
土壤中盾壳霉双标记平板计数法的建立及应用
2017-10-09 00:52杨秀超俞文静段作营夏森玉沈志松李华钟
植物保护 2017年5期
杨秀超, 俞文静, 段作营, 夏森玉, 沈志松, 金 坚, 李华钟*
(1. 江南大学工业微生物技术教育部重点实验室, 无锡 214122; 2. 无锡楗农生物科技有限公司,无锡 214122; 3. 江南大学药学院, 无锡 214122)
土壤中盾壳霉双标记平板计数法的建立及应用
杨秀超1, 俞文静1, 段作营1, 夏森玉2, 沈志松2, 金 坚3, 李华钟1*
(1. 江南大学工业微生物技术教育部重点实验室, 无锡 214122; 2. 无锡楗农生物科技有限公司,无锡 214122; 3. 江南大学药学院, 无锡 214122)
盾壳霉是一种重要的生防菌,其在土壤中的存活数量直接关系到防治病害的效果。然而目前没有对土壤中盾壳霉直接计数的方法,构建一种简单易行的土壤中盾壳霉计数方法对研究盾壳霉在土壤中的存活动态具有重要意义。本研究利用农杆菌转化法构建了潮霉素基因和绿色荧光蛋白基因标记的双标记盾壳霉菌株,并测定转化子的生长速度、产孢量和菌核致腐能力,初步分析了该方法计数土壤盾壳霉的有效性和可行性。结果显示,潮霉素基因和绿色荧光蛋白基因可以稳定地遗传和表达,并且部分转化子生长速度、产孢量和菌核致腐能力与出发盾壳霉菌株JN-CM没有显著差异。加入土壤中的盾壳霉转化子可以在含潮霉素(50 μg/mL)、氯霉素(100 μg/mL)和链霉素(100 μg/mL)的PDA平板培养,杂菌得到充分抑制,呈现绿色荧光的盾壳霉转化子被有效检出,检出限达到2×103个/g土。本研究所构建的计数方法能有效检出施入土壤中的盾壳霉并进行活菌计数,可以用于盾壳霉JN-CM产品在土壤中的定殖、生长、繁殖和存活情况的研究。应用双标记平板计数法研究了不同温度、湿度、接种量和添加菌核等条件下盾壳霉JN-CM在土壤中的存活规律。结果显示,在含有核盘菌菌核的土壤中,盾壳霉JN-CM可以通过重寄生维持一段时间(12周)的数量增长,在长达半年左右(24周)的时间里其存活率仍然可以维持在65%左右。在不含菌核的土壤中,在一般土壤温度(10~20℃)范围内,无论土壤水分含量高低,其半年存活率也可以维持在50%左右。因此,可以预测,连续施用盾壳霉JN-CM生防制剂,可以使其数量在土壤中长期维持在一定的水平范围,达到长效防治效果。
生物防治; 盾壳霉; 农杆菌介导转化; 平板计数
核盘菌Sclerotiniasclerotiorum(Lib.)de Bary是一种世界性分布的植物病原真菌,能侵染75科450种植物,以草本植物居多[1]。在我国,核盘菌的寄主植物也高达36科214种,核盘菌引起的菌核病给我国粮油作物和蔬菜作物造成了严重的危害[2]。目前,我国主要采用化学杀菌剂来控制菌核病,但化学杀菌剂引起的环境污染、抗性菌株产生和农药残留等问题越来越严重,寻找安全、环保、高效的防治方法成为了必然选择。盾壳霉Coniothyriumminitans是核盘菌的重寄生菌,被认为是防治菌核病最有效的生防菌之一[3-4]。国内外许多研究表明在土壤中施用盾壳霉培养物可以有效地控制核盘菌菌核的存活及子囊盘萌发,从而控制核盘菌的初侵染源数量,目前已有多种商品化盾壳霉产品用于防治大田作物、温室及园艺作物菌核病[5-10]。
本实验室筛选获得一株盾壳霉菌株,并命名为JN-CM,通过与无锡楗农生物科技有限公司合作已完成了JN-CM盾壳霉产品的生产工艺开发,并取得盾壳霉新农药正式登记证,进入生产和推广阶段。推广中农民普遍关心药效持续时间和施药次数等问题。而盾壳霉防效与其在土壤中存活的数量是直接相关的[11-12]。因此,有必要了解盾壳霉施入土壤后的定殖、生长、繁殖和存活情况。由于施入土壤中盾壳霉的数量有限,且土壤中存在大量杂菌,因此不能采用普通的平板菌落计数法对盾壳霉进行直接计数,必须构建抗性菌株采用选择性平板或者PCR扩增的方法进行检测[13-14]。为了提高检出的灵敏度和特异性,本研究构建了含有潮霉素抗性选择标记和绿色荧光蛋白鉴别标记的双标记盾壳霉重组菌株,其生长速度、产孢量和致腐能力与出发菌株JN-CM没有显著差异,可以用来代替JN-CM盾壳霉产品进行土壤定殖、生长、繁殖和存活情况研究。
1 材料与方法
1.1 材料
