
三种木蠹蛾越冬幼虫的过冷却能力差异比较
2017-10-09 00:52徐丽丽宗世祥
植物保护 2017年5期
田 斌, 徐丽丽, 王 涛, 宗世祥*
(1.北京林业大学林木有害生物防治北京市重点实验室, 北京 100083;2.北京市门头沟区林业工作站, 北京 102300)
三种木蠹蛾越冬幼虫的过冷却能力差异比较
田 斌1, 徐丽丽1, 王 涛2, 宗世祥1*
(1.北京林业大学林木有害生物防治北京市重点实验室, 北京 100083;2.北京市门头沟区林业工作站, 北京 102300)
以宁夏地区3种木蠹蛾越冬幼虫(榆木蠹蛾、沙蒿木蠹蛾和沙棘木蠹蛾)为研究对象,分析了3种木蠹蛾越冬幼虫在不同时期的过冷却点和结冰点,同时比较了沙棘木蠹蛾不同虫龄阶段的过冷却点和结冰点。结果表明,3种木蠹蛾的过冷却能力基本相似,沙蒿木蠹蛾在3种木蠹蛾中对低温环境的适应能力较强。低温驯化对木蠹蛾越冬幼虫的过冷却能力影响明显,榆木蠹蛾和沙蒿木蠹蛾2013年3月的过冷却点及结冰点显著高于2014年1月,两种木蠹蛾表现出耐结冰型昆虫的特征。沙棘木蠹蛾各虫龄阶段之间的过冷却能力差异不显著,但高龄越冬幼虫具有更好的适应温度变化的能力。
木蠹蛾; 越冬幼虫; 过冷却点; 结冰点
昆虫是变温动物,温度对昆虫的影响至关重要。其中低温作为重要的环境因素直接影响着昆虫的生长、繁殖和扩散。对于北半球的昆虫来讲,明确昆虫的耐寒性对于监测种群动态,预测向北扩散的可能性以及害虫防治方面起到重要作用[1]。昆虫的耐寒性早在1927年就有报道,过冷却点的发现为耐寒性的研究开辟了新的领域[2-3]。此后,过冷却点作为界定昆虫的耐寒能力以及耐寒策略分析的一项重要指标,被广泛应用于昆虫种群的发生动态及地理分布的预测研究。
根据昆虫的过冷却点与致死温度的关系,传统的耐寒策略将昆虫分为两类:一类是不耐结冰型(freezing intolerant),此类昆虫通过降低自身的过冷却点使体液避免结冰,抵御冷伤害,对于此类昆虫而言,通常认为过冷却点为其生存下限温度;另一类为耐结冰型(freezing tolerant),这一类昆虫通常具有较高的过冷却点,温度下降时主动形成冰核物质促使细胞外液先结冰,从而阻止细胞内液结冰,以此避免更大的损伤[4]。此后,Bale在此基础上又将依赖过冷却能力的不耐结冰型昆虫再细化,总结出耐结冰型(freeze-tolerance)、避免结冰型(freeze-avoidance)、耐受寒冷型(chill-tolerance)、冷敏感型(chill susceptible)和机会主义型(opportunistic survival) 5种类型,将所有昆虫的耐寒性对策根据定义和生态相关标准归入一个明确的范畴[5]。
