
Saponins from Panax japonicus attenuate age-related neuroinflammation via regulation of the mitogenactivated protein kinase and nuclear factor kappa B signaling pathways
2017-12-15 09:23LiliDengDingYuanZhiyongZhouJingzhiWanChangchengZhangChaoqiLiuYaoyanDunHaixiaZhaoBoZhaoYuanjianYangTingWang
中国神经再生研究(英文版) 2017年11期
Li-li Deng, Ding Yuan, Zhi-yong Zhou Jing-zhi Wan Chang-cheng Zhang Chao-qi Liu Yao-yan Dun Hai-xia Zhao Bo ZhaoYuan-jian Yang, Ting Wang
1 College of Medical Science, China Three Gorges University, Yichang, Hubei Province, China
2 Renhe Hospital, Second College of Clinical Medical Science, China Three Gorges University, Yichang, Hubei Province, China
3 Medical Experimental Center, Jiangxi Mental Hospital/Affiliated Mental Hospital of Nanchang University, Nanchang, Jiangxi Province, China
How to cite this article:DengLL, Yuan D, Zhou ZY, Wan JZ, Zhang CC, Liu CQ, Dun YY, Zhao HX, Zhao B, Yang YJ, Wang T (2017) Saponins from Panax japonicus attenuate age-related neuroinflammation via regulation of the mitogen-activated protein kinase and nuclear factor kappa B signaling pathways. Neural Regen Res 12(11):1877-1884.
Funding: This work was supported by the National Natural Science Foundation of China, No. 81374001, 81673778, 81273895; the Foundation for Innovative Research Groups of the Natural Science Foundation of Hubei Province of China, No. 2013CFA014.
Introduction
Neuroinflammation is associated with the onset and evolution of many dementia-related neurodegenerative disorders(Lu et al., 2011). Furthermore, it is characterized by increased microglial activation and often accompanied by upregulation of inflammatory cytokines, such as tumor necrosis factor(TNF-α) and interleukin-1β (IL-1β) (Ownby, 2010; Hein et al., 2012). Many studies have shown the appearance of neuroin-flammatory changes with aging, even in the healthy brain. These accumulative changes can be neurotoxic and alter important neuronal functions, ultimately contributing to the pathogenesis of neurodegenerative disorders (McGeer et al., 2004; Gemma et al., 2007; Lynch MA, 2010; Pizza et al.,2011).
It is well known that microglial cells are the resident brain macrophages, and the first line of defense against central nervous system injury and microbial invasion (Pierre, 2017).Accumulating evidence indicates that microglia activation in aging models results in proinflammatory cytokine secretion,which leads to subsequent neuronal damage and synaptic and cognitive impairment (Bardou et al., 2013; Farso et al., 2013).Thus, inhibition of microglial activation and the resultant neuroinflammation is a promising strategy for alleviating inflammation-related neurodegenerative disease (Liu et al., 2003).
Rhizoma Panacis japonica, a common traditional herbal medicine, is the dried rhizome ofPanax japonicus, and widely used by the Hmong and Tujia people of China. In the plant kingdom, it belongs to the Araliaceae Panax family. It is widespread throughout Japan and southwest China and some ethnic minority groups, and is used as a substitute for Ginseng root (Li et al., 2012). Saponins fromPanax japonicus(SPJ) are the bioactive rhizome component ofPanax japonicus, and can easily be obtained (Ou et al., 2010). There are dammarane-type and oleanolic acid-type saponins, which mainly contain oleanolic acid alkane pentacyclic triterpenes (Zou et al., 2002). Moreover,SPJ has been shown to have many pharmacological effects, such as anti-fatigue, anti-apoptosis, antioxidant, and anti-inflammation (Yamahara et al., 1987; Qian et al., 2008; Li et al., 2010). A previous study suggested that SPJ exerts an anti-inflammatory effect in lipopolysaccharide-stimulated RAW 264.7 cells, mainlyviainhibition of p38, extracellular signal-regulated kinase 1/2 (ERK1/2), mitogen-activated protein kinase (MAPK), and nuclear factor kappa B (NF-κB) activation (Dai et al., 2014). SPJ has also been shown to attenuate D-galactose-induced cognitive impairment by anti-apoptotic and anti-oxidative effects in rats(Wang et al., 2015). However, whether SPJ has an anti-inflammatory effect during brain aging, and correspondingly, the underlying mechanisms remain unknown. Here, our study aimed to investigate age-related neuroin-flammatory changes and the effect of SPJ on neuroinflammation during natural brain aging,as well as potential mechanisms.
Materials and Methods
Animals
Seventy male specific-pathogen-free Sprague-Dawley rats weighing 180–200 g were obtained from the Laboratory Animal Center of China Three Gorges University of China(license No. SCXK (E) 2012-0001). Rats were housed under standard conditions in controlled humidity (50 ± 10%) and temperature (23 ± 3°C) conditions, and a 12-hour light/dark cycle with free access to water and food.
The study protocol was approved by the Animal Ethics Committee of China Three Gorges University (approval No.2012030D). The experimental procedure followed the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1986).