供试菌株和质粒:大肠杆菌DH5α购自天根生化科技有限公司;农杆菌EHA105感受态细胞购自上海士峰生物科技有限公司;盾壳霉菌株JN-CM由本实验室分离保存;双元载体pTFCM由华中农业大学姜道宏教授惠赠;含有红绿荧光基因的载体pCI-eGFP-DsRed和pET-20b(+)由本实验室保存。
试剂:T4 DNA连接酶、限制性内切酶XbaⅠ、HindⅢ、ClaⅠ、BamHⅠ、KpnⅠ、SacⅠ、EcoRⅠ和DNA聚合酶等购自TaKaRa公司(中国);质粒提取试剂盒、胶回收试剂盒、PCR产物回收试剂盒、乙酰丁香酮、硫酸链霉素、氯霉素、卡那霉素等购自上海生工生物公司;2-(N-吗啉)乙磺酸钠 (MES)购自上海麦克林生化科技有限公司;潮霉素购自北京索莱宝科技有限公司;其余均为国产或进口分析纯试剂。
1.2 PCR扩增引物
本文中进行PCR扩增的引物见表1,均由上海生工生物工程股份有限公司合成。用下划线标出了限制性内切酶位点,各引物对对应的限制性内切酶分别是:P1 和P2,XbaⅠ和λ-HindⅢ;P3和P4,ClaⅠ和BamHⅠ;P5和P6,SacⅠ和KpnⅠ。
表1质粒构建所用引物
Table1Primersusedforplasmidconstruction

引物Primers扩增片段Amplifiedfragment片段长度/bpLength序列(5′⁃3′)SequencesP1P2P3P4P5P6PtrpC⁃HYG⁃TtrpC2110eGFP738PtrpC⁃eGFP⁃TtrpC1794GCTCTAGAAAGAAGGATTACCTCTAAACACGCAAGCTTCGACAGAAGATGATATTGAAGGCCATCGATGCCTCGAGAATTCACCATCGGGATCCGATTACTTGTACAGCTCGTCCGAGCTCGAAAGAAGGATTACCTCTAAACACGGGGTACCCGACAGAAGATGATATTGAAGG
1.3 潮霉素抑制盾壳霉生长最低浓度测定
潮霉素是抗真菌类抗生素,但不同真菌对潮霉素敏感程度不同,因此在将潮霉素抗性基因导入盾壳霉之前需要确定盾壳霉对潮霉素的敏感程度。盾壳霉菌株JN-CM在PDA平板培养活化直至菌落长满整个平板,用打孔器打取直径为0.5 cm的琼脂块分别接种到潮霉素浓度为20、25、30、35、40、45、50和55 μg/mL的PDA平板中央,置于20℃培养,每个浓度设3个重复。十字交叉法测定5 d和9 d时菌落直径,计算抑制盾壳霉JN-CM生长的最低浓度。
1.4 双标记双元载体pTFCM-eGFP构建
以表1中所列引物进行PCR扩增,按照图1所示流程构建双标记双元载体。采用热激转化法将构建好的质粒转入DH5α感受态细胞,并用含50 μg/mL卡那霉素的LB平板进行筛选,挑取单菌落进行培养并提取质粒,进行PCR和单酶切电泳图谱鉴定,对鉴定正确的质粒进行测序验证。

图1 双标记双元载体构建流程图Fig.1 Flow chart of construction of double-labeling binary vector
1.5 农杆菌的转化
将-80℃保存的农杆菌感受态细胞取出于冰上融化,每100 μL感受态细胞加1 μg质粒DNA混匀,依次于冰上静置10 min、液氮5 min、37℃水浴5 min、冰上静置5 min,加入700 μL无抗生素的LB液体培养基,于28℃振荡培养2~3 h。取100 μL培养后的菌液涂布到含有卡那霉素的LB平板上,倒置放于28℃培养箱培养2~3 d。挑取单菌落,液体培养5~8 h,取1 μL菌液做模板,以P3、P4为引物进行PCR验证。
1.6 盾壳霉的共转化
农杆菌的培养参考Bundock等所描述方法[15]。农杆菌介导的盾壳霉转化试验参考李模孝等所描述方法,并进行部分改动[16-17]。挑取含有重组质粒的农杆菌到含50 μg/mL卡那霉素的LB培养基中,28℃,200 r/min振荡培养1 d,取100 μL农杆菌菌液加入到含有50 μg/mL卡那霉素的10 mL MM培养基中,200 r/min, 28℃培养2 d,测定菌液的OD600,用含有4 μmol/mL乙酰丁香酮(As)和1.0 g/L 2-(N-吗啉)乙磺酸钠 (MES)的诱导培养基(IM)稀释至OD600为0.15,在28℃,220 r/min振荡培养6 h备用。