木蠹蛾科属于鳞翅目Lepidoptera,是重要的钻蛀型害虫,主要为害阔叶树根及树干。在我国,木蠹蛾的发生面积广,单株虫口密度大(100~500余头),发生面积从百公顷到千公顷以上不等[6-7]。其中,榆木蠹蛾Deserticossustsingtauana(Bang-Haas, 1912),主要为害白榆[8],在宁夏2年发生1代,主要为害枝干和根茎部,以幼虫在蛀道内越冬[9];沙蒿木蠹蛾Deserticossusartemisiae(ChouetHua, 1986)主要分布在我国宁夏、内蒙古和陕西等干旱荒漠草原区,在宁夏2年发生1代,以幼虫为害油蒿ArtemisiaordosicaKrasch.(黑沙蒿)和籽蒿A.sphaerocephalaKrasch.(白沙蒿)的主茎和根部,造成灌丛成片死亡,以各龄幼虫在蛀道里越冬[10];沙棘木蠹蛾Eogystiahippophaecolus(Hua, Chou, Fang & Chen, 1990),在辽宁建平约4年1代[11],主要以幼虫为害沙棘的主干和根部,初孵幼虫主要钻蛀树干的韧皮部,造成树木表皮干枯,极少数钻蛀木质部,幼虫于入冬前转移至地下钻蛀根部为害,树根大部分被蛀空,导致整株枯死,以幼虫在蛀道内越冬[12]。
目前,国内外关于木蠹蛾的研究报道大多集中于生物学、信息素、发生预测及防治方面,至今未见有关其耐寒研究的报道。本文以宁夏地区3种发生较为严重的木蠹蛾越冬幼虫为研究对象,通过对比3种木蠹蛾的过冷却能力及不同时期、不同发育阶段过冷却能力变化,明确该地区主要鳞翅目钻蛀类害虫对低温环境的适应差异。为进一步研究该地区钻蛀性害虫的耐寒策略、生理生化调节规律及种群建立可能性分析提供理论基础。
1 材料与方法
1.1 材料来源
分别于2013年3月和2014年1月,在宁夏彭阳和盐池采集榆木蠹蛾、沙蒿木蠹蛾和沙棘木蠹蛾越冬幼虫。采集到的越冬幼虫置于寄主根部的土中保存后在24 h内带回实验室不加任何处理,选择生长状态良好的健康幼虫进行各项指标的测定。
1.2 测定方法
1.2.1 沙棘木蠹蛾虫龄测定
将幼虫于显微镜(LEICA EZ 4 D)下拍照,根据显微镜放大倍数在照片中测量其头壳宽度和体长,依照所测得的数据,对幼虫进行虫龄划分[13],根据越冬幼虫的虫龄分布情况,选取8~10龄、11~13龄和14~16龄3个虫龄阶段进行相关耐寒性指标测定。
1.2.2 过冷却点及结冰点测定
每种木蠹蛾取10头以上幼虫作过冷却点及结冰点测定。通过过冷却点测定仪+低温冰箱连接计算机测量系统对存活的木蠹蛾幼虫进行过冷却点的测定:首先,用封口膜包裹不同木蠹蛾幼虫虫体将其固定在热敏电阻探头上,另一端与四路昆虫过冷却点测定仪(江苏森意经济发展有限公司)相连;然后,将固定虫体的探头放入-40~150℃的高低温试验箱(北京雅士林试验设备有限公司)中设置恒温-30℃,并用保护材料(脱脂棉)包裹虫体,使虫体保持缓慢的速率(1℃/min)持续降温;最后,由昆虫过冷却点测试系统通过计算机软件记录木蠹蛾幼虫体表温度的变化,测出其过冷却点和结冰点。