Preparation of SPJ
FreshPanax japonicuswas obtained from the Enshi Chunmuying Medicinal Materials Planting Base (Enshi, China), and authenticated by Dr. Kun Zou from Hubei Key Laboratory of Natural Products Research and Development (China Three Gorges University, Yichang, Hubei Province, China). SPJ extraction and analysis has been previously reported (He et al.,2012). Briefly, driedPanax japonicusroot (1 kg) was crushed and extracted three times with 10 L of 60% ethanol by recirculation for 2 hours. The ethanol extract (extract rate 58%) was then dissolved in distilled water and further extracted separately with petroleum ether, ethyl acetate, andn-butanol. Then-butanol extract (extract rate 37%) was redissolved in water,passed through a chromatographic column containing macroporous adsorption resin (D101; Tianjin Pesticide Factory,Tianjin, China), rinsed with water, and then eluted with 30%,60%, and 90% ethanol in water to produce three fractions. The 90% fraction was de fined as the re finedn-butanol extract. Refinedn-butanol extract (extract rate 16.8%) was redissolved in water at a concentration of 10 mg/mL and diluted with methanol-water to 1 mg/mL. High performance liquid chromatography analysis was performed using a SEYMC-Pack ODS-AQ column (4.6 mm × 250.0 mm, 5 µm). The sample was eluted with mobile phases of acetonitrile (A) and 0.4% phosphate solution (B) at a flow rate of 1.0 mL/min. The system was run with a gradient program: 0–5 minutes, 5% A; 5–20 minutes,5–30% A; 20–30 minutes, 30% A; 30–50 minutes, 30%–85% A;and 50–60 minutes, 85% A. Sample injection volume was 10 μL, with an ultraviolet detector used for detection.
Drug administration
Sprague-Dawley rats were divided into control groups (3-,9-, 15-, and 24-month-old groups) and SPJ-treated groups,with 10 individuals in each group. For SPJ-treated groups,18-month-old rats were orally administrated SPJ at three doses of 10, 30, or 60 mg/kg once daily for 6 months until they were 24 months old. Rats in the 3-, 9-, 15-, and 24-monthold control and SPJ-treated groups were rapidly decapitated.Prefrontal cortex and hippocampal tissue were harvested and stored at −80°C for analyses.
Immunohistochemical staining
Brain tissue was embedded in paraffin, cut into 5 µm thick sections, deparaffinized in xylene, and rehydrated with graded alcohol. Citrate buffer (pH 6.0) was used for antigen retrieval,with brain sections incubated for 5 minutes at 95–100°C in a water bath. After cooling and incubating with 5% bovine se-rum albumin for 20 minutes to block endogenous peroxidase activity, goat polyclonal anti-ionized calcium-binding adapter molecule 1 (Iba1) primary antibody was used (diluted 1:200 in PBS solution; Abcam, Cambridge, UK) to identify microglia, with sections incubated overnight at 4°C in a humidified chamber. After washing three times with PBS, polyvalent biotinylated donkey anti-goat IgG (H + L) secondary antibody (diluted 1:2,000; KPL Company, Maryland, WA, USA)was added for 20 minutes at room temperature, followed by incubation with streptavidin peroxidase for 20 minutes. Subsequently, 3,3′-diaminobenzidine was used as the chromogen for color development. Finally, sections were counterstained with hematoxylin for 2 minutes, dehydrated, cleared in xylene,and fixed in mounting media. In five consecutive sections,1,500 µm2of the same area was examined using an TP1020 inverted fluorescence microscope (Leica, Wetzlar, Germany).Semi-quantitative evaluation was performed using the Photo and Image Autoanalysis System (Leica), with a person who was blinded to the experiment.
Western blot assay
Total protein and nuclear protein were extracted from the prefrontal cortex and hippocampus using the Total Protein Extraction Kit (P1250; Applygen Technologies Inc., Beijing,China) and Nuclear and Cytosol Fractionation Kit (P1200;Applygen Technologies Inc.), respectively, in accordance with the manufacturer’s instructions. A bicinchoninic acid protein assay kit was used to determine protein concentration.Samples (50 μg each protein) were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes by electrophoretic transfer. After blocking in 5% nonfat milk in Tris-buffered saline with Tween for 1 hour at room temperature,membranes were incubated at 4°C overnight with primary antibodies. The primary antibodies used were TNF-α (goat polyclonal antibody, diluted 1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), IL-1β (rabbit polyclonal antibody,diluted 1:500; Santa Cruz Biotechnology), inducible nitric oxide synthase (iNOS) (mouse monoclonal antibody, diluted 1:500; Santa Cruz Biotechnology), cyclooxygenase-2 (COX-2)(rabbit monoclonal antibody, diluted 1:1,000; Cell Signaling,Danvers, MA, USA), NF-κB p65 (mouse monoclonal antibody, diluted 1:1,000; Cell Signaling), lamin A (rabbit polyclonal antibody, diluted 1:1,000; Santa Cruz Biotechnology),phospho-NF-κB (p-NF-κB) p65 (rabbit polyclonal antibody,diluted 1:1,000; Cell Signaling), phospho-inhibitor of kappa B(p-IκBα) (mouse monoclonal antibody, diluted 1:1,000; Cell Signaling), IκBα (rabbit monoclonal antibody, diluted 1:1,000;Cell Signaling), ERK (rabbit polyclonal antibody, diluted 1:500, Santa Cruz Biotechnology), phospho-ERK (p-ERK)(mouse monoclonal antibody, diluted 1:500; Santa Cruz Biotechnology), c-Jun N-terminal kinase (JNK) (rabbit polyclonal antibody, diluted 1:1,000; Cell Signaling), phospho-c-Jun N-terminal kinase (p-JNK) (rabbit monoclonal antibody,diluted 1:1,000; Cell Signaling), p38 (rabbit polyclonal antibody, diluted 1:1,000; Cell Signaling), phospho-p38 (p-p38)(rabbit monoclonal antibody, diluted 1:1,000; Cell Signaling),and β-actin (mouse monoclonal antibody, diluted 1:1,000;Santa Cruz Biotechnology). Afterwards, horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG was incubated for 1 hour at room temperature. Target protein bands were observed by enhanced chemiluminescence on Kodak film.Relative protein levels were quantified by scanning densitometry and analyzed by ImageJ software (Materialize NV,Leuven, Belgium). Results were normalized to gray values of β-actin or lamin A expression in each group.