将盾壳霉JN-CM接种到PDA平板培养21 d,用无菌水洗下平板上孢子并离心收集分生孢子,用农杆菌细胞悬液稀释孢子数到108个/mL备用。制备含有4 μmol/mL As 和1.0 g/L MES共转化诱导培养基(CO-IM)的无菌平板,并在上面铺上一层硝酸纤维素膜,取制备好的农杆菌和盾壳霉孢子悬液200 μL涂布在平板上,25℃黑暗培养2 d,将硝酸纤维素膜揭起正面朝下盖在含有50 μg/mL潮霉素和500 μg/mL氨苄青霉素的PDA平板上。20℃培养5~8 d,长出来的菌落转移到含60 μg/mL潮霉素的PDA平板,能继续生长推定为转化子。随机选择几株可能的转化子进行培养,收集菌丝提取总DNA,以P3和P4为引物扩增绿色荧光基因片段,PCR产物经1%琼脂糖凝胶电泳分析鉴定。
1.7 转化子稳定性测定
随机选择5株转化子在不含抗生素的PDA平板培养10 d以上,用无菌水洗脱平板上孢子,并用2层擦镜纸过滤掉菌丝,得到纯孢子悬浮液。将孢子悬浮液涂布到无抗生素PDA平板,20℃培养10 d。如此重复直至第10代。测定第10代转化子中潮霉素抗性基因和绿色荧光基因表达情况。
1.8 转化子生长速度、产孢量和菌核致腐能力测定
菌丝生长速度:用打孔器打取直径0.5 cm的盾壳霉菌落边缘的琼脂块接种到不含抗生素的PDA平板中央,20℃培养,十字交叉法测定培养5 d和9 d的菌落直径,每菌株设置3个重复。比较出发菌株和转化菌株1~5的差异[18-20]。
孢子萌发率:将出发菌株和转化子在PDA平板培养至产生大量孢子,洗脱孢子并计数。稀释使孢子浓度为107个/mL,取100 μL涂布到无抗生素PDA平板,每株菌3个重复。分别培养36 h后置于显微镜下(200×)计数孢子萌发情况,每个平板随机检测5个视野(每镜头孢子数≥30),统计各平板孢子的萌发率[18-20]。
产孢量:用接种环刮取盾壳霉点接到PDA平板中央于20℃培养21 d,用直径0.5 cm打孔器随机打取5个琼脂块加入10 mL无菌水并充分研磨,用血球计数板计数。每株菌设3个重复[21]。
菌核致腐能力:将河沙洗净灭菌后干燥,称取20 g到无菌培养皿,加入大小均匀的菌核20粒,将出发菌株和5株转化子在PDA平板培养15 d,洗脱孢子并稀释孢子数到106个/mL,每个平板加4 mL稀释好的孢子悬浮液并搅拌混匀,20℃避光培养28 d,每株菌设3个重复[19]。菌核腐烂程度分级标准按姜道宏等的分级方法[22],参照方中达的病情指数计算公式计算菌核腐烂指数[23]。
1.9 施入土壤中盾壳霉转化子计数
将得到的转化子在PDA平板上培养,无菌水洗下平板表面孢子得到分生孢子悬浮液,将孢子悬浮液分别稀释到104、106、108个/mL,将同一批孢子悬浮液分别加入到灭菌土和未灭菌土中使孢子浓度达到104、106和108个/g。放置约4~6 h使孢子悬浮液充分被土壤吸收。取样后进行梯度稀释,用含潮霉素(50 μg/mL)、氯霉素(100 μg/mL)和链霉素(100 μg/mL)的PDA平板20℃培养6 d,对呈绿色荧光的菌落进行计数。同时,对孢子悬浮液直接稀释,在上述平板上进行计数,并比较不同试验组的结果。
1.10 不同条件土壤中盾壳霉存活情况
土壤中盾壳霉的存活与生长往往会受到温度、水分等条件的影响,同时菌核的存在对盾壳霉的保留和生长具有促进作用。因此,本研究对含水量11%、17%、23%,温度10、20、30℃,接种量2×104、2×105、2×106个/g,是否添加菌核等4个方面进行试验,每组3个重复,并用构建的双标记平板计数法进行计数。
1.11 数据处理
潮霉素抑制盾壳霉最低浓度,转化子和JN-CM菌株生长、繁殖、菌核致腐能力比较,盾壳霉选择性平板计数结果等数据采用SPSS 22软件进行处理,用Duncan氏新复极差法进行差异显著性检验。
2 结果与分析
2.1 潮霉素抑制盾壳霉生长最低浓度
用0.5 cm直径的打孔器打取生长旺盛的菌落边缘琼脂块接种到含不同潮霉素浓度的PDA平板中央,通过十字交叉法测定菌丝生长速度。从表2中可以看出,随着潮霉素浓度增加,盾壳霉菌株JN-CM生长速度明显受到抑制,当潮霉素浓度达到40 μg/mL时,JN-CM完全不能生长,因此培养基中潮霉素浓度为40 μg/mL是抑制盾壳霉JN-CM生长的最低浓度。