1.2.3 数据处理方法
利用GraphPad Prism 6 统计软件中的One-way ANOVA 进行单因素方差分析,采用Tukey检验进行多重比较,利用t检验进行两个时期木蠹蛾过冷却点及结冰点差异比较。
2 结果与分析
2.1 不同种类木蠹蛾越冬幼虫过冷却点及结冰点
不同木蠹蛾越冬幼虫的过冷却点及结冰点如表1所示。榆木蠹蛾、沙蒿木蠹蛾和沙棘木蠹蛾的过冷却点基本相同,但沙蒿木蠹蛾的结冰点明显低于榆木蠹蛾(P=0.047 8)和沙棘木蠹蛾(P=0.000 8)。
表1三种木蠹蛾越冬幼虫过冷却点及结冰点(2013年3月)1)
Table1SCPsandFPsoftheoverwinteringlarvaeofthreeCossidaespeciesinMarch2013
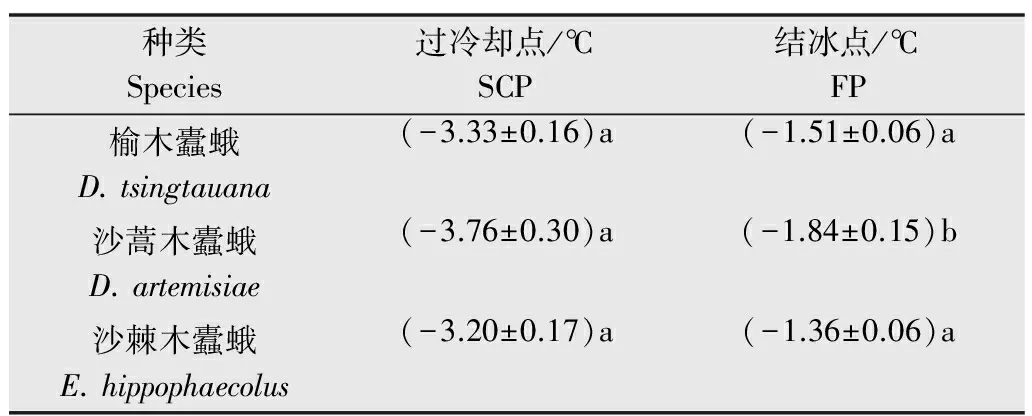
种类Species过冷却点/℃SCP结冰点/℃FP榆木蠹蛾D.tsingtauana(-3.33±0.16)a(-1.51±0.06)a沙蒿木蠹蛾D.artemisiae(-3.76±0.30)a(-1.84±0.15)b沙棘木蠹蛾E.hippophaecolus(-3.20±0.17)a(-1.36±0.06)a
1) 表中数据为平均值±SE,表2和表3相同。同列数据后不同小写字母表示有显著差异(P<0.05)。 Data in the table are the mean±SE; the same in table 2 & table 3. Different lowercase letters in the same column indicate significant difference among species (P<0.05).
3种木蠹蛾过冷却点的频次分布如图1所示,沙棘木蠹蛾和榆木蠹蛾的过冷却点变化范围基本相同,沙蒿木蠹蛾的过冷却点变化范围较其他2种整体降低1℃;2013年3种木蠹蛾越冬幼虫的冰点相对集中在-2℃与-1℃之间(沙蒿木蠹蛾有部分个体可以下降到-3℃)。
2.2 不同时期木蠹蛾越冬幼虫过冷却点及冰点
不同时期木蠹蛾越冬幼虫过冷却点及结冰点如表2所示。2013年3月榆木蠹蛾的过冷却点和结冰点明显高于2014年1月(P1<0.000 1,P2<0.000 1),沙蒿木蠹蛾的变化规律同上(P3=0.000 2,P4<0.000 1)。
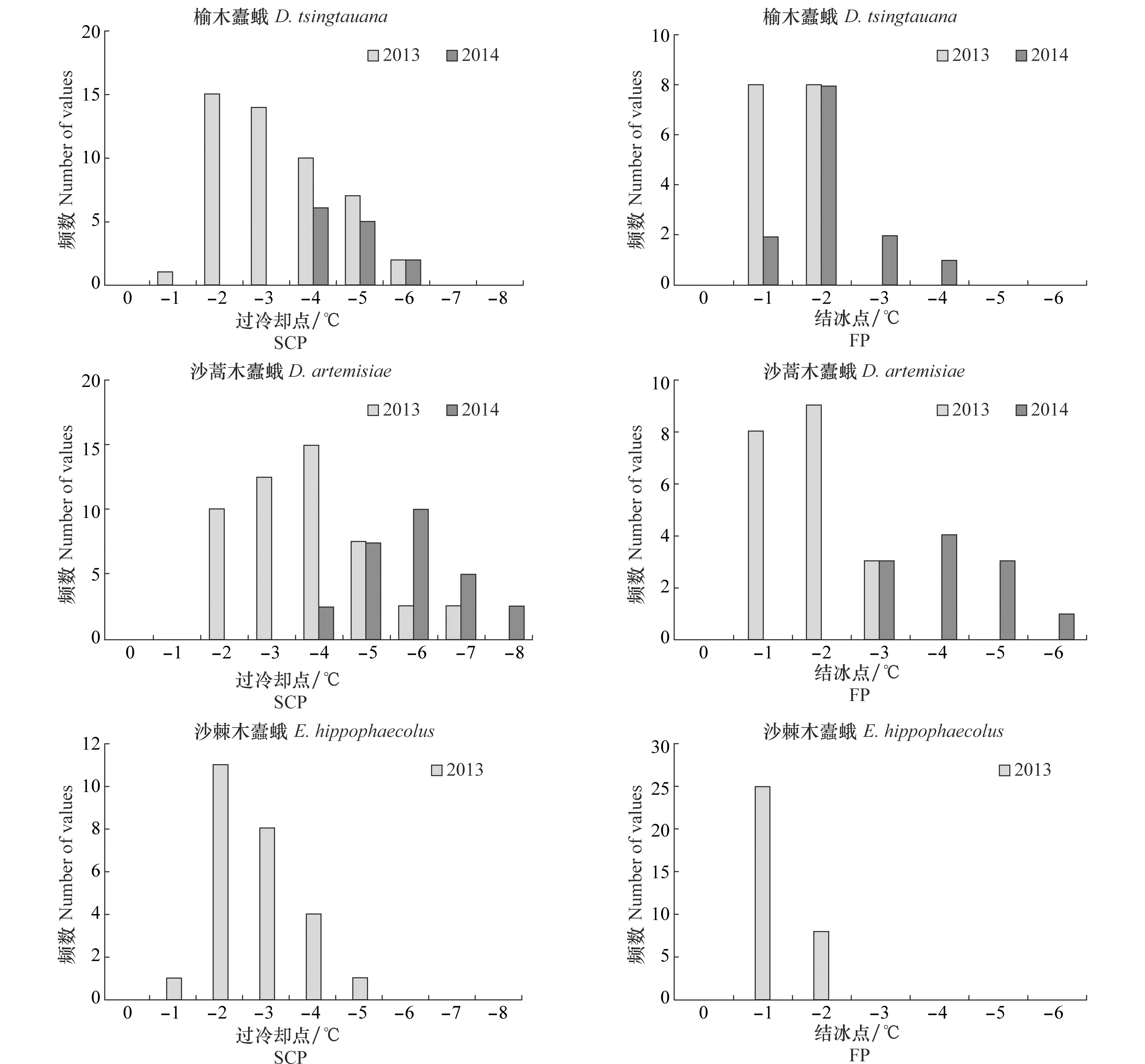