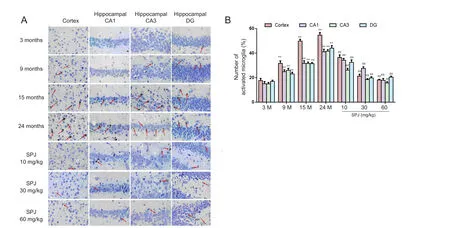
Figure 1 Effect of SPJ on microglia morphology (Iba-1-immunoreactive cells) in the cortex and hippocampus of aging rats .(A) Microglia morphology was observed by immunohistochemistry using a TP1020 inverted fluorescence microscope (original magnification,400×). Red arrows indicate microglia activation. (B) Quantification of activated microglia in prefrontal cortex and hippocampal CA1, CA3,and dentate gyrus regions. **P < 0.01, vs. 3-month-old group; ##P <0.01, vs. 24-month-old group (mean ± SD, n = 5, one-way analysis of variance followed by the least significant difference post hoc test). 3-,9-, 15-, and 24-month-old groups (intragastric saline); SPJ was administrated at 10, 30, and 60 mg/kg once daily for 6 months until the rats were 24 months old. SPJ: Saponins from Panax japonicus; M: months.
Statistical analysis
Data are expressed as the mean ± SD. All statistical analyses were performed using GraphPad Prism 10.0 for the Macintosh software (GraphPad Software, San Diego, CA, USA). Differences between groups were examined by one-way analysis of variance.Post hoctesting was performed for inter-group comparisons using the least significant difference test.P< 0.05 was considered statistically significant.
Results
SPJ inhibited microglial activation in the cortex and hippocampus of aging rats
Microglia activation was detected by immunohistochemistry of Iba-1 protein, which is a specific microglia marker. In the prefrontal cortex and hippocampal CA1, CA3, and dentate gyrus regions, we found that aging induced microglial activation, as displayed by an irregular shape, enlarged cell size, thickened and shortened processes, and intensified Iba1 immunostaining density (Figure 1A;P< 0.01). Furthermore, Iba-1-positive cell number increased gradually with aging. Compared with 3-month-old rats, the number of activated microglia in 24-month-old rats increased 4.1-times, 3.7-times, 4.8-times,and 7.3-times in the prefrontal cortex and hippocampal CA1,CA3, and dentate gyrus, respectively (Figure 1B;P< 0.01).Conversely, SPJ treatment mitigated these morphological changes in activated microglia, and reduced the number of activated microglia in the prefrontal cortex and hippocampal regions compared with 24-month-old rats.
SPJ suppressed TNF-α and IL-1β protein levels in the cortex and hippocampus of aging rats
To investigate the effect of SPJ on proinflammatory cytokine expression in aging rats, IL-1β and TNF-α protein expression was detected by western blotting. Our results show an age-related increase in IL-1β and TNF-α protein levels in the cortex and hippocampus. Compared with the 3-month-old group,TNF-α protein levels in 24-month-old rats increased 3.6-fold and 4.5-fold in the cortex and hippocampus, respectively (P< 0.01). Similarly, IL-1β protein levels increased in the same regions of 24-month-old rat brain by 3.3-fold and 6.5-fold,respectively (P< 0.01). While SPJ treatment significantly attenuated the increased IL-1β and TNF-α protein expression levels in aging rats (Figure 2;P< 0.01 orP< 0.05). These data demonstrate that SPJ inhibits production of proinflammatory cytokines such as TNF-α and IL-1β in aged rats.
SPJ attenuated iNOS and COX-2 protein levels in the cortex and hippocampus of aging rats
Protein levels of iNOS and COX-2 increased age-dependently in the cortex and hippocampus (Figure 3). Compared with the 3-month-old group, iNOS protein levels in the cortex and hippocampus of the 24-month-old group increased 2.9-fold and 2.5-fold, respectively (P< 0.01). Similarly, COX-2 protein levels increased 6.1-fold and 11.8-fold in the cortex and hippocampus, respectively, of 24-month-old aging rats. After SPJ treatment for 6 months (P< 0.01), iNOS and COX-2 protein levels decreased significantly in the 24-month-old group compared with the 24-month-old non-SPJ treatment group (P< 0.01 orP< 0.05). These findings demonstrate that SPJ inhibits expression of inflammatory mediators such as COX-2 and iNOS.
SPJ inhibited the NF-κB signaling pathway in the cortex and hippocampus of aging rats
Previous studies have suggested that NF-κB activation is detected in microglia and contributes to neuroinflammation(Kaur et al., 2015; Park et al., 2015). To determine whether the NF-κB signaling pathway is involved in the anti-neuroin-flammatory effect of SPJ on aging rats, we examined nuclear NF-κBp65 and p-NF-κBp65 protein levels by western blot assay.Nuclear NF-κBp65 and p-NF-κBp65 protein levels were both enhanced age-dependently in the cortex and hippocampus(Figure 4). Compared with 3-month-old rats, nuclear NF-κBp65 protein levels increased 9.5-fold and 6.3-fold in the cortex and hippocampus, respectively, of 24-month-old aging rats. Similarly, p-NF-κBp65 protein levels increased 4.5-fold and 2.5-fold, respectively, in the cortex and hippocampus of 24-month-old aging rats. In contrast, aging-induced enhancement of nuclear NF-κBp65 and p-NF-κBp65 protein levels in the cortex and hippocampus were significantly suppressed when 24-month-old aging rats were treated with SPJ for 6 months (P< 0.05 orP< 0.01). In the unstimulated condition,cytoplasmic NF-κB is sequestered by IκBα. Upon stimulation, IκBα is rapidly phosphorylated or degraded (Wan et al.,2010). Phosphorylated and degraded IκBα levels increased with age, and were signifi-cantly suppressed by SPJ treatment in 24-month old aging rats (Figure 4;P< 0.01 orP< 0.05). These findings clearly demonstrate that SPJ attenuates aging-induced neuroinflammation, partly by inhibition of NF-κB activation.