表2潮霉素对盾壳霉JN-CM菌株在PDA平板生长的影响1)
Table2EffectofhygromycinonthegrowthofJN-CMstrainonPDAplate
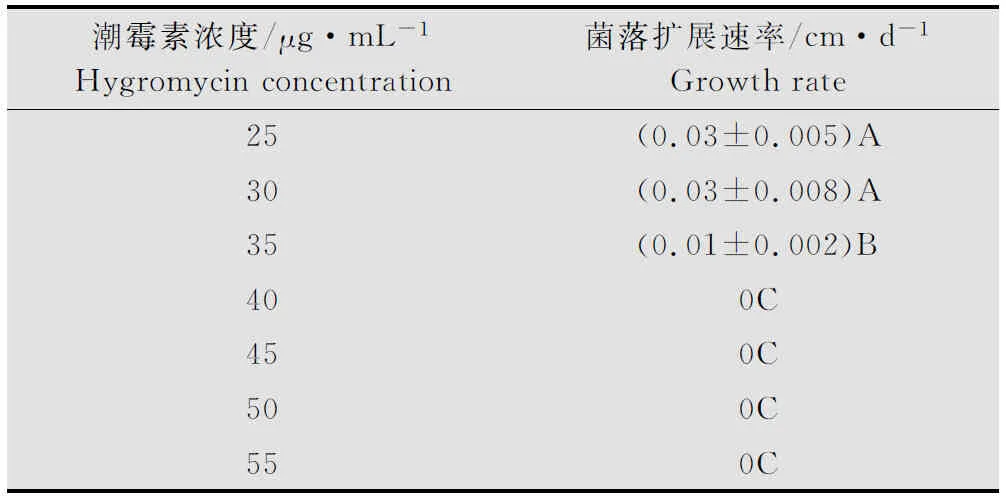
潮霉素浓度/μg·mL-1Hygromycinconcentration菌落扩展速率/cm·d-1Growthrate25(0.03±0.005)A30(0.03±0.008)A35(0.01±0.002)B400C450C500C550C
1) 不同大写字母表示差异极显著(P<0.01)。 The different capital letters indicated significant difference at 0.01 level.
2.2 双标记双元载体pTFCM-eGFP构建
平板上随机挑取8个单克隆,经菌液PCR扩增eGFP片段,8个克隆在738 bp附近都有阳性条带。选择其中6个提取质粒,用EcoRⅠ进行酶切鉴定,形成的3条大小为1 535、2 083、10 121 bp片段与图中特异性条带所在位置相符(图2)。测序结果与设计序列比对结果一致,表明双标记双元载体构建成功。
2.3 农杆菌的转化
在平板上随机挑取10个单克隆,经菌液PCR鉴定, 7个克隆在738 bp处有特异性条带,与扩增的eGFP片段大小相符。说明双元载体pTFCM-eGFP导入农杆菌成功。
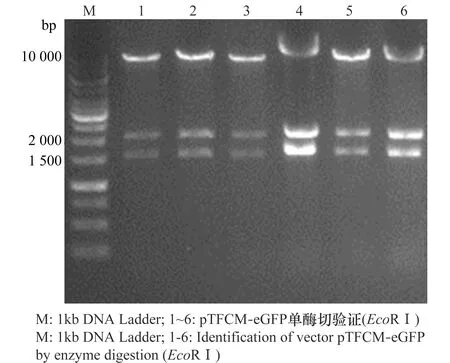
图2 双元载体pTFCM-eGFP酶切鉴定Fig.2 Identification of binary vector pTFCM-eGFP by enzyme digestion
2.4 盾壳霉转化子验证
随机选取5株可能的转化子,并编号为1~5。提取5株转化子总DNA进行PCR验证。扩增条带为2 110 bp的PtrpC-HYG-TtrpC片段和1 794 bp的PtrpC-eGFP-TtrpC片段,5株转化子电泳图特异性条带位置与理论值相符,出发菌株CK没有条带,说明两个片段完整插入盾壳霉中。
2.5 转化子稳定表达抗性基因和绿色荧光基因
转化子转代培养10代后仍能在含50 μg/mL潮霉素平板上生长,而出发菌株JN-CM不能生长(图3a)。同时将培养8 d的转化子制片用荧光显微镜观察,5株转化子都能观察到明显绿色荧光(图3c)。表明潮霉素基因和增强型绿色荧光基因在转化子内是稳定表达的,并具有良好的遗传稳定性。
2.6 转化子生长繁殖能力和对菌核致腐能力
比较转化子与出发菌株JN-CM生长速度、产孢量和对菌核致腐能力,结果见表3。从结果可以看出,5株转化子菌丝生长速度与出发菌株JN-CM没有显著差异;转化子孢子36 h萌发率与出发菌株JN-CM也无显著差别,且都超过了94%。这表明外源基因的插入未影响菌丝生长和孢子萌发。除转化子3的产孢量和菌核致腐能力显著低于其他转化子和出发菌株JN-CM,其他转化子产孢量和致腐能力没有明显变化,这可能是基因的插入破坏了产孢相关基因,而产孢量的下降导致了其对菌核寄生能力的降低。

图3 盾壳霉转化子抗性基因和荧光基因表达情况Fig.3 Expression of resistance gene and fluorescent gene in transformants of Coniothyrium minitans
由于除转化子3外,其他转化子的孢子萌发率、菌丝生长速度、产孢量、致腐能力等与出发菌株JN-CM均无显著变化,可以代替盾壳霉JN-CM产品进行土壤定殖、存活和生长、繁殖研究,因此,随机选取转化子1进行后续试验。
表3盾壳霉转化子及JN-CM生长速度、产孢量和菌核致腐能力比较1)
Table3Growthrate,conidiaproductionandsclerotiaparasiticabilityofConiothyriumminitanstransformantsandstrainJN-CM
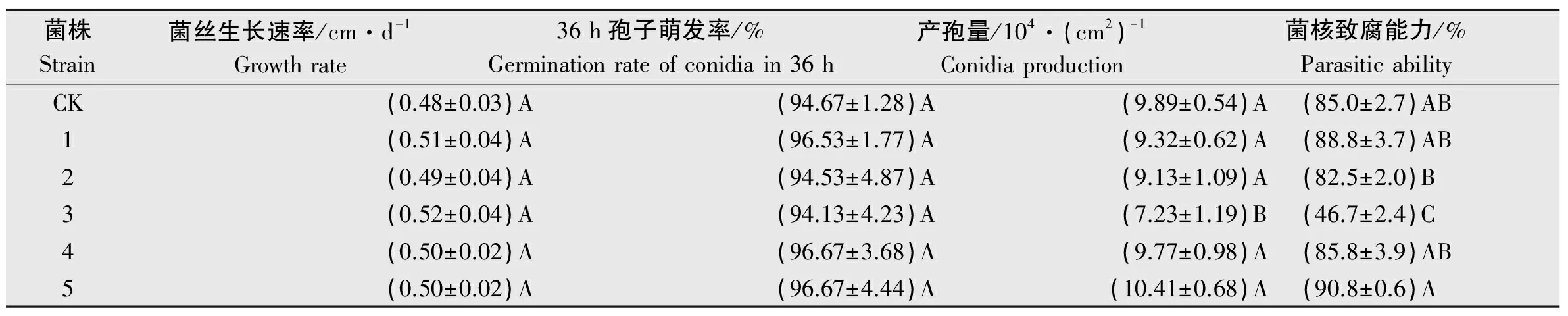
