
阳春砂种子休眠与萌发过程中同工酶活性变化的研究
2018-01-24 10:44,,,
种子 2017年11期
, , ,
(广州中医药大学中药学院, 广东 广州 510006)
阳春砂(AmomumvillosumLour.)为姜科豆蔻属植物,其干燥成熟果实作砂仁药用[1]。砂仁具有化湿开胃、温脾止泻、理气安胎的功效[1],为中医治疗肠胃疾病的常用药材,主要产于广东、广西、云南、福建、海南等省区,是我国著名的四大南药之一。春砂仁种皮坚硬,具种皮障碍休眠和轻度胚休眠的特性,种子出芽慢且出芽不整齐。近年来,众多学者多侧重于研究筛选快速打破阳春砂种子休眠、促进种子萌发的方法[2-5],而有关阳春砂种子休眠与萌发过程中的生理特性研究却鲜见报道。种子的休眠与萌发是一个相互影响和制约的生理代谢过程,是在遗传、物质代谢、激素平衡、信号传导、二次休眠等因素的共同作用下完成的[6],种子萌发发育时所需的营养物质和能量主要来源于蛋白质、淀粉、脂肪等主要贮藏物质,这些贮藏物质在提供营养的同时,蛋白质酶、淀粉酶、脂肪酶及其他相关的物质代谢酶也在相应地发生变化。种子萌发是通过种子内几乎所有酶系统活性增加和新酶系统的出现为征兆的[7]。
同工酶技术广泛应用于植物、动物、微生物、农业及医学等领域的物种遗传变异、鉴定及生长发育过程等方面[8]。近年来,也应用于种子休眠与萌发研究中,并发现过氧化物酶(POD)、酯酶(EST)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)等与其有关。目前有关阳春砂种子休眠和萌发过程同工酶的研究尚无相关报道。本试验针对阳春砂种子具有休眠特性、萌发困难等特点, 开展其休眠和萌发过程中同工酶(过氧化物酶、超氧化物歧化酶、过氧化氢酶和酯酶)活性及同工酶谱的研究, 探讨种子萌发过程中同工酶活性的变化,为阳春砂种子休眠与萌发过程生理调控机制的探讨提供科学依据。
1 材料、仪器与试剂
1.1 材 料
分别于2013年和2014年,采摘春砂仁道地产区完全成熟的新鲜果实,剥弃果皮,剥开种子团,去除假种皮与杂质,置阴凉通风处,摊晾至干,装于密封袋内置4 ℃贮存,备用。
1.2 仪器与试剂
高速低温冷冻离心机(北京普析通用仪器有限责任公司);移液枪(国产DRAGON);Mini-垂直型电泳仪(美国Bio-Rad公司);POD活性测定试剂盒(南京建材有限公司);SOD活性测定试剂盒(南京建材有限公司);丙烯酰胺-甲叉丙烯酰胺(Arc-Bis)、 过硫酸铵(AP)、N,N,N′,N′-四甲基乙二胺(TEMED)、三羟甲基氨基甲烷(Tris)、甘氨酸,均购自美国Bio-Rad公司;其余均为国产分析纯。
2 方 法
2.1 种子的引发处理及发芽试验
取阳春砂种子适量,置烧杯中,用适量浓H2SO4浸泡3 min后,倾去浓H2SO4,用蒸馏水冲洗至pH呈中性,将种子置纱网上揉搓至有香气后,用400 mg/L的赤霉素溶液低温浸种72 h,倾去赤霉素溶液,用蒸馏水冲洗3次,然后将种子置于以纱布作芽床的培养皿中,于25~28 ℃光照条件下培养,分别于浸种前、浸种后、培养10 d、种子露白(培养大约30 d)、胚芽长至1 cm时(培养大约40 d)取样,用密封袋保存,置-80 ℃贮存,备用。
2.2 阳春砂种子POD、SOD同工酶活性的测定
分别取-80 ℃下的各样品适量,液氮研磨至粉末,参照试剂盒的方法测定各样品的POD及SOD同工酶的活性,记录并绘制曲线图。
2.3 酶液的制备
分别称取-80 ℃下的各样品1 g,经液氮研磨至粉末,按1∶2的比例加入0.1 mol/L的磷酸缓冲液(pH=7.5)2 mL冰浴研磨成浆,转移至离心管,4 ℃下、10 000 r/min离心15 min,吸取上清液,-20 ℃贮存备用。超氧化物歧化酶(SOD)和过氧化氢酶(CAT) 同工酶提取时需用0.1 mol/L磷酸缓冲液(pH=7.5,含5 mmol/L DTT和5% PVP)。
2.4 阳春砂种子POD、SOD、CAT及EST同工酶电泳
对提取到的酶液进行Native-PAGE电泳,浓缩胶采用4%、分离胶采用7.5%,点样量为20μL/样孔。浓缩胶采用80 V电压,待样品在浓缩胶压成直线进入分离胶后,120 V电泳,直至溴酚蓝指示带距离底部1~1.5 cm处停止。再采用联苯胺染色法、NBT染色法、铁氰化钾染色法和坚牢蓝染色法分别对POD、SOD、CAT及EST同工酶染色至条带清晰,并拍照记录。
3 结果与分析
3.1 阳春砂种子休眠与萌发过程POD、SOD同工酶活性变化趋势
结果见图1和图2。从图中可看出,阳春砂种子浸种后POD同工酶酶活性急速下降,随后在种子萌动至露白前呈缓慢上升趋势,而在露白至长出胚芽期间活性又急剧上升。阳春砂种子在休眠与萌发过程中SOD酶的活性变化则是:种子经激素浸种处理后酶活性有所下降,后整体略呈直线上升的趋势。