SPJ modulated the MAPK pathway in the cortex and hippocampus of aging rats
The MAPK signaling pathway (which includes ERK, JNK,and p38) has previously been reported to activate NF-κB and further initiate pro-inflammatory responses (Liu et al., 2015a,b). Accordingly, we next examined phosphorylation levels of MAPK signaling in the cortex and hippocampus by western blot assay. We detected age-related increases in phosphorylation levels of ERK, JNK, and p38 in the cortex and hippocampus (Figure 5;P< 0.01 orP< 0.05). Treatment with SPJ in 24-month-old aging rats markedly suppressed aging-induced phosphorylation levels of ERK, JNK, and p38 (Figure 5;P< 0.01 orP< 0.05). These results show that inhibition of aging-associated neuroinflammation by SPJ is mediated, at least partly, by suppression of the MAPK pathway and its subsequent inhibition of NF-κB activation.
Discussion
We have previously shown in rats that SPJ has an anti-inflammatory effect and attenuates cognitive impairment via anti-oxidative and anti-apoptotic effects (Dai et al., 2014;Wang et al., 2015). In addition, accumulating evidence shows the aging brain is characterized by neuroinflammation and increased levels of inflammatory cytokines and activated microglia, which make the aged brain more vulnerable to disruptive effects from both intrinsic and extrinsic factors, such as stress, infection, or disease (Sparkman et al., 2008). Herein,we investigated the anti-inflammatory effects of SPJ in aging rat brain. Our results show for the first time that SPJ attenuates aging-induced neuroinflammation, possibly by suppressing microglial activation and inhibiting NF-κB and MAPK signaling components.
Mounting evidence indicates that age-related decline in cognitive performance is associated with altered levels of pro-inflammatory cytokines, a process that may be mediated by changes in microglia function (Godbout et al., 2009; Lekander et al., 2011). Age-associated increases in total number of activated microglia, and significant age-related alterations in microglial morphology and function have also been observed in postmortem human brain (DiPatre et al., 1997; Sheffield et al., 1998; Miller et al., 2007). Activated microglia secrete many neurotoxic factors that contribute to the pathogenesis of neurodegenerative disorders and affect complex central nervous system functions, such as depression, sleep, and cognition (Janelsins et al., 2008; Westman et al., 2014; Dursun et al., 2015; Sun et al., 2016). Thus, inhibition of microglial activation is a crucial mechanism in treatment of neurological disorders associated with inflammation. Previous studies have shown that ginsenoside Rg3 effectively suppresses proinflammatory cytokines, including IL-6, IL-1β, and TNF-α in lipopolysaccharide or amyloid β-stimulated BV-2 microglial cells(Bae et al., 2006; Joo et al., 2008), ultimately attenuating the inflammatory response. Consistent with these findings, our results show that SPJ significantly inhibits microglial activation and decreases production of pro-inflammatory cytokines(such as IL-1β and TNF-α) and inflammatory mediators (such as COX-2 and iNOS) in the aging brain.
Aging significantly triggers degradation and phosphorylation of cytosolic IκBα, and subsequent phosphorylation of cytosolic NF-κB p65 subunit and levels of nuclear NF-κB p65 subunit (Giardina et al., 2002). Under normal physiological conditions, NF-κB complexes are inactive in the cytoplasmic form and associated with an inhibitory protein termed IκB.Once activated by aging and other external antigens, NF-κB is rapidly freed from IκB, and translocates into the nucleus. NF-κB is regarded as a key regulator of inflammation, as it acts as a necessary transcription factor for induction of COX-2,iNOS, IL-1β, and TNF-α (Li et al., 2010). Emerging evidence suggests that NF-κB activation is found in microglia of patients with Alzheimer’s disease (AD), and misregulation of NF-κB can lead to AD development (Hu et al., 2005; Zhang et al., 2009; Lukiw 2012; Liu et al., 2015 a, b; Holtman et al.,2015). Taken together, these studies suggest that activated NF-κB plays a critical role in regulation of neuroinflammation in AD. Furthermore, it has been shown that ginsenoside Rg1, an active ingredient ofPanax ginseng, relieves tert-butyl hydroperoxide-induced cell impairment in mouse microglial BV2 cells through the NF-κB pathway (Lu et al., 2015). Similarly,our present study shows that SPJ prevents aging-induced NF-κB activation in a concentration-dependent manner. These studies suggest that SPJ may exert anti-inflammatory effects via downstream inflammatory factors and down-regulation of NF-κB activation in the aging brain.