菌株Strain菌丝生长速率/cm·d-1Growthrate36h孢子萌发率/%Germinationrateofconidiain36h产孢量/104·(cm2)-1Conidiaproduction菌核致腐能力/%ParasiticabilityCK(0.48±0.03)A(94.67±1.28)A(9.89±0.54)A(85.0±2.7)AB1(0.51±0.04)A(96.53±1.77)A(9.32±0.62)A(88.8±3.7)AB2(0.49±0.04)A(94.53±4.87)A(9.13±1.09)A(82.5±2.0)B3(0.52±0.04)A(94.13±4.23)A(7.23±1.19)B(46.7±2.4)C4(0.50±0.02)A(96.67±3.68)A(9.77±0.98)A(85.8±3.9)AB5(0.50±0.02)A(96.67±4.44)A(10.41±0.68)A(90.8±0.6)A
1) 不同大写字母表示差异极显著(P<0.01)。 The different capital letters indicated significant difference at 0.01 level.
2.7 施入盾壳霉转化子选择平板计数结果
选择两个不同的地点挖取土样,去掉表面杂草和碎石后取表层10 cm的土壤,将土壤自然风干研磨过筛。取两种不同土样各5 g分别加入50 mL无菌水,稀释后涂布到抗性平板。链霉素100 μg/mL 和氯霉素100 μg/mL可以完全抑制土壤细菌在PDA平板上的生长。50、100、150 μg/mL 3种浓度潮霉素对土壤真菌的抑制没有明显区别,都表现为少量个别种类的真菌可以生长。标记后的盾壳霉带有绿色荧光,可以和杂菌进行区分,因此盾壳霉可以用抗性平板检出,检出限达到2×103个/g。
为验证土壤微生物和土壤对平板计数有无明显影响,分别测定了盾壳霉孢子浓度为104、106和108个/mL的孢子悬浮液和孢子含量为104、106和108个/g灭菌土、未灭菌土在选择平板上生长情况,结果显示,同一浓度下灭菌土壤和未灭菌土壤平板计数结果与对孢子悬浮液直接稀释进行平板计数处于同一水平,且不同浓度和不同稀释度下平板计数结果也无明显差异(图4)。表明土壤微生物和土壤对平板计数无明显影响,施入土壤中的盾壳霉可以用所建立的双标记平板法进行计数。

图4 不同样品中盾壳霉转化子在抗性平板上的萌发情况Fig.4 Germination of Coniothyrium minitans transformants in different samples on the resistant plate
2.8 不同条件土壤中盾壳霉存活情况
2.8.1 温度对盾壳霉在土壤中存活的影响
温度是影响盾壳霉菌丝生长和分生孢子萌发的重要因素。将含水量为17%,盾壳霉孢子浓度为2.0×105个/g的土壤分别置于10、20、30℃培养24周,考察温度对土壤中盾壳霉数量变化的影响,平板菌落计数法测定结果如图5所示。10℃和20℃条件下盾壳霉存活曲线下降趋势非常相似,在24周时仍能维持50%左右的存活率;30℃条件下,土壤中盾壳霉的存活率下降较快,施入后第3周存活率便降为20%以下,在24周时存活率仅有5%。说明高温不利于盾壳霉在土壤中的存活,这可能是持续的高温对孢子造成了损伤甚至致其死亡。当温度较低时,这种影响明显降低,存活时间也变得更长。
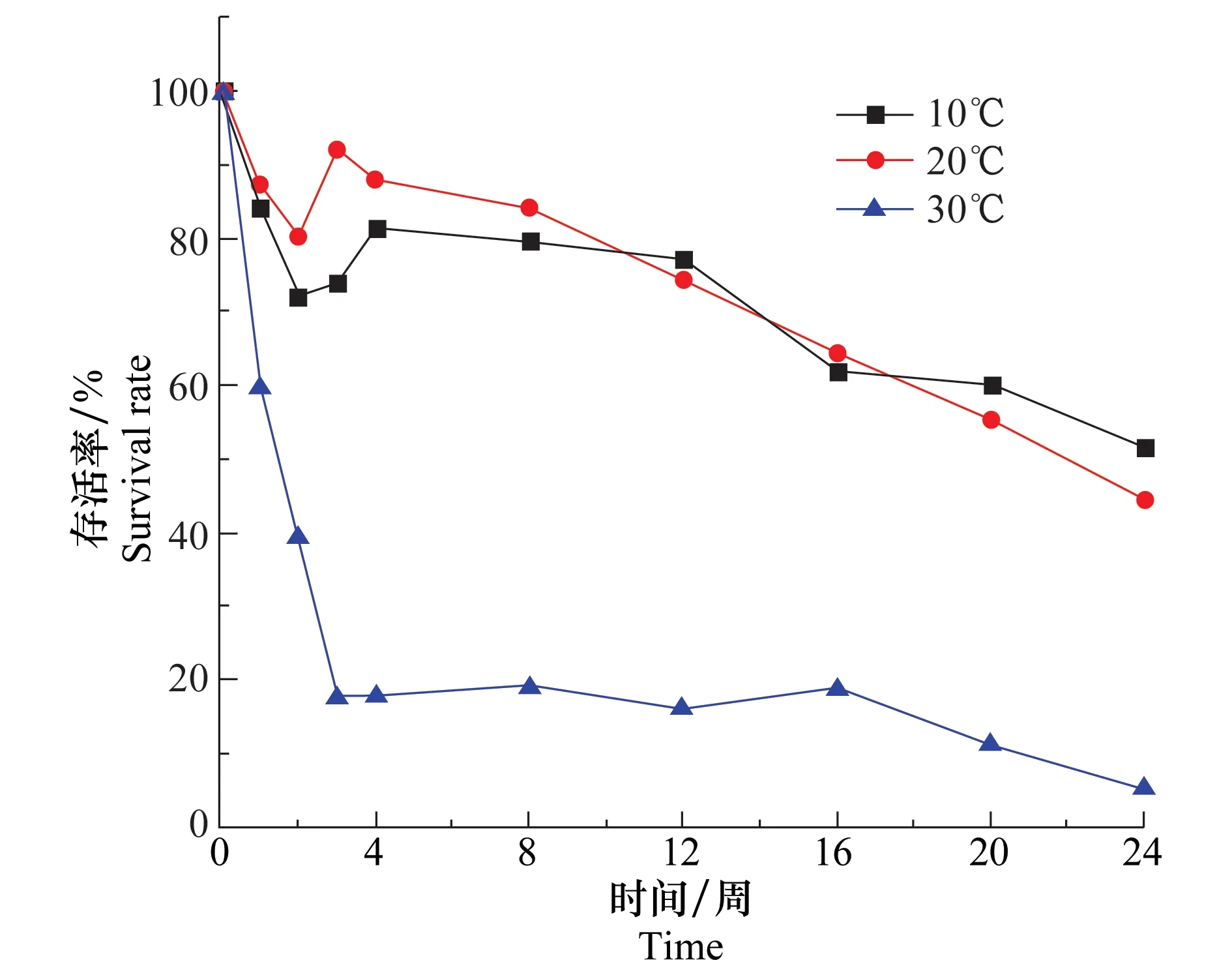
图5 温度对土壤中盾壳霉存活的影响Fig.5 Effect of temperature on the survival of Coniothyrium minitans in soil
2.8.2 土壤水分含量对盾壳霉存活的影响
水分会影响盾壳霉分生孢子的萌发和菌丝的生长,并且寄生菌核时,需要较高的水分含量。因此,土壤水分含量是影响盾壳霉存活的重要因素。在20℃,土壤中盾壳霉孢子含量为2.0×105个/g时测定3种含水量(11%、17%、23%)条件下盾壳霉在土壤中存活情况,结果如图6所示。由图中曲线可知,3种不同含水量条件下盾壳霉存活率下降趋势相同,在24周时,3组试验盾壳霉存活率均能保持在45%以上,表明土壤水分含量不会显著影响盾壳霉孢子在土壤中的存活。
2.8.3 接种量对盾壳霉数量变化的影响
