
5—氨基乙酰丙酸(ALA)对秦岭高山杜鹃耐热性的诱导效应
2017-11-30 00:37李小玲华智锐张丹婷
江苏农业科学 2017年20期
李小玲+华智锐+张丹婷


摘要:在40 ℃高温胁迫下,测定秦岭高山杜鹃幼苗在不同浓度ALA(25、50、75、100、125 mg/L)处理后幼苗热害指数,叶片叶绿素、脯氨酸、丙二醛(MDA)含量,超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性。结果发现,ALA可以提高高温胁迫下秦岭高山杜鹃幼苗叶片叶绿素含量及SOD、POD、CAT活性,降低MDA含量,促进叶片内游离脯氨酸含量的积累,并在一定程度上降低热害指数。其中,50 mg/L ALA叶面喷施处理时对提高秦岭高山杜鹃耐热性相对效果最好。结果表明,叶面喷施ALA可以降低秦岭高山杜鹃的热害指数,缓解热损伤,提高植株的耐热性。
关键词:ALA;耐热性;高山杜鹃;生理特性
中图分类号: S685.210.1 文献标志码: A 文章编号:1002-1302(2017)20-0176-04
杜鹃属中陕西特有种较多,且具有单种蕴藏量大、抗寒性极强、观赏价值及药用价值高等特点,成为西北地区急需开发的野生资源之一[1]。而镇安木王国家森林公园的杜鹃花集中分布在双头马景区和茨沟杜鹃花景区,多为大叶美容杜鹃,在跑马梁深处和鹰嘴峰上分布的则为秀雅杜鹃[2]。近年来,由于全球气温不断升高,高温逆境对植物的生理生态过程造成了巨大的影响,而这对植物的耐热性也提出了更高的要求。杜鹃属植物主要分布在高海拔地区,生长繁殖要求冷凉的环境,对高温的耐受力不强,在其研究中温度已成为制约其生长的主要环境因子。因此,人们在对该属植物的引种栽培及育种研究中,耐热性的研究显得至关重要。
5-氨基乙酰丙酸(5-aminolevulinic acid,ALA),又称为δ-氨基酮戊酸,是生物体活细胞中的一种非蛋白氨基酸[3],是叶绿素、钴胺素、亚铁血红素等所有卟啉类化合物生物合成的前体物质[4]。大量研究表明,ALA除了作为叶绿素生物合成的重要中间产物外,还能够促进作物的光合作用[5-8],增加产量[9],提高甜瓜[5]、西瓜[10]、黄瓜及辣椒的抗冷性[11-12],参与植物抗逆性生理反应,具有调节光合色素的合成、提高光合效率、提高生物体内保护酶活性、保护细胞膜稳定性等功能[4-6],有类似于植物激素的调控效应[13]。目前,对于ALA的应用研究主要集中在抗冷性、抗盐性、抗旱性等方面,而在抗高温方面的研究则较少,有关ALA在高山杜鹃耐热性生理方面的研究未见报道。本研究以秦岭高山杜鹃为材料,研究在高温胁迫下ALA对植物生理状态的影响,旨在为杜鹃属植物的引种栽培、耐热育种和园林应用提供一定的理论参考。
1 材料与方法
1.1 试验材料及试剂
所用秦岭高山杜鹃幼苗采自镇安木王国家森林公园,ALA购自西安晶博试剂公司,含量≥99.5%,分析纯试剂AR级。
1.2 试验方法
1.2.1 材料预处理 将秦岭高山杜鹃幼苗移栽在直径为 15 cm 左右的花盆中,每盆栽幼苗3株,基质为腐叶土与松针土,体积比1 ∶ 2。处理期间每天傍晚喷施0.1%硫酸亚铁水溶液,经过1周的缓苗期后,每天18:00叶面喷施外源物质ALA,连续喷施1周;ALA浓度分别为25、50、75、100、125 mg/L(溶液现用现配),每种浓度处理3盆;以蒸馏水作为对照,1周后选取生长状态大体一致的幼苗于人工气候箱中进行高温胁迫。胁迫温度:40 ℃/30 ℃(昼/夜),光照度 10 000 lx,高温处理4 d。每间隔24 h进行1次相关生理指标的测定、热害指数的统计和计算。