NF-κB activation is also regulated by MAPK, which is composed of three components: p38, JNK, and ERK1/2 (Karin et al., 2000). An increasing number of studies indicate that hyperphosphorylation of the MAPK pathway activates NF-κB,producing inflammatory factors that ultimately contribute to neurodegenerative diseases (Waetzig et al., 2005; Lagalwar et al., 2006; Colombo et al., 2009; Petrov et al., 2015). Wang et al. (2014) also demonstrated increased p-p38 and JNK expression in AD and Parkinson’s disease patients. Moreover,p38/MAPK signaling plays a crucial role in pathogenesis of Parkinson’s disease, triggering undesired phenotypes such as microglia activation, neuroinflammation, oxidative stress,and apoptosis (Jha et al., 2015). Some ginsenosides (e.g., Rg1 and Rh2) are effective in treating acute ischemic chronic neuroinflammation and brain injury by blocking generation of cytokines and pro-inflammatory mediators through inhibition of NF-κB and MAPK activation (Choi et al., 2007; Huang et al., 2016; Quan et al., 2016). Here, our results also suggest that aging significantly triggers phosphorylation of p38,ERK1/2, and JNK in rat cortex and hippocampus. In contrast,SPJ treatment notably down-regulated phosphorylated levels of p38, ERK1/2, and JNK compared with 24-month aging rats, suggesting that SPJ has anti-neuroin-flammatory effects in aging ratsviaregulation of the MAPK signaling pathway.In summary, SPJ attenuates aging-induced neuroinflammation by suppression of pro-inflammatory cytokine expression, microglia activation, and inflammatory mediators in the rat cortex and hippocampus. This anti-inflammatory effect induced by aging in the cortex and hippocampus is possibly mediated by decreasing IκBα degradation, activation of NF-κB,and suppression of phosphorylation of p38, JNK, and ERK1/2.Our results suggest that SPJ has potent anti-inflammatory effects and may be a potential therapeutic strategy for the treatment of neurodegenerative-related diseases.
Author contributions:WT, YYJ, ZCC and LCQ designed this study. DLL,WJZ, DYY and ZB performed experiments. YD and ZZY analyzed data. ZHX and DLL wrote the paper. All authors approved the final version of the paper.
Conflicts of interest:None declared.
Research ethics:The study protocol was approved by the Animal Ethics Committee of China Three Gorges University (approval No. 2012030D).The experimental procedure followed the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1986). All efforts were made to minimize the number and suffering of the animals used in the experiment.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Bae EA, Kim EJ, Park JS, Kim HS, Ryu JH, Kim DH (2006) Ginsenosides Rg3 and Rh2 inhibit the activation of AP-1 and protein kinase A pathway in lipopolysaccharide/interferon-gamma-stimulated BV-2 microglial cells. Planta Med 72:627-633.
Bardou I, Brothers HM, Kaercher RM, Hopp SC, Wenk GL (2013) Differential effects of duration and age upon the consequences of neuroinflammation in the hippocampus. Neurobiol Aging 34:2293-2301.
Choi K, Kim M, Ryu J, Choi C (2007) Ginsenosides compound K and Rh(2)inhibit tumor necrosis factor-alpha-induced activation of the NF-kappaB and JNK pathways in human astroglial cells. Neurosci Lett 421:37-41.
Colombo A, Bastone A, Ploia C, Sclip A, Salmona M, Forloni G, Borsello T(2009) JNK regulates APP cleavage and degradation in a model of Alzheimer’s disease. Neurobiol Dis 33:518-525.
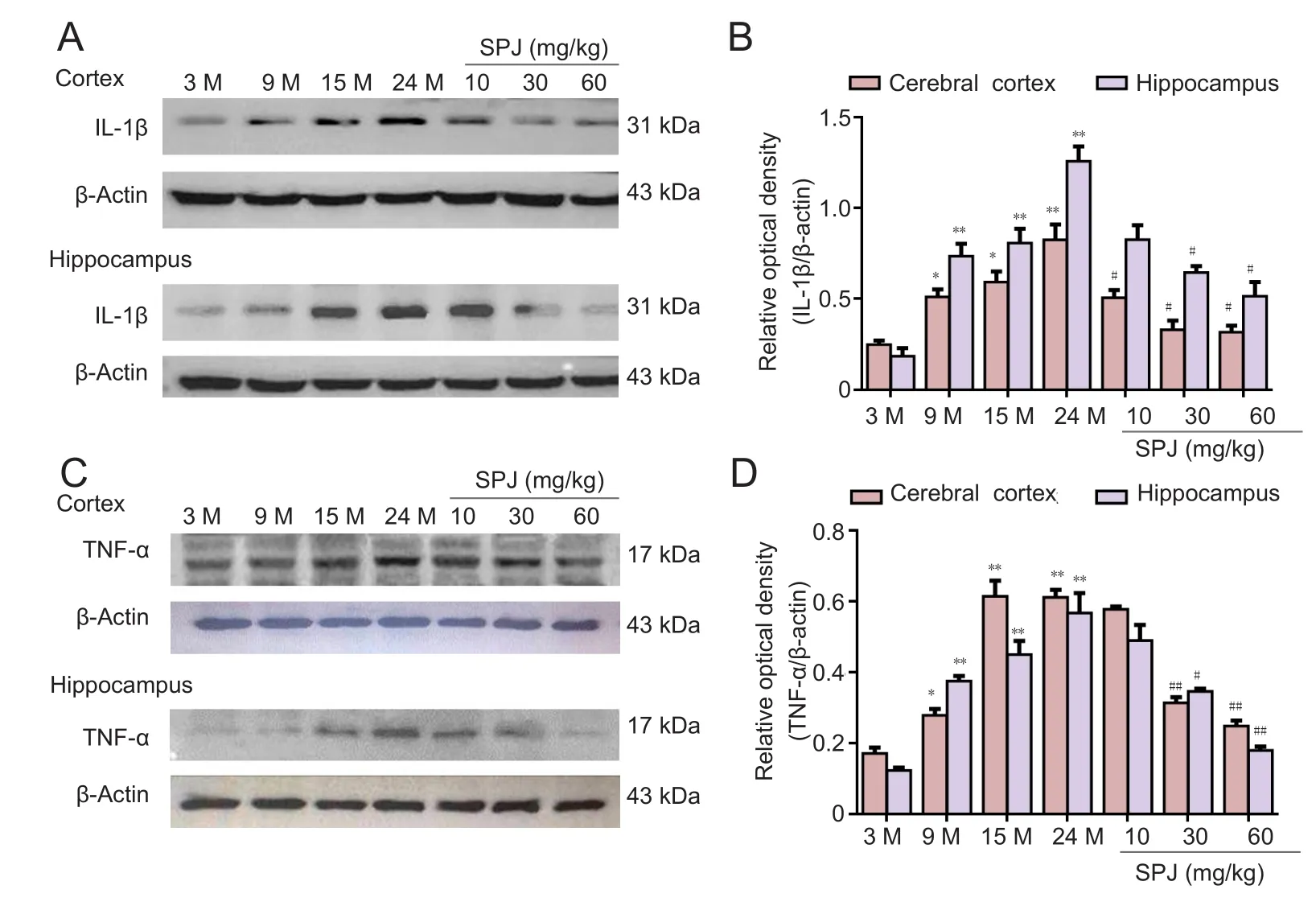
Figure 2 Effect of SPJ on IL-1β and TNF-α protein levels in the cortex and hippocampus of aging rats.(A) Representative IL-1β protein bands examined by western blot assay. (B) Data quantification of panel (A). (C) Representative TNF-α protein bands examined by western blot assay.(D) Data quantification of panel (C). *P < 0.05, **P < 0.01,vs. 3-month-old group; #P <0.05, ##P < 0.01, vs. 24-monthold group (mean ± SD, n = 4,one-way analysis of variance followed by the least significant difference post hoc test). 3-, 9-,15-, and 24-month-old groups(intragastric saline); SPJ was administrated at 10, 30, and 60 mg/kg once daily for 6 months until the rats were 24 months old. SPJ: Saponins from Panax japonicus; IL-1β: interleukin 1 beta; TNF-α: tumor necrosis factor alpha; M: months.