图3 阳春砂种子休眠与萌发过程中POD同工酶图谱
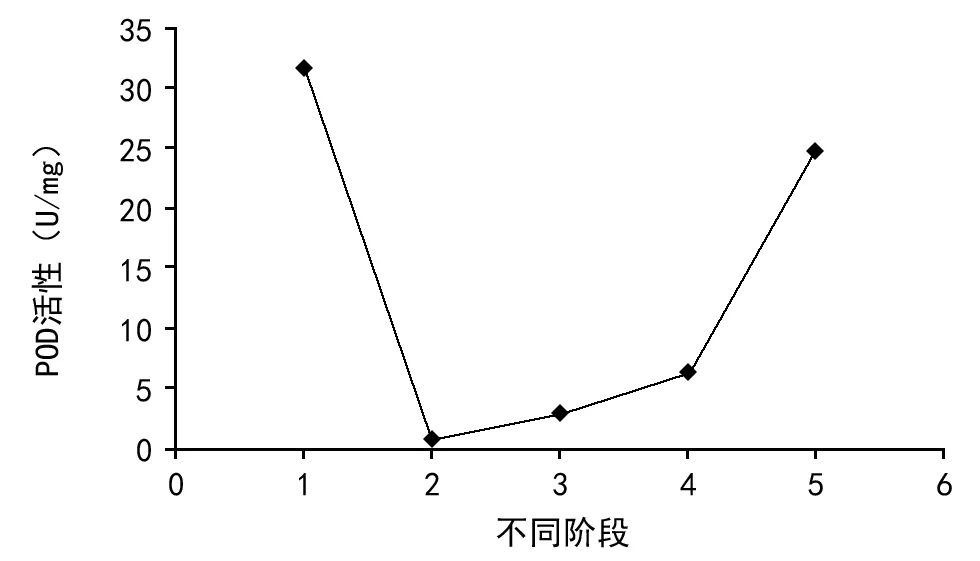
注:1为浸种前;2为浸种后;3为培养10 d;4为露白;5为胚芽1 cm。下同。图1 阳春砂种子休眠与萌发过程中POD同工酶活性变化