在大田试验中,不同的报道所使用盾壳霉的浓度不尽相同。在20℃,土壤含水量为17%条件下,孢子浓度为2.0×104、2.0×105、2.0×106个/g的盾壳霉在土壤中存活情况如图7所示。虽然3组试验存活率曲线没有重合,但3条曲线下降的趋势差别不大,24周时,3个接种浓度存活率都在40%左右,这说明施入土壤的盾壳霉的量对其在土壤中的存活规律影响不显著。
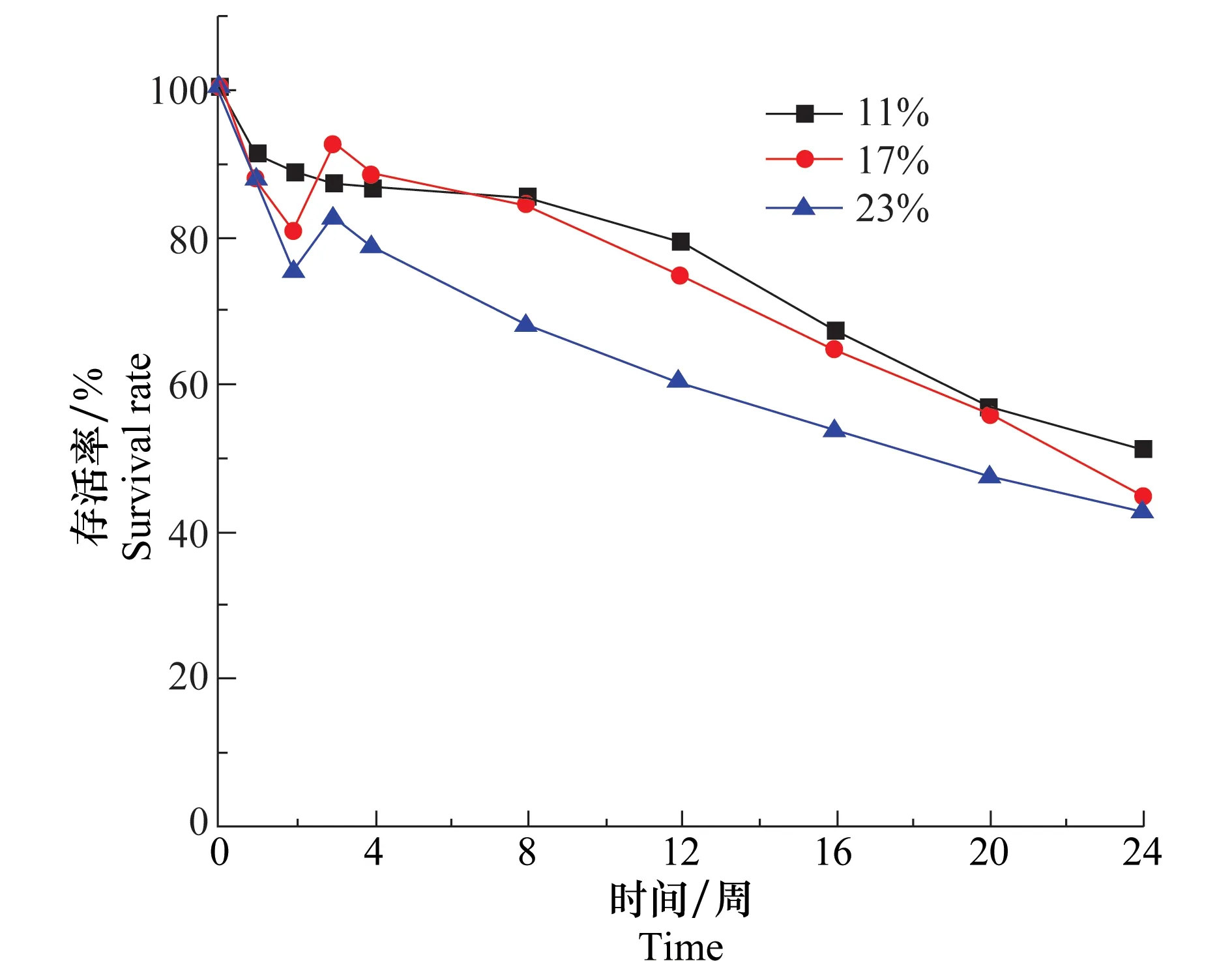
图6 土壤水分含量对盾壳霉存活的影响Fig.6 Effect of soil humidity on the survival of Coniothyrium minitans in soil
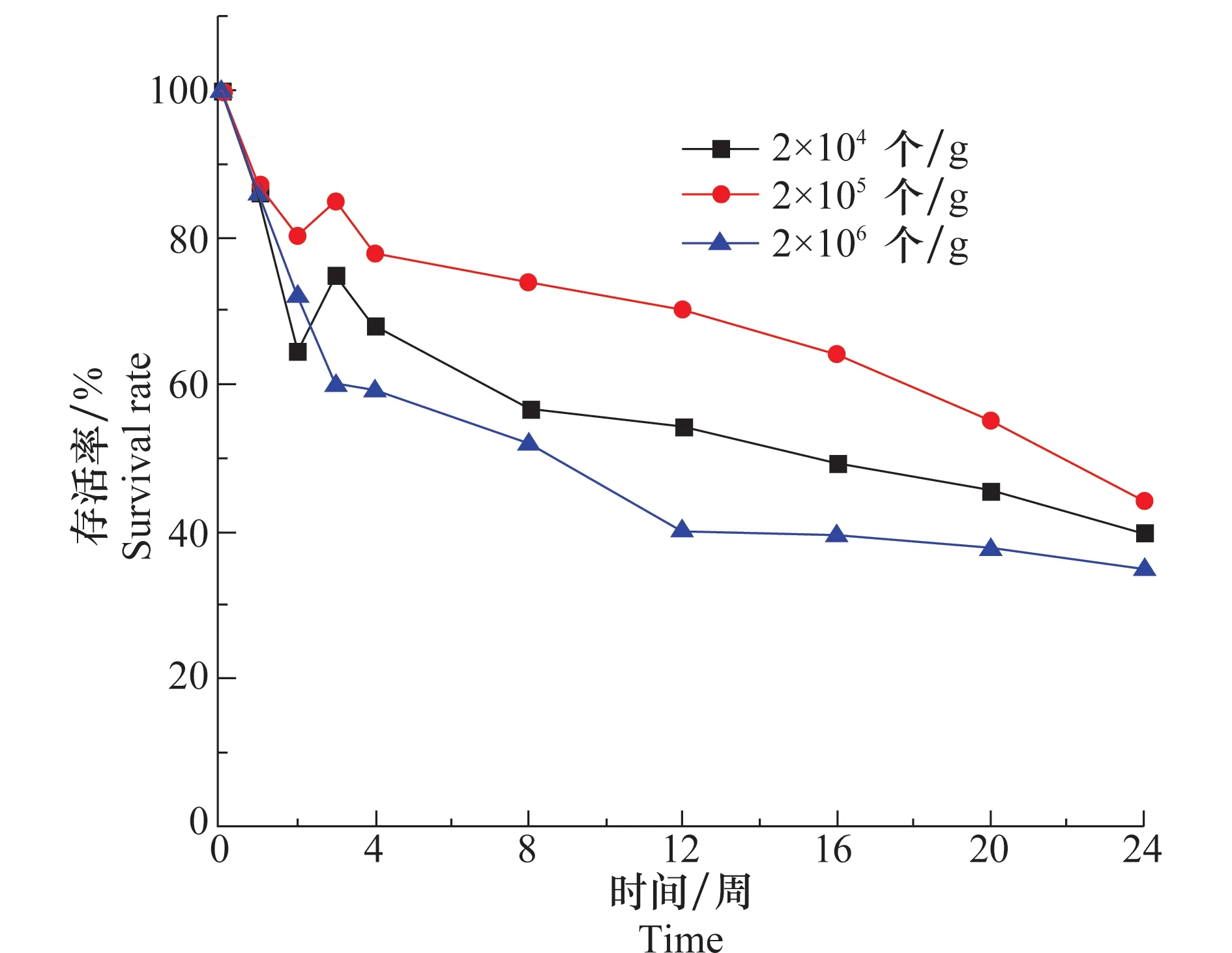
图7 不同接种量时盾壳霉在土壤中存活情况Fig.7 Survival of Coniothyrium minitans in soil at different inoculation amount
2.8.4 添加菌核对盾壳霉存活的影响
现有研究普遍认为,菌核的存在有利于盾壳霉在土壤中的保留,并且可以刺激盾壳霉数量的增加。将一定量的菌核加入含水量为17%,盾壳霉孢子浓度为2.0×105个/g的土壤中,置于20℃培养24周,测定不同时间盾壳霉数量,并与不加菌核土壤组进行对比,确定在土壤含有菌核时盾壳霉数量变化规律,结果如图8所示。从图中可知,未添加菌核组盾壳霉数量持续下降,而添加菌核组在施入盾壳霉第3周开始出现明显的上升趋势并一直持续到了12周,并在12周开始呈现下降趋势,但到第24周其存活率还可以维持在65%左右。这可能是由于在不含有菌核或菌核数量非常少时,土壤中的微生物对盾壳霉形成竞争和拮抗,从而导致盾壳霉数量持续下降,而在土壤含有菌核时,菌核会对盾壳霉产生刺激并引起盾壳霉对菌核进行寄生,以菌核为营养进行繁殖,从而引起盾壳霉数量的快速上升,当寄生结束时,一些菌体可能快速死亡,从而表现出盾壳霉数量的快速下降。
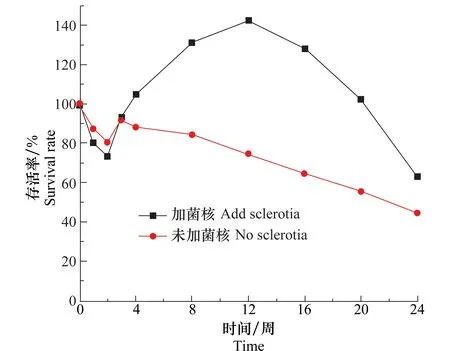
图8 土壤添加菌核对盾壳霉存活的影响Fig.8 Effects of adding sclerotia in soil on the survival of Coniothyrium minitans
3 结论与讨论
本研究成功构建了潮霉素筛选标记和绿色荧光鉴别标记的双标记双元载体,并用农杆菌转化法成功导入盾壳霉菌株,转化子中潮霉素抗性基因和绿色荧光基因能够稳定地遗传和表达。转化子36 h萌发率与对照组JN-CM处于同一水平,萌发率均在94%以上。除了转化子3以外,其他转化子产孢量和对菌核致腐能力没有明显改变,说明可以用它们代替JN-CM用于土壤定殖、存活和生长、繁殖研究。
将含有转化子孢子的灭菌土和未灭菌土壤稀释液以及孢子悬浮液涂布到含潮霉素的PDA平板上,在各供试浓度下同浓度不同样品的计数结果无明显差异,表明土壤微生物和土壤对盾壳霉计数没有明显影响。Bennett等[24]用灭菌土壤、巴氏消毒的土壤、未灭菌土壤研究盾壳霉在土壤中存活情况,使用含金霉素(25 mg/mL)和 Triton X-100 (2 mL/L)的PDA平板进行检测,在施入盾壳霉浓度为106个/g时,3种土壤中盾壳霉的数量变化情况区别不大,但在施入103个/g时,未灭菌土壤一组由于存在过多的其他真菌生长,只有少数盾壳霉能被检测到,而本研究构建的方法在盾壳霉孢子浓度较低时(2×103个/g),只有个别种类的少数杂菌长出,对计数没有明显影响,因此可以将本研究构建的方法应用到盾壳霉在自然土壤中存活情况的研究中,这将为盾壳霉在田间的存活动态提供重要参考。杨龙[25]通过适应性诱变获得抗农利灵盾壳霉突变菌株,建立了盾壳霉的抗性平板检测方法,并对盾壳霉在土壤中存活进行了研究,然而这种方法是在抗药突变机理尚不明确的情况下建立的,抗性是在压力环境下的一种适应还是产生了抗药基因也未知,而本研究构建的方法将潮霉素抗性基因插入盾壳霉基因组,抗药原理明确,遗传表达稳定。同时本研究构建的检测方法能够排除自然土壤中存在的野生盾壳霉菌株的干扰,可以用于施入土壤盾壳霉存活动态的研究。