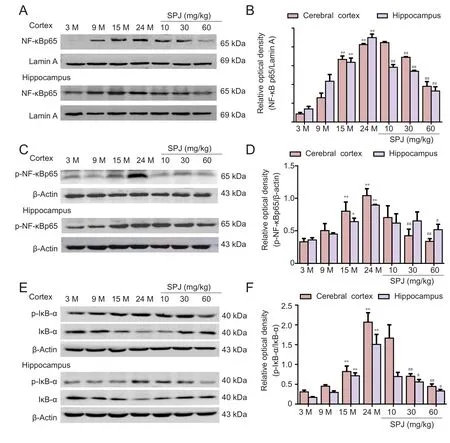
Figure 4 Effect of SPJ on the NF-κB signaling pathway in the cortex and hippocampus of aging rats.(A) Representative bands showing nuclear translocation of NF-κB p65 protein by western blot assay. (B) Data quantification of western blot assay (A). (C)Representative p-NF-κB p65 protein bands examined by western blot assay. (D) Data quantification of panel (C). (E)Representative p-IκB-α and IκB-α protein bands examined by western blot assay. (F) Data quantification of p-IκB-α. *P <0.05, **P < 0.01, vs. 3-monthold group; #P < 0.05, ##P < 0.01,vs. 24-month-old group (mean± SD, n = 4, one-way analysis of variance followed by the least significant difference post hoc test). 3-, 9-, 15-, and 24-monthold group (intragastric saline);SPJ was administrated at 10,30, and 60 mg/kg once daily for 6 months until the rats were 24 months old. SPJ: Saponins from Panax japonicus; p-NF-κB: phospho-NF-κB; p-IκB-α:phospho-IκB-α; M: months.
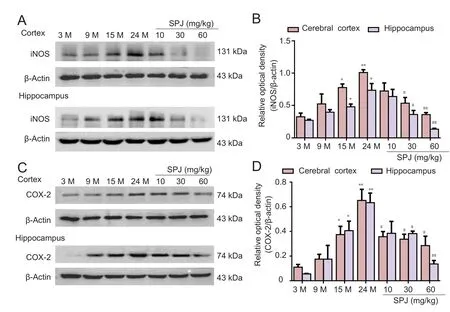
Figure 3 Effect of SPJ on iNOS and COX-2 production in the cortex and hippocampus of aging rats.(A) Representative iNOS protein bands examined by western blot assay. (B) Data quantification of panel (A). (C) Representative COX-2 protein bands examined by western blot assay. (D) Data quantification of panel (C). *P <0.05, **P < 0.01, vs. 3-month-old group; #P < 0.05, ##P < 0.01, vs.24-month-old group (mean ± SD,n = 4, one-way analysis of variance followed by the least significant difference post hoc test). 3-,9-, 15-, and 24-month-old groups(intragastric saline); SPJ was administrated at 10, 30, and 60 mg/kg once daily for 6 months until the rats were 24 months old. SPJ:Saponins from Panax japonicus;iNOS: inducible nitric oxide synthase; COX-2: cyclooxygenase-2;M: months.
Dai YW, Yuan D, Wan JZ, Zhang CC, Liu CQ, Wang T (2014) Saponins from Panax japonicus protect against LPS-induced RAW 264.7 macrophages Inflammation via NF-kB signaling pathways. Zhongguo Zhongyao Zazhi 11:2076-2080.
DiPatre PL, Gelman BB (1997) Microglial cell activation in aging and alzheimer disease: partial linkage with neurofibrillary tangle burden in the hippocampus. J Neuropathol Exp Neurol 56:143.
Dursun E, Gezen-Ak D, Hanağası H, Bilgiç B, Lohmann E, Ertan S, Atasoy İL, Alaylıoğlu M, Araz ÖS, Önal B, Gündüz A, Apaydın H, Kızıltan G,Ulutin T, Gürvit H, Yılmazer S (2015) The interleukin 1 alpha, interleukin 1 beta, interleukin 6 and alpha-2-macroglobulin serum levels in patients with early or late onset Alzheimer’s disease, mild cognitive impairment or Parkinson’s disease. J Neuroimmunol 283:50-57.