1.2.2 幼苗相关生理指标的测定 分别称取不同处理下的高山杜鹃叶片0.2 g,测定其叶绿素含量(叶绿素a含量、叶绿素b含量、叶绿素a+b含量和叶绿素b/叶绿素a)、游离脯氨酸含量、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性和过氧化氢酶(CAT)活性。通过观察并统计高山杜鹃热害指数分析其热害情况。测定方法:叶绿素含量采用丙酮-碳酸钙法[14]测定,脯氨酸含量采用邹琦的磺基水杨酸法[15]测定;MDA含量测定参考硫代巴比妥酸(TBA)[14]检测法。SOD活性采用李合生[16]氮蓝四唑(NBT)法测定;POD活性的测定采用愈创木酚法[14];CAT活性测定采用紫外吸收法测定[16]。以上指标重复测定3次,求其平均值,热害指数参考赵冰等的方法[17]计算。
2 结果与分析
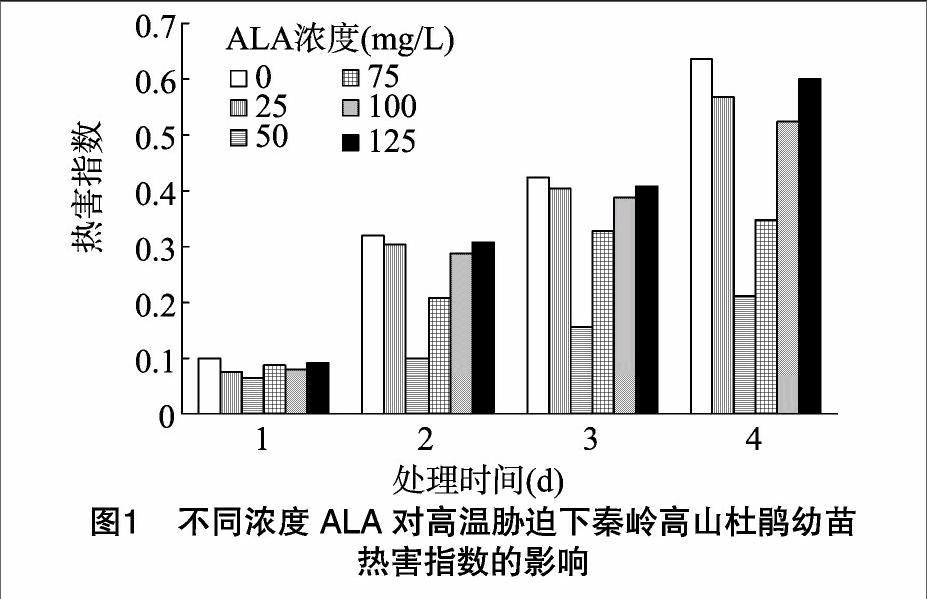
2.1 ALA对高温胁迫下秦岭高山杜鹃幼苗热害指数的影响
在逆境条件下,植物形态特征的变化可作为评价植物受损情况及抗逆性强弱的直接指标。在高温处理1 d后,高山杜鹃幼苗出现了不同程度的热损伤,随着时间的延长,植株叶片开始出现萎缩枯黄,且枯黄面积不断加大。对照组热害指数持续上升且高于所有处理组,而经不同浓度外源ALA处理后杜鹃幼苗热害指数增长相对缓慢且均低于对照组。结果表明,高温胁迫抑制了高山杜鹃幼苗生长,适宜浓度ALA通过缓解高温胁迫对植物造成的损伤,提高植物的耐热性。当ALA浓度为50 mg/L时,热害指数增长最缓慢,植株热损伤程度最低,生长状况相对较好(图1)。
2.2 ALA对高温胁迫下秦岭高山杜鹃幼苗生长相关生理指标的影响
2.2.1 ALA对高温胁迫下秦岭高山杜鹃幼苗叶片叶绿素含量的影响 高温逆境下,2种叶绿素含量都呈现下降的趋势(图2),这说明高溫胁迫显然抑制了秦岭高山杜鹃幼苗叶片中叶绿素含量。经过不同浓度的ALA处理后,叶绿素a、叶绿素b、叶绿素a+b含量和叶绿素b/叶绿素a多表现出先上升后下降的趋势,且均高于对照组的结果。25~100 mg/L ALA高温处理3 d时2种叶绿素含量及叶绿素b/叶绿素a的值均达到最大,且50 mg/L ALA处理的效果最好,在此浓度下秦岭高山杜鹃幼苗叶片中光合色素含量最大,光合作用也最强(图2至图5)。由此可见,适宜浓度的外源ALA可提高光合色素的含量,且有效地缓解了高温胁迫对高山杜鹃幼苗叶片光合色素合成的抑制效应,促进了秦岭高山杜鹃叶片的光合作用,有利于植物在逆境下生长。endprint
2.2.2 ALA对高温胁迫下秦岭高山杜鹃幼苗叶片脯氨酸含量的影响 高温胁迫期间,脯氨酸含量呈现缓慢上升趋势,外施不同浓度ALA处理后,游离脯氨酸含量均高于对照组,当ALA浓度为50 mg/L时,脯氨酸含量上升最快,且在处理4 d达到最大值(图6)。由此可见,秦岭高山杜鹃在经历高温胁迫时脯氨酸含量呈上升趋势,而叶面喷施不同浓度的外源ALA促进了脯氨酸含量的积累,提高了植物的渗透调节能力,缓解了热害对植物的损伤。
