber japonicus and Illex coindetii[J].Scientia Marina(Espana),1995(59):347-355.
[27] FRANCO-SANTOSR,VIDAL E.Beak development of early squid paralarvae(Cephalopoda:Teuthoidea)may reflect an adaptation to a specialized feedingmode[J].Hydrobiologia,2014,725(1):85-103.
[28] HERNAN'DEZ-GARCÍA V,PIATKOWSKIU,CLARKE M.Development of the darkening of Todarodes sagittatus beaks and its relation to growth and reproduction[J].South African Journal of Marine Science,1998,20(1):363-373.
[29] GARCÍA V H.Growth and pigmentation process ofthe beaks of Todaropsis eblanae(Cephalopoda:Ommastrephidae)[J].Berliner Palaobiol Abh,Berlin,2003(3):131-140.
[30] 方 舟,陈新军,陆化杰,等.北太平洋两个柔鱼群体角质颚形态及生长特征[J].生态学报,2014,34(19):5405-5415.
FANG Z,CHEN X J,LU H J,et al.Morphology and growth of beaks in two cohorts for neon flying squid(Ommastrephes batramii)in the northwest Pacific Ocean[J].Acta Ecologica Sinica,2014,34(19):5405-5415.
[31] 胡贯宇,陈新军,方 舟.个体生长对秘鲁外海茎柔鱼角质颚形态变化的影响[J].水产学报,2016,40(1):36-44.
HU G Y,CHEN X J,FANG Z.Effect of individual growth on beak morphometry of jumbo flying squid,Dosidicus gigas off the Peruvian Exclusive Economic Zone[J].Journal of Fisheries of China,2016,40(1):36-44.
[32] 陆化杰,陈新军,方 舟.西南大西洋阿根廷滑柔鱼2个不同产卵群间角质颚外形生长特性比较[J].中国海洋大学学报:自然科学版,2012,42(10):33-40.
LU H J,CHEN X J,FANG Z.Comparison of the beak morphologic growth charateristics between two spawning polutions of Illex argentinus in Southwest Atlantic Ocean[J].Periodical of ocean university of China:Natural Science Edition,2012,42(10):33-40.
[33] JACKSON G D,BUXTON N G,GEORGE M J.Beak length analysis of Moroteuthis ingens(Cephalopoda:Onychoteuthidae)from the Falkland Islands region of the Patagonian shelf[J].Journal of the Marine Biological Association of the United Kingdom,1997(77):1235-1238.
[34] LEFKADITOU E,BEKAS P.Analysis of beak morphometry of the horned octopus Eledone cirrhosa(Cephalopoda:Octopoda)in the Thracian Sea(NE Mediterranean)[J].Mediterranean Marine Science,2004,5(1):143-150.
[35] KASHIWADA J,RECKSIEK C W,KARPOV K.Beaks of the market squid,Loligo opalescens,as tools for predator studies[J].Califomia Cooperatiove Oceanic Fisheries Investigations,1979(20):65-69.
[36] WOLFF G A.Identification and estimation of size from the beaks of18 species of cephalopods from the Pacific Ocean[M].US Department of Commerce,National Oceanic and Atmospheric Admiministration,National Marine Fisheries Service,1984.
[37] KLAGESN,COOPER J.Diet of the Atlantic petrel Pterodroma incerta during breeding at South Atlantic Gough Island[J].Marine Ornithology,1997,25(1-2):13-16.
[38] PIATKOWSKI U,PÜTZ K,HEINEMANN H.Cephalopod prey of king penguins(Aptenodytes patagonicus)breeding at Volunteer Beach,Falkland Islands,during austral winter 1996[J].Fisheries Research,2001,52(1):79-90.
[39] GROÈGER J,PIATKOWSKIU,HEINEMANN H.Beak length analysis of the Southern Ocean squid Psychroteuthis glacialis(Cephalopoda:Psychroteuthidae)and its use for size and biomass estimation[J].Polar Biology,2000,23(1):70-74.
[40] JACKSON G D.The use of beaks as tools for biomass estimation in the deepwater squid Moroteuthis ingens(Cephalopoda:Onychoteuthidae)in New Zealand waters[J].Polar Biology,1995,15(1):9-14.
[41] MARKAIDA U,QUIÑÓNEZ-VELÁZQUEZ C,SOSANISHIZAKI O.Age,growth and maturation of jumbo squid Dosidicus gigas(Cephalopoda:Ommastrephidae)from the Gulf of California,Mexico[J].Fisheries Research,2004,66(1):31-47.
Beak morphological characteristics of Dosidicus gigas off the Peruvian Exclusive Econom ic Zone(EEZ)and their relationship w ith body size and daily age
HU Guan-yu1,JIN Yue1,CHEN Xin-jun1,2,3,4
(1.College of Marine Sciences of Shanghai Ocean University Shanghai 201306,China;2.National Distant-water Fisheries Engineering Research Center,Shanghai Ocean University,Shanghai 201306,China;3.Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources,Shanghai Ocean University,Ministry of Education,Shanghai 201306,China)
Beak is an important hard structure of cephalopod used for estimating the individual size and biomass.In this study,samples of jumbo flying squid(Dosidicus gigas)were collected by the Chinese squid jigger fleet during 2009-2014 off the Peruvian Exclusive Economic Zone(79°22′W-84°30′W,10°00′S-18°16′S).The beaks were sampled from 1 346 ind and the morphologies of these beaks were analyzed.Moreover,the relationships between the morphological parameters and mantle length,body weight and daily age were built.The results showed that the average values of the beak morphological parameters of females were greater than those ofmales.Among the 12 beak morphological parameters,the upper crest length,the upper rostral length,the upper lateral wall length,the lower crest length,the lower lateral wall length,the lower wing length were highly correlated with the mantle length,the body weight and the daily age.The relationships between the beak morphological parameters and themantle length were best described by linear models,and their were significant differences between females and males(P<0.05).The linear functions were optimal to describe the relationships between the beakmorphological parameters and daily age,and there was no significant difference between females andmales(P>0.05).Except for the upper lateralwall length,the other beak morphological parameters were best fitted with the body weight by power models and no significant difference was found between females and males(P<0.05).The principle component analysis indicated that the contribution rate of the first principle component of upper and lower beak morphological parameterswere 95.69%and 95.11%,respectively.The beak morphological parameterswith themaximum load coefficientwere upper crest length(UCL)and lower lateral wall length(LLWL).The relation formula established above can be used for estimating the size,biomass and daily age of Dosidicusgigas in this region.
Dosidicus gigas;beak;morphological characteristics;body size;daily age
Q 959.216
A
1004-2490(2017)04-0361-11
2016-09-12
国家自然科学基金(NSFC41476129)
胡贯宇(1991-),男,研究生,安徽宿州人,研究方向:海洋渔业生物学。E-mail:huguanyu@126.com
陈新军,教授。E-mail:xjchen@shou.edu.cn
猜你喜欢
小哥白尼(野生动物)(2021年9期)2022-01-17
好孩子画报(2020年10期)2020-11-02
故事作文·低年级(2020年10期)2020-10-21
数学小灵通(1-2年级)(2020年6期)2020-06-24
儿童故事画报·智力大王(2020年1期)2020-04-28
养猪(2020年1期)2020-02-19
今日农业(2019年11期)2019-08-15
国外畜牧学·猪与禽(2018年11期)2018-05-14
文物保护与考古科学(2016年1期)2016-04-16
农村农业农民·B版(2015年9期)2015-10-16