图1 2013年3月及2014年1月3种木蠹蛾越冬幼虫过冷却点和结冰点的频次分布Fig.1 Distribution of SCPs & FPs among the overwintering larvae of three Cossidae species in March 2013 & January 2014

表2 2种木蠹蛾越冬幼虫的过冷却点及结冰点1)
1)*表示同种木蠹蛾不同时期相同耐寒指标有显著差异(P<0.05)。*indicates significant difference in the same species during different periods (P<0.05).
比较两种木蠹蛾不同时期的过冷却点频次分布(图1),榆木蠹蛾2013年3月的过冷却点下限与2014年1月的一致,但后者更为集中,上限较前者低3℃,沙蒿木蠹蛾与榆木蠹蛾类似,2014年1月的过冷却点下限较2013年3月低1℃,上限低2℃。两种木蠹蛾不同时期的冰点频次分布如图1,2014年1月的过冷却点较2013年3月有整体向低温方向移动的明显趋势,同时2014年1月两种木蠹蛾的冰点频次分布的离散程度明显高于2013年3月。
2.3 不同虫龄阶段沙棘木蠹蛾越冬幼虫的过冷却点及结冰点
比较不同虫龄阶段越冬幼虫的过冷却点及结冰点(表3)可以发现,各虫龄阶段的过冷却点和结冰点均维持在相同的水平,虫龄间没有显著差异。从SCP(图2)和FP(图3)的频次分布结果来看,低龄组的变化范围明显比高龄组集中,8~10龄组的过冷却点变化范围在-4.5~-2.0℃之间,冰点变化范围在-1.8~-1.0℃之间,而14~16龄组的过冷却点最低可以探测到-6.0℃,最高可以探测到-1.5℃,冰点也可以探测到-3.4℃。

图2 沙棘木蠹蛾3个虫龄阶段越冬幼虫过冷却点的频次分布Fig.2 Distribution of SCPs among three instars of Eogystia hippophaecolus
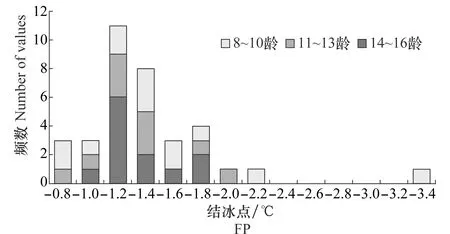
图3 沙棘木蠹蛾3个虫龄阶段越冬幼虫结冰点的频次分布Fig.3 Distribution of FPs among three instars of Eogystia hippophaecolus

表3 三个虫龄阶段沙棘木蠹蛾越冬幼虫的过冷却点及结冰点1)
1) 同列数据后相同字母表示木蠹蛾不同发育时期耐寒指标无显著差异(P>0.05)。 The same letters in the same column indicate no significant differences among different instars (P>0.05).