2.2.3 ALA对高温胁迫下秦岭高山杜鹃幼苗叶片MDA含量的影响 随着高温胁迫时间的延长,高山杜鹃幼苗叶片MDA含量呈上升趋势(图7),说明高温逆境下细胞质膜发生了过氧化作用,引起质膜正常的生理功能发生紊乱。通过不同浓度的ALA处理后,随着浓度升高,处理2~4 d MDA含量呈现先降后升的趋势,在ALA浓度为50 mg/L时,叶片中MDA含量大幅度降低至所有处理中最低,减缓的效果最为明显,75、100 mg/L ALA处理也不同程度地缓解了高温胁迫引起的MDA量升高的趋势,但效果不明显。而当ALA浓度为125 mg/L时,处理3~4 d MDA含量则高于对照组。结果说明,适宜浓度的外源ALA处理能够有效地减缓高温胁迫下高山杜鹃幼苗生长受到的伤害。
2.2.4 ALA对高温胁迫下秦岭高山杜鹃幼苗叶片SOD、POD、CAT活性的影响 随着高温胁迫时间的延长,3种抗氧化酶的活性呈先上升后下降的趋势(图8至图10),这说明高山杜鹃幼苗对胁迫的反应比较明显。经不同浓度的外源ALA处理后,3种抗氧化酶的活性均整体呈现出升高的趋势,并与对照形成明显差异,且当ALA达到50 mg/L时,其效果最好。SOD、POD、CAT活性随着时间的推移持续上升,在处理1 d时上升幅度均达到最大值,分别为60.21、59.69、30.98 U/mg;处理2~4 d,虽在75、100 mg/L ALA处理时保护酶活性有所降低,但仍高于对照组,当ALA浓度为 125 mg/L 时3种保护酶活性最低,且低于对照。
3 讨论
植物在其整个生长发育过程中常常会遭受各种非生物因子的胁迫, 其中温度是影响植物生长发育的重要生态因子之一。高温逆境下,植物起初表现为生理代谢紊乱,通过启动自身的调控系统维持在逆境下生长,当高温损伤超过植物本身的防御能力时,植物就会表现出相应的热害症状,严重时萎蔫死亡。本试验结果表明,随着高温胁迫时间的延长,秦岭高山杜鹃叶片失绿变黄甚至萎蔫枯萎,热害指数呈现逐渐上升的趋势,经不同浓度的ALA处理后,热害指数上升程度减弱,且50 mg/L ALA处理的效果最好,热害指数最低。由此表明,叶面喷施ALA可以降低植株的热害指数,缓解热损伤,提高植株的耐热性。
叶绿素含量与光合机能大小密切相关。ALA能够调节植物叶绿素的合成,促进叶绿素a转化为叶绿素b,防止叶绿素酶分解,从而保护光合器官。Downton等研究表明,叶绿素含量下降是引起光合速率降低的根本原因[18-19]。汪良驹等研究表明,ALA处理可以提高甜瓜叶片叶绿素含量,尤其是叶绿素b的含量,从而导致叶绿素b/叶绿素a比值增大,有利于植株在弱光下捕获光能,提高光合速率[20]。本试验中,高温胁迫后秦岭高山杜鹃幼苗叶片的叶绿素含量呈持续下降趋势,说明高温胁迫影响了植物体内叶绿素的合成,对植物的光合器官造成了损害。经不同浓度的外源ALA处理后,叶绿素a、叶绿素b、叶绿素a+b含量和叶绿素b/叶绿素a均有不同程度的提高,且均高于对照组,外施50 mg/L ALA处理后高山杜鹃幼苗叶片叶绿素含量和叶绿素b/叶绿素a均达到最大值。由此表明,不同浓度的外源ALA处理,能不同程度地促进叶绿素的合成,减缓高温胁迫下秦岭高山杜鹃幼苗叶片叶绿素含量和叶绿素b/叶绿素a的下降,从而改善高山杜鹃叶片光合作用的进行,有利于植物在逆境下生长。