Farso M, Menard C, Colby-Milley J, Quirion R (2013) Immune marker CD68 correlates with cognitive impairment in normally aged rats. Neurobiol Aging 34:1971-1976.
Gemma C, Bickford PC (2007) Interleukin-1beta and caspase-1: players in the regulation of age-related cognitive dysfunction. Rev Neurosci 18:137-148.
Giardina C, Hubbard AK (2002) Growing old with nuclear factor–κB. Cell Stress Chaperones 7:207-212.
Godbout JP, Johnson RW (2009) Age and neuroinflammation: a lifetime of psychoneuroimmune consequences. Immunol Allergy Clin North Am 29:321-337.
He H, Xu J, Xu Y, Zhang C, Wang H, He Y, Wang T, Yuan D (2012) Cardioprotective effects of saponins from Panax japonicus on acute myocardial ischemia against oxidative stress-triggered damage and cardiac cell death in rats. J Ethnopharmacol 140:73-82.
Hein AM, O’Banion MK (2012) Neuroinflammation and cognitive dysfunction in chronic disease and aging. J Neuroimmune Pharmacol 7:3-6.
Holtman IR, Raj DD, Miller JA, Schaafsma W, Yin Z, Brouwer N, Wes PD,Möller T, Orre M, Kamphuis W, Hol EM, Boddeke EWGM, Eggen BJL(2015) Induction of a common microglia gene expression signature by aging and neurodegenerative conditions: a co-expression meta-analysis.Acta Neuropathol Commun 3:31.
Hu X, Nesic-Taylor O, Qiu J, Rea HC, Fabian R, Rassin DK, Perez-Polo JR(2005) Activation of nuclear factor-kappaB signaling pathway by interleukin-1 after hypoxia/ischemia in neonatal rat hippocampus and cortex. J Neurochem 93:26-37.
Huang L, Liu LF, Liu J, Dou L, Wang GY, Liu XQ, Yuan QL (2016) Ginsenoside Rg1 protects against neurodegeneration by inducing neurite outgrowth in cultured hippocampal neurons. Neural Regen Res 11:319-325.
Janelsins MC, Mastrangelo MA, Park KM, Sudol KL, Narrow WC, Oddo S,LaFerla FM, Callahan LM, FederoffHJ, Bowers WJ (2008) Chronic neuron-specific tumor necrosis factor-alpha expression enhances the local inflammatory environment ultimately leading to neuronal death in 3xTg-AD mice. Am J Pathol 173:1768-1782.
Jha SK, Jha NK, Kar R, Ambasta RK, Kumar P (2015) p38 MAPK and PI3K/AKT signalling cascades inparkinson’s disease. Int J Mol Cell Med 4:67-86.Joo SS, Yoo YM, Ahn BW, Nam SY, Kim YB, Hwang KW, Lee DI (2008)Prevention of inflammation-mediated neurotoxicity by Rg3 and its role in microglial activation. Biol Pharm Bull 31:1392-1396.
Karin M, Delhase M (2000) The I kappa B kinase (IKK) and NF-kappa B: key elements of proinflammatory signalling. Semin Immunol 12:85-98.
Kaur U, Banerjee P, Bir A, Sinha M, Biswas A, Chakrabarti S (2015). Reactive oxygen species, redox signaling and neuroinflammation in Alzheimer’s disease: the NF-κB connection. Curr Top Med Chem 15:446-457.
Lagalwar S, Guillozet-Bongaarts AL, Berry RW, Binder LI (2006) Formation of phospho-SAPK/JNK granules in the hippocampus is an early event in Alzheimer disease. J Neuropathol Exp Neurol 65:455-464.
Lekander M, von Essen J, Schultzberg M, Andreasson AN, Garlind A, Hansson LO, Nilsson LG (2011) Cytokines and memory across the mature life span of women. Scand J Psychol 52:229-235.
Li CY, Zhang J, Li JP, Su HM, Lin Q, Li B (2012) Research on chemical constituents and biological activities of rhizoma of Panax japonicus. Guid J Tradit Chin Med Pharm 4:68-71.
Li YG, Ji DF, Zhong S, Shi LG, Hu GY, Chen S (2010) Protect against alcohol-induced hepatic injury in mice by up-regulating the expression of GPX3, SOD1 and SOD3. Alcohol Alcohol 45:320.
Liu B, Hong JS (2003) Role of microglia in inflammation-mediated neurodegenerative diseases: mechanisms and strategies for therapeutic intervention. J Pharmacol Exp Ther 304:1-7.
Liu H, Wang J, Wang J, Wang P, Xue Y (2015a) Paeoniflorin attenuates Abeta1-42-induced inflammation and chemotaxis of microglia in vitro and inhibits NF-kappaB- and VEGF/Flt-1 signaling pathways. Brain Res 1618:149-158.
Liu H, Deng Y, Gao J, Liu Y, Li W, Shi J, Gong Q (2015b) Sodium hydrosulfide attenuates beta-amyloid-induced cognitive de ficits and neuroinflammation viamodulation of MAPK/NF-κB pathway in rats. Curr Alzheimer Res 12:673-83.
Lu D, Zhu LH, Shu XM, Zhang CJ, Zhao JY, Qi RB, Wang HD, Lu DX (2015)Ginsenoside Rg1 relieves tert-Butyl hydroperoxide-induced cell impairment in mouse microglial BV2 cells. J Asian Nat Prod Res 17:930-945.
Lu MJ, Zhu Y, Sun J, Yang XR (2011) Microglia mediates inflammation injury in mouse models of Parkinson’s disease. Zhongguo Zuzhi Gongcheng Yanjiu 15:1945-1948.