通过对不同条件土壤中盾壳霉存活情况研究发现,在含有核盘菌菌核的土壤中,盾壳霉JN-CM可以通过重寄生维持一段时间(12周)的数量增长,在长达半年左右(24周)的时间里其存活率仍然可以维持在65%左右。即使在不含菌核的土壤中,在一般土壤温度(10~20℃)范围内,无论土壤水分含量高低,其半年存活率也可以维持在50%左右。虽然较高温度(30℃)条件下,其存活率快速下降,但除夏天的表层土外,大田土壤温度很少会持续维持30℃以上。因此,可以预测,连续施用盾壳霉JN-CM生防菌,可以使其数量在土壤中长期维持在一定的水平范围,达到长效防治效果。
[1] Boland G J, Hall R. Index of plant hosts ofSclerotiniasclerotiorum[J]. Canadian Journal of Plant Pathology, 1994, 16: 93-108.
[2] 杨新美. 油菜菌核病(Sclerotiniasclerotiorum)在我国的寄主范围及生态特性的调查研究[J]. 植物病理学报, 1959, 5(2): 111-122.
[3] Huang H C, Erickson R S.Factors affecting biological control ofSclerotiniasclerotiorumby fungal antagonists [J]. Journal of Phytopathology, 2008, 156(10): 628-634.
[4] Zeng Wenting, Kirk W W, Hao Jianjun. Field management ofSclerotiniastem rot of soybean using biological control agents [J]. Biological Control, 2012, 60(2): 141-147.
[5] De V T, Antoine N, Buitelaar R M, et al. The fungal biocontrol agentConiothyriumminitans: production by solid-state fermentation, application and marketing [J]. Applied Microbiology and Biotechnology, 2001, 56(1):58-68.
[6] Campbell W A.A new species ofConiothyriumparasitic on sclerotia [J]. Mycologia, 1947, 39(2): 190-195.
[7] Rabeendran N, Jones E E, Moot D J, et al. Biocontrol ofSclerotinialettuce drop byConiothyriumminitansandTrichodermahamatum[J]. Biological Control, 2006, 39(3): 352-362.
[8] Turner G J, Tribe H T. Preliminary field plot trials on biological control ofSclerotiniatrifoliorumbyConiothyriumminitans[J]. Plant Pathology, 1975, 24(2): 109-113.
[9] Jones E E, Mead A, Whipps J M. Evaluation of differentConiothyriumminitansinoculum sources and application rates on apothecial production and infection ofSclerotiniasclerotiorumsclerotia [J]. Soil Biology & Biochemistry, 2003, 35(3): 409-419.
[10] Yang Long, Li Guoqing, Zhang Jing, et al. Compatibility ofConiothyriumminitans, with compound fertilizer in suppression ofSclerotiniasclerotiorum[J]. Biological Control, 2011, 59(2):221-227.