3 讨论
3.1 不同木蠹蛾越冬幼虫的过冷却能力差异
造成昆虫耐寒性差异的因素主要分为环境变化的影响(季节性气温变化、地理环境、个体发育阶段差异、低温驯化作用以及时间和温度的交互作用)及自身生理生化机制的作用(抗冻保护剂的产生及其作用、冰核剂及抗冻蛋白的作用等)[4]。即使在同一气候条件下,不同昆虫的过冷却能力也可能存在明显差异,例如栖息于红花尔基樟子松林内的红松实小卷蛾、曲姬蜂和广肩小蜂幼虫,在1988年10月过冷却点分别为-45℃以下、-32.51℃和-38.25℃,造成这种差异是因为三者越冬环境的差异[14]。本文中,3种木蠹蛾生活在相同的气候条件中,又同为钻蛀性害虫,越冬条件基本相似,因此从2013年3月份的数据来看,榆木蠹蛾、沙蒿木蠹蛾和沙棘木蠹蛾的过冷却能力基本相同,2014年1月份的数据再次佐证了榆木蠹蛾和沙蒿木蠹蛾过冷却能力的相似性。此外,榆木蠹蛾与沙棘木蠹蛾的过冷却点与结冰点频次分布范围基本相同,沙蒿木蠹蛾的两项指标较前两个物种有明显的向低温转移的趋势,说明沙蒿木蠹蛾在3种木蠹蛾中对低温环境的适应能力较强。
3.2 不同时期木蠹蛾越冬幼虫的过冷却能力差异
不同越冬时期的昆虫过冷却能力有明显差异,这主要是由于长期低温驯化所致。昆虫在越冬之前经历一个温度逐渐降低的过程,使他们能够为度过低温环境做一些生理上的准备[4]。不同时期二点委夜蛾的越冬幼虫过冷却点变化明显,11月份(越冬初期)-16.24℃,1月份(越冬中期)-23.16℃,3月份(越冬末期)恢复至-19.91℃[15]。桃小食心虫的越冬幼虫过冷却点也呈现出明显的季节性变化,越冬中期的过冷却点和冰点达到最低,显著低于其他时期,而越冬前期和后期没有明显差异[16]。本文中,榆木蠹蛾和沙蒿木蠹蛾在两个时期的过冷却点及结冰点的差异明显,说明2013年3月份的越冬末期的两种木蠹蛾幼虫在经历了一个夏天的解冻复苏和第二年的越冬前期驯化之后较之前具备了更强的过冷却能力。其中沙蒿木蠹蛾的变化幅度更为明显,这可能与两者越冬环境不同有关,榆树为木本植物,根茎组织发达,有更好的保温效果,因此对蛀道内越冬幼虫的长期驯化效果不及沙蒿。
一般来说,昆虫的耐寒类型分为两种,即耐结冰型和不耐结冰型。耐结冰型昆虫的过冷却点较高,可以在过冷却点以下生存[17]。而不耐结冰型的昆虫过冷却点通常比较低,昆虫不能在过冷却点以下温度存活[18-19]。红脂大小蠹越冬幼虫的过冷却点为-11.98℃,却能在-23.5℃条件下安全越冬[20]。本文中,2013年3月彭阳地区的月平均气温为7.3℃,2014年1月该地区的月平均气温为-3.3℃,其中最低气温为-17℃,两种木蠹蛾的过冷却点均值在2014年1月份均贴近月平均气温,而自2013年11月至2014年3月,有110天日最低气温低于-3℃,两种木蠹蛾在这么长时间低于过冷却点环境下仍能够顺利越冬,表现出了耐结冰型昆虫的一般特征,可以初步推测榆木蠹蛾和沙蒿木蠹蛾可能为耐结冰型昆虫。
3.3 不同虫龄阶段木蠹蛾越冬幼虫的过冷却能力差异