脯氨酸可以反映植物抗逆性的强弱,主要起到维持渗透调节、防止植物体内水分的丢失、提高原生质胶体的稳定性等作用。正常情况下,脯氨酸合成酶类通过反馈抑制作用控制植物体内游离脯氨酸的含量,当植物处于逆境条件下,脯氨酸合成酶类对脯氨酸的反馈抑制效应降低,导致体内游离脯氨酸含量升高[21]。在逆境胁迫条件下,耐热植物与不耐热植物相比,体内积累了更多的脯氨酸,并且耐热性强弱与植物体内脯氨酸维持积累的时间长短呈现正相关[22]。徐刚等研究结果表明,ALA通过提高植物体内渗透调节物质含量以及抗氧化酶活性来提高植物的抗逆性[23]。本试验结果表明,与对照组相比,外施不同浓度的ALA可提高高温逆境下高山杜鹃幼苗的脯氨酸含量,且ALA浓度为50 mg/L时,作用效果最好,脯氨酸含量最高。
Martineau等研究认为,高温伤害的本质是植物细胞膜受损及膜透性的增加[24],而MDA为膜脂过氧化的产物之一,可作为脂质过氧化的指标来衡量细胞膜脂过氧化程度和植物抗逆性的强弱。有研究表明,植物抗热性的强弱与高温胁迫下MDA含量的增加呈现负相关[25]。徐晓洁等研究表明,外施ALA减缓了番茄幼苗体内MDA的积累,缓解了逆境对植物的伤害[26]。本试验中,高温胁迫后,MDA的含量持续升高,当喷施一定浓度的ALA处理后,MDA的含量有不同程度的降低,说明外源ALA能够抑制膜脂过氧化作用,降低膜系统受到的损害,提高膜的稳定性,提高植物在逆境下的生存能力。
超氧化物歧化酶(SOD)是生物体内主要的抗氧化酶,它可催化超氧阴離子自由基( O-2 · )生成O2和H2O2,而生成的H2O2则由POD、CAT清除。这3种酶的活性高低直接反映了植物抗逆性的强弱。王凯红等认为这3种抗氧化酶与杜鹃属植物的耐热性在很大程度上具有相关性,可用来鉴定杜鹃属植物的耐热性[27]。本试验结果表明,高温胁迫下秦岭高山杜鹃幼苗叶片中抗氧化酶(SOD、POD、CAT)活性均有提高,、外施不同浓度的ALA后,SOD、POD、CAT活性随着浓度升高呈先升后降趋势,且在ALA浓度为50 mg/L时活性最大,当ALA浓度为125 mg/L时3种保护酶活性最低,且多低于对照,说明高浓度ALA反而会降低其活性,加剧高温胁迫的影响。endprint
综上所述,ALA可以提高高温胁迫下秦岭高山杜鹃幼苗叶片叶绿素含量及SOD、POD、CAT活性,降低MDA含量,促进叶片内游离脯氨酸含量的积累,并在一定程度上降低热害指数。50 mg/L ALA叶面喷施处理对提高秦岭高山杜鹃耐热性相对效果最好。
参考文献:
[1]赵 冰,杜宇科,付玉梅,等. 镇安木王国家森林公园野生杜鹃花资源调查[J]. 安徽农业科学,2010,38(8):4000-4001,4041.
[2]赵 冰,张 果,司国臣,等. 秦岭野生杜鹃花属植物种质资源调查研究[J]. 西北林学院学报,2013,28(1):104-109.
[3]赵艳艳,胡晓辉,邹志荣,等. 不同浓度5-氨基乙酰丙酸(ALA)浸种对NaCl胁迫下番茄种子发芽率及芽苗生长的影响[J]. 生态学报,2013,33(1):62-70.
[4]马 娜,齐 琳,高晶晶,等. 5-ALA对高温下无花果扦插幼苗的生长及叶片叶绿素荧光特性的影响[J]. 南京农业大学学报,2015,38(4):546-553.