[11] Gerlagh M, Goossen-van de Geijn H M, Hoogland A E, et al. Effect of volume and concentration of conidial suspensions ofConiothyriumminitanson infection ofSclerotiniasclerotiorumsclerotia [J]. Biocontrol Science and Technology, 2004, 14(7): 675-690.
[12] Jones E E, Clarkson J P, Mead A, et al. Effect of inoculum type and timing of application ofConiothyriumminitansonSclerotiniasclerotiorum: influence on apothecial production [J]. Plant Pathology, 2004, 53(5): 621-628.
[13] Jones E E, Stewart A, Whipps J M. Use ofConiothyriumminitanstransformed with the hygromycin B resistance gene to study survival and infection ofSclerotiniasclerotiorumsclerotia in soil [J]. Mycological Research, 2003, 107(3): 267-276.
[14] Jones E E, Stewart A.Coniothyriumminitanssurvival in soil and ability to infect sclerotia ofSclerotiniasclerotiorum[J]. New Zealand Plant Protection, 2011, 64: 168-174.
[15] Bundock P, Den D A, Beijersbergen A, et al. Trans-kingdom T-DNA transfer fromAgrobacteriumtumefacienstoSaccharomycescerevisiae[J].The EMBO Journal, 1995,14(13):3206-3214.
[16] Li Moxiao, Gong Xiaoyan, Zheng Jin, et al. Transformation ofConiothyriumminitans, a parasite ofSclerotiniasclerotiorum, withAgrobacteriumtumefaciens[J]. FEMS Microbiology Letters, 2005, 243(2): 323-329.
[17] 李模孝. 盾壳霉T-DNA标记插入突变体库的构建及其质量评估[D]. 武汉: 华中农业大学, 2013.
[18] 夏森玉, 雷楗勇, 金坚, 等. 生防菌盾壳霉抗药性突变菌株的选育[J]. 工业微生物, 2013, 43(5): 57-62.
[19] 夏森玉. 生防菌盾壳霉抗药性突变菌株的选育[D]. 无锡: 江南大学, 2013.
[20] 金强, 段作营, 夏森玉,等. 盾壳霉抗逆性菌株的筛选[J]. 工业微生物, 2016, 46(4):8-13.
[21] 魏玮. 盾壳霉产孢和生长相关基因的克隆及功能验证[D]. 武汉: 华中农业大学, 2013.
[22] 姜道宏, 李国庆, 易先宏, 等. 菌核寄生菌盾壳霉的研究Ⅱ.不同菌株培养特性及寄生致腐菌核能力的比较[J]. 华中农业大学学报, 1996(3): 229-232.
[23] 方中达. 植病研究方法[M]. 北京: 中国农业出版社, 1998.
[24] Bennett A J, Leifert C, Whipps J M. Survival of the biocontrol agentsConiothyriumminitans, andBacillussubtilis, MBI 600 introduced into pasteurised, sterilised and non-sterile soils [J]. Soil Biology & Biochemistry, 2003, 35(12):1565-1573.
[25] 杨龙. 生防菌盾壳霉防治油菜菌核病的生态学基础及其应用研究[D]. 武汉: 华中农业大学, 2009.
(责任编辑: 杨明丽)
Establishmentandapplicationofdouble-labelingplatecountingmethodfordetectionofConiothyriumminitansinsoil
Yang Xiuchao1, Yu Wenjing1, Duan Zuoying1, Xia Senyu2,Shen Zhisong2, Jin Jian3, Li Huazhong1
(1.KeyLaboratoryofIndustrialBiotechnology,MinistryofEducation,SchoolofBiotechnology,JiangnanUniversity,Wuxi214122,China; 2.WuxiJiannongBiologicalTechnologyCo.,Ltd,Wuxi214122,China;3.SchoolofPharmaceuticalSciences,JiangnanUniversity,Wuxi214122,China)
Coniothyriumminitansis an important biocontrol fungus. The quantity of the fungus surviving in the soil affects control efficacy directly. At present, there is still no direct way to count the quantity ofC.minitansin the soil, therefore, a simple and easy method for countingC.minitansin the soil will be very important. In this study, a double-labelingC.minitansstrain with hygromycin resistance gene and enhanced green fluorescent protein gene was constructed byAgrobacterium-mediated transformation. Subsequently, the stability, growth rate, reproduction capacity and sclerotium-parasitizing ability of the transformants were measured. The effectiveness and feasibility of the method were also analyzed. The results showed that the hygromycin resistance gene and enhanced green fluorescent gene could inherited and expressed stably, and there was no significant difference betweenC.minitansstrain JN-CM and the transformants in the ability of growth rate, reproduction and parasitizing on sclerotia ofSclerotiniasclerotiorum. PDA plates containing hygromycin (50 μg/mL), chloramphenicol (100 μg/mL) and streptomycin (100 μg/mL) could inhibit most of the microbes in the soil, while the transformants with green fluorescence could be detected and counted. The detection limit was 2 × 103spores per gram soil. Thus, the counting method can be used to study the colonization, growth, reproduction and survival ofC.minitansJN-CM in soil. The effects of temperature, soil humidity, inoculation amount, sterilization and adding of sclerotia on the survival ofC.minitansin soil were further investigated by the double-labeling plate counting method. The results showed that in the soil with sclerotia, the number ofC.minitansJN-CM increased in a period of time (12 weeks) and its survival rate can be maintained at about 65% in about half a year (24 weeks). While, in the soil without sclerotia, regardless of the soil moisture, the half-year survival rate can still be maintained at about 50% under the soil temperature of 10-20℃. Therefore, it can be predicted that continuous application ofC.minitansJN-CM biocontrol agent can keep the number ofC.minitansin the soil within a certain range for a long time, indicating that the long-term control effect can be achieved.
biological control;Coniothyriumminitans;Agrobacterium-mediated transformation; plate count
S 476
: ADOI: 10.3969/j.issn.0529-1542.2017.05.022
2017-01-19
: 2017-03-30
2015江苏省重点研发计划(现代农业)重点项目(BE2015309)
* 通信作者 E-mail: hzhli@jiangnan.edu.cn
猜你喜欢
中国生物防治学报(2022年4期)2022-09-16
四川蚕业(2022年1期)2022-06-06
西北农业学报(2021年5期)2021-06-28
中国食用菌(2021年1期)2021-03-12
中国食用菌(2019年9期)2019-10-21
爱你(2018年24期)2018-08-16
爱你·阳光少年(2018年8期)2018-05-14
大自然探索(2017年11期)2017-11-28
中国人兽共患病学报(2017年2期)2017-03-16
科学大众(中学)(2015年9期)2015-10-12