昆虫在不同生长发育阶段也会表现出不同的低温胁迫耐受性。一种摇蚊DiamesamendotaeMuttkowski在不同发育阶段中过冷却点差异明显,幼虫最高,成虫最低,其中不同发育阶段的幼虫过冷却点仍存在差异[21]。松墨天牛Monochamusalternatus卵的耐寒性显著高于其幼虫和蛹,而成虫则表现出了最弱的低温胁迫耐受性[22]。但在本文中,比较不同虫龄阶段沙棘木蠹蛾越冬幼虫的结冰点和过冷却点均没有明显差异,造成这种结果的原因可能是因为沙棘木蠹蛾4年1代[23],世代重叠现象普遍存在。这说明在同时期,不同虫龄的越冬幼虫均接受同样低温环境的胁迫,需要同样的低温耐受能力才能顺利完成越冬,所以单就同一种群而言,一个时期不同虫龄阶段沙棘木蠹蛾的越冬幼虫过冷却能力差异并不明显。由此推断另外3种也存在世代重叠现象的木蠹蛾越冬幼虫也有可能存在同样的现象。另外,虽然3个虫龄阶段的沙棘木蠹蛾越冬幼虫过冷却能力基本相当,但从过冷却点和冰点的频次分布可以看出,高龄组较低龄组的过冷却点和冰点范围更宽泛,且两个指标的极值均出现在高龄组也说明老龄幼虫对于温度变化的适应能力较低龄幼虫更强。
[1] 景晓红, 康乐. 昆虫耐寒性的测定与评价方法[J]. 应用昆虫学报, 2004, 41(1):7-10.
[2] Wharton D A, Aalders O, Bale J S, et al. Desiccation stress and recovery in the anhydrobiotic nematodeDitylenchusdipsaci(Nematoda: Anguinidae)[J]. European Journal of Entomology, 1999, 96(2):199-203.
[3] Sømme L. The history of cold hardiness research in terrestrial arthropods [J]. Cryo Letters, 2000, 21(5):289-296.
[4] 景晓红, 康乐. 昆虫耐寒性研究[J]. 生态学报, 2002, 22(12):2202-2207.
[5] Bale J S.Insect cold hardiness: A matter of life and death [J]. European Journal of Entomology, 1996, 93(3):369-382.
[6] 方德齐, 陈树良. 木蠹蛾种类及其分布的初步考察[J]. 山东林业科技, 1984(4):44-46.
[7] 方德齐, 陈树良, 李宪臣. 中国木蠹蛾研究进展情况[J]. 陕西林业科技, 1992(2):29-35.
[8] 萧刚柔. 中国森林昆虫[M]. 北京:中国林业出版社, 1992:765.
[9] 杨美红, 牛辉林, 张金桐,等. 榆木蠹蛾生物学特性观察[J]. 应用昆虫学报, 2012, 49(3):735-741.
[10] 王建伟, 骆有庆, 宗世祥. 沙蒿木蠹蛾生物学特性研究[J]. 昆虫学报, 2011, 54(7):809-814.
[11] 宗世祥, 骆有庆, 许志春,等. 沙棘木蠹蛾幼虫龄期的初步研究[J]. 应用昆虫学报, 2006, 43(5):626-631.
[12] 宗世祥. 沙棘木蠹蛾生物生态学特性的研究[D]. 北京:北京林业大学, 2006.
[13] 宗世祥, 骆有庆, 许志春,等. 沙棘木蠹蛾幼虫龄期的初步研究[J]. 应用昆虫学报, 2006, 43(5):626-631.