[5]张严玮,房伟民,黄素华,等. 外源ALA对低温胁迫下切花菊光合作用及生理特性的影响[J]. 南京农业大学学报,2014,37(1):47-52.
[6]张春平,何 平,袁凤刚,等. 外源5-氨基乙酰丙酸对干旱胁迫下草珊瑚叶绿素荧光特性及能量分配的影响[J]. 中草药,2012,43(1):164-172.
[7]张春平,何 平,韦品祥,等. 外源5-氨基乙酰丙酸对盐胁迫下紫苏种子萌发及幼苗抗氧化酶活性的影响[J]. 中草药,2011,42(6):1194-1200.
[8]汪良驹,石 伟,刘 晖,等. 外源5-氨基乙酰丙酸处理对小白菜叶片的光合作用效应[J]. 南京农业大学学报,2004,27(2):34-38.
[9]姚素梅,王维金. ALA对两系杂交粳稻子粒灌浆和产量的影响[J]. 华中农业大学学报,2004,23(5):495-499.
[10]柳翠霞,罗庆熙,李跃建,等. 外源5-氨基乙酰丙酸(ALA)对弱光下黄瓜生长指标及抗氧化酶活性的影响[J]. 中国蔬菜,2011,26(16):72-78.
[11]尹璐璐,于贤昌,王英华,等. 5-氨基乙酰丙酸对黄瓜幼苗抗冷性的影响[J]. 西北农业学报,2007,16(4):166-169.
[12]刘 涛,郭世荣,徐 刚,等. 5-氨基乙酰丙酸对辣椒植株低温胁迫伤害的缓解效应[J]. 西北植物学报,2010,30(10):2047-2053.
[13]Bindu R C,Vivekanandan M. Hormonal activites of 5-aminolevulinic acid in callus induction and micro propagation[J]. Plant Growth Regulation,1998,26(1):854-859.
[14]張志良,翟伟菁,李小方. 植物生理学实验指导[M]. 北京:高等教育出版社,2009:274.
[15]邹 琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2000:161-162.
[16]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:260-261.
[17]赵 冰,付玉梅,丁惠惠,等. Ca2+处理对秦岭高山杜鹃耐热性的研究[J]. 西北林学院学报,2010,25(6):29-32.
[18]Downton W J S,Loveys B R,Grant W J R. Stomatal closure fully accounts for the inhibition of photosynthesis by abscisic acid[J]. New Phytologist,1988,108(3):263-266.
[19]李小玲,华智锐. 外源ALA对盐胁迫下商洛黄芩生理特性的影响[J]. 江西农业学报,2016,28(2):70-73.
[20]汪良驹,姜卫兵,黄保健. 5-氨基乙酰丙酸对弱光下甜瓜幼苗光合作用和抗冷性的促进效应[J]. 园艺学报,2004,31(3):321-326.
[21]杨少辉. 高温胁迫对植物影响的研究进展[J]. 分子植物育种,2006,4(3):139-142.
[22]张春平,何 平,魏品祥,等. 高温胁迫对不同基因型紫苏种子萌发,幼苗抗氧化酶活性的影响[J]. 中草药,2011,42(6):1194-1200.
[23]徐 刚,刘 涛,高文瑞,等. 5-氨基乙酰丙酸对蔬菜生理作用的研究进展[J]. 金陵科技学院学报,2010,26(4):52-57.
[24]Martineau J R,Specht J E. Temperature tolerance in soybeans[J]. Crop Science,2009,19:75-81.
[25]陈文志. 药物处理对报春花抗热性的影响研究[D]. 雅安:四川农业大学,2007:23-25.
[26]徐晓洁,邹志荣,乔 飞,等. ALA对NaCl胁迫下不同品种番茄植株光合作用、保护酶活性及果实产量的影响[J]. 干旱地区农业研究,2008,26(4):131-135.
[27]容 丽,陈 训,汪小春. 百里杜鹃属13种植物叶片解剖结构的生态适应性[J]. 安徽农业科学,2009,37(3):1084-1088.endprint
猜你喜欢
铝加工(2017年1期)2017-03-07
山东农业科学(2016年12期)2017-01-21
现代园艺(2016年9期)2016-07-16
现代园艺(2016年2期)2016-03-15
山西果树(2015年6期)2015-12-11
中国塑料(2015年6期)2015-11-13
质量与标准化(2015年11期)2015-07-21
福建林业(2015年2期)2015-04-09