Lukiw WJ (2012) NF-kappaB-regulated, proin flammatory miRNAs in Alzheimer’s disease. Alzheimers Res Ther 4:47.
Lynch MA (2010) Age-related neuroin flammatory changes negatively impact on neuronal function. Front Aging Neurosci 1:6.
McGeer PL, McGeer EG (2004) inflammation and the degenerative diseases of aging. Ann N Y Acad Sci U S A 1035:104-116.
Miller KR, Streit WJ (2007) The effects of aging, injury and disease on microglial function: a case for cellular senescence. Neuron Glia Biol 3:245-253.
Ou YL, Xiang DY, Wu X, Xiang DX (2010) Chemical constituents and pharmacological actions research of Panax japonicus. Zhongcaoyao 6:1023-1027.Ownby RL (2010) Neuroinflammation and cognitive aging. Curr Psychiatry Rep 12:39-45.
Park WH, Kang S, Piao Y, Pak CJ, Oh MS, Kim J, Kang MS, Pak YK (2015)Ethanol extract of Bupleurum falcatum and saikosaponins inhibit neuroinflammation via inhibition of NF-κB. J Ethnopharmacol 174:37-44.
Petrov D, Luque M, Pedros I, Ettcheto M, Abad S, Pallas M, Verdaguer E,Auladell C, Folch J, Camins A (2015) Evaluation of the role of jnk1 in the hippocampus in an experimental model of familial Alzheimer’s disease.Mol Neurobiol 53:6183-6193.
Pierre WC, Smith PL, Londono I, Chemtob S, Mallard C, Lodygensky GA(2016) Neonatal microglia: the cornerstone of brain fate. Brain Behav Immun doi: 10.1016/j.bbi.2016.08.018.
Pizza V, Agresta A, D’Acunto CW, Festa M, Capasso A (2011) Neuroinflammation and ageing: current theories and an overview of the data. Rev Recent Clin Trials 6:189-203.
Qian LN, Qian LN, Chen P, Li XL, Xiao YY (2008) Study on the antifatigue function of total saponins from Panax japonicus. Zhongguo Yiyuan Yaoxue Zazhi 15:1238-1240.
Quan QK, Li X, Yuan HF, Wang Y, Liu WL (2016) Ginsenoside Rg1 inhibits high-voltage-activated calcium channel currents in hippocampal neurons of beta-amyloid peptide-exposed rat brain slices. Chin J Integr Med doi:10.1007/s11655-015-2301-4.
Shef field LG, Berman NEJ (1998) Microglial Expression of MHC Class II increases in normal aging of nonhuman primates. Neurobiol Aging 19:47-55.
Sparkman NL, Johnson RW (2008) Neuroinflammation associated with aging sensitizes the brain to the effects of infection or stress. Neuroimmunomodulation 15:323-330.
Sun H, Liang R, Yang B, Zhou Y, Liu M, Fang F, Ding J, Fan Y, Hu G (2016)Aquaporin-4 mediates communication between astrocyte and microglia:implications of neuroinflammation in experimental Parkinson’s disease.Neuroscience 317:65-75.
Waetzig V, Czeloth K, Hidding U, Mielke K, Kanzow M, Brecht S, Goetz M, Lucius R, Herdegen T, Hanisch UK (2005) c-Jun N-terminal kinases(JNKs) mediate pro-inflammatory actions of microglia. Glia 50:235-246.
Wan F, Lenardo MJ (2010) The nuclear signaling of NF-κB – current knowledge, new insights, and future perspectives. Cell Res 20:24-33.
Wang S, Zhang C, Sheng X, Zhang X, Wang B, Zhang G (2014) Peripheral expression of MAPK pathways in Alzheimer’s and Parkinson’s diseases. J Clin Neurosci 21:810-814.
Wang T, Di G, Yang L, Dun Y, Sun Z, Wan J, Peng B, Liu C, Xiong G, Zhang C, Yuan D (2015) Saponins from Panax japonicus attenuate D-galactose-induced cognitive impairment through its anti-oxidative and anti-apoptotic effects in rats. J Pharm Pharmacol 67:1284-1296.
Westman G, Berglund D, Widen J, Ingelsson M, Korsgren O, Lannfelt L,Sehlin D, Lidehall AK, Eriksson BM (2014) Increased inflammatory response in cytomegalovirus seropositive patients with Alzheimer’s disease.PLoS One 9:e96779.
Yamahara J, Kubomura Y, Miki K, Fujimura H (1987) Anti-ulcer action of Panax japonicus rhizome. J Ethnopharmacol 19:95-101.
Zhang X, Luhrs KJ, RyffKA, Malik WT, Driscoll MJ, Culver B (2009) Suppression of nuclear factor kappa B ameliorates astrogliosis but not amyloid burden in APPswe/PS1dE9 mice. Neuroscience 161:53-58.
Zou K, Zhu S, Tohda C, Cai S, Komatsu K (2002) Dammarane-type triterpene saponins from Panax japonicus. J Nat Prod 65:346-351.

- 中国神经再生研究(英文版)的其它文章
- The role of general anesthetics and the mechanisms of hippocampal and extra-hippocampal dysfunctions in the genesis of postoperative cognitive dysfunction
- Activation of the Akt/mTOR signaling pathway: a potential response to long-term neuronal loss in the hippocampus after sepsis
- Taking out the garbage: cathepsin D and calcineurin in neurodegeneration
- MicroRNAs as diagnostic markers and therapeutic targets for traumatic brain injury
- Interferon regulatory factor 2 binding protein 2: a new player of the innate immune response for stroke recovery
- Endogenous retinal neural stem cell reprogramming for neuronal regeneration