[14] 王志英, 岳书奎, 张国财. 几种森林昆虫耐寒性的研究[C]∥青年生态学者论丛.北京:中国科学技术出版社, 1991.
[15] 刘玉娟, 张天涛, 白树雄,等. 越冬期不同阶段二点委夜蛾越冬幼虫耐寒性变化[J]. 昆虫学报, 2014, 57(3):379-384.
[16] 王鹏, 凌飞, 于毅,等. 桃小食心虫越冬幼虫过冷却能力及体内生化物质动态[J]. 生态学报, 2011, 31(3):638-645.
[17] 王锦林, 冯宇倩, 宗世祥. 昆虫耐寒性的类型、测定方法及影响因素综述[J]. 安徽农学通报, 2013(9):32-35.
[18] Denlinger D L. Relationship between cold hardiness and diapause [M]. Insects at Low Temperature.Springer US, 1991:174-198.
[19] Chippendale R E. Insects at low temperature[J]. American Entomologist, 1991, 37(2):119-121.
[20] 赵建兴, 杨忠岐, Jean-Claude Gregoire. 红脂大小蠹Dendroctonusvalens(Coleoptera,Scolytidae)和大唼蜡甲Rhizophagusgrandis(Coleoptera,Rhizophagidae)的耐寒性[J]. 环境昆虫学报, 2009, 31(1):20-28.
[21] Bouchard R W, Carrillo M A, Kells S A, et al. Freeze tolerance in larvae of the winter-activeDiamesamendotae, Muttkowski (Diptera: Chironomidae): a contrast to adult strategy for survival at low temperatures [J]. Hydrobiologia, 2006, 568(1):403-416.
[22] Ma Ruiyan, Hao Shugang, Kong Weina, et al. Cold hardiness as a factor for assessing the potential distribution of the Japanese pine sawyerMonochamusalternates(Coleoptera:Cerambycidae) in China [J]. Annals of Forest Science,2006,63:449-456.
[23] 路常宽, 骆有庆, 许志春,等. 沙棘木蠹蛾的人工饲养[J]. 应用昆虫学报, 2005, 42(2):211-214.
(责任编辑: 田 喆)
ComparisonofthesupercoolingcapacityinthreespeciesofCossidaeoverwinteringlarvae
Tian Bin1, Xu Lili1, Wang Tao2, Zong Shixiang1
(1.KeyLaboratoryofBeijingfortheControlofForestPests,BeijingForestryUniversity,Beijing100083,China;2.MentougouForestryStation,Beijing102300,China)
The supercooling points (SCPs) and freezing points (FPs) of the overwintering larvae of three species of Cossidae (Deserticossustsingtauana,D.artemisiaeandEogystiahippophaecolus) were analyzed in different time points in Ningxia Hui Autonomous Regions, and the SCPs and FPs ofE.hippophaecolusin different instars were also detected. The results showed that the supercooling capacity of the three Cossidae larvae were basically the same.D.artemisiaehad the best adaptation to low temperature. Cold acclimation had a significant effect on the supercooling capacity; the SCPs and FPs ofD.tsingtauanaandD.artemisiaein March 2013 were higher than those in January 2014, and both Cossidae species showed the characteristics of “freezing-tolerance”. No difference in supercooling capacity were found among the three instars ofE.hippophaecolus, but the older larvae showed better adaptation to temperature variation.
Cossidae; overwintering larva; supercooling point; freezing point
S 763.3
: ADOI: 10.3969/j.issn.0529-1542.2017.05.018
2016-09-18
: 2017-02-03
国家自然科学基金(31470651)
* 通信作者 E-mail: zongsx@126.com
猜你喜欢
今日农业(2022年13期)2022-09-15
婚姻与家庭·性情读本(2022年5期)2022-05-16
中国水土保持(2022年3期)2022-03-24
山西林业(2021年2期)2021-07-21
青春期健康(2020年2期)2020-12-05
恋爱婚姻家庭·养生版(2020年6期)2020-07-09
恋爱婚姻家庭(2020年18期)2020-07-08
现代农村科技(2018年6期)2018-01-18
江苏农业科学(2016年7期)2016-10-20