图2 阳春砂种子休眠与萌发过程中SOD同工酶活性变化
3.2 阳春砂种子休眠与萌发过程同工酶谱带研究
3.2.1 POD同工酶谱
阳春砂种子休眠与萌发过程的POD同工酶出现谱带可划分为Ⅰ、Ⅱ、Ⅲ、Ⅳ 4个区带(见图3),其中干种子的POD同工酶谱带共有5条,均分布于Ⅱ区;浸种后及培养10 d种子的POD同工酶谱均在Ⅲ和Ⅳ区各新增1条条带,Ⅱ区则均减为1条,且颜色较浅,显示其活性较弱;种子露白及胚芽长1 cm时,Ⅲ和Ⅳ区的条带数保持不变,后者在Ⅰ区新增了1条新的条带,而二者在Ⅱ区的条带数则递增,颜色呈逐渐加深的趋势,显示其活性增强,表明阳春砂种子休眠与萌发过程中POD同工酶活性呈先减弱后不断增强的趋势,期间POD同工酶的种类也发生了相应的变化。
3.2.2 SOD同工酶谱
如图4所示,阳春砂种子浸种前后SOD同工酶的谱带均较亮,浸种后较浸种前减弱,说明浸种后种子代谢活动有所减弱,而培养10 d后种子至种子露白到长出胚芽期间,SOD同工酶谱带亮度又有所增强,表明其活性呈先减弱后不断增强的趋势,这与SOD活性测定结果一致。
3.2.3 CAT同工酶谱
如图5所示,浸种前后及培养10 d的阳春砂种子的CAT同工酶谱带均只有1条,且颜色深浅变化不明显。露白及长出胚芽时种子CAT同工酶谱带均变为2条,且条带颜色不断加深,由此表明,阳春砂种子中CAT同工酶在萌发前期相对稳定,到露白期活性开始增强,酶种类也在增加。
3.2.4 EST同工酶谱
如图6所示,阳春砂种子休眠与萌发过程中EST同工酶一共出现4条条带,浸种后种子的EST同工酶谱带数较干种子减少1条,且条带色泽减弱,表明浸种后的阳春砂种子EST同工酶的活性减弱,培养10 d及露白的种子EST同工酶合成增加,并出现新酶带Ⅱ、Ⅲ和Ⅳ,且同工酶活力在种子露白期达到高峰,而长出胚芽的种子条带数又减至2条。由此表明,阳春砂休眠与萌发过程的EST同工酶的活性呈先减弱后增强再减弱的变化趋势,种子露白时活性达到最强。
4 讨 论
研究筛选了阳春砂种子POD、SOD、CAT及EST同工酶聚丙烯酰胺凝胶电泳条件:同工酶酶液提取方法筛选结果显示,提取液体积比1∶2时最佳,不同提取液对POD及EST同工酶的提取效果无显著差异,SOD及CAT同工酶需用0.1 mol/L的磷酸缓冲液(pH=7.5,含5 mmol/L DTT和5% PVP)防止氧化进行提取。单向垂直电泳条件的筛选结果显示,胶浓度、点样量及电压均对最后的结果影响不大。因此,阳春砂种子同工酶电泳的关键在于酶液的制备,确保提取过程中酶的活性对后续的实验至关重要。

图4 阳春砂种子休眠与萌发过程中SOD同工酶图谱

图5 阳春砂种子休眠与萌发过程中CAT同工酶图谱

图6 阳春砂种子休眠与萌发过程中EST同工酶图谱
POD同工酶与植物体内多种代谢活动有关,参与细胞分化和一些生理活性物质的合成与降解,在植物抗逆境胁迫中,与SOD、CAT一样都是非常关键的酶,能够消除氧自由基对细胞的损伤;EST也参与植物体生长发育过程中的多种生理过程[9-13]。研究结果显示,阳春砂种子从开始培养到长出胚芽阶段,POD、SOD、CAT同工酶活性基本呈上升趋势,并且在露白到长出胚芽阶段急速上升,说明种子开始萌发时,整个生理代谢活跃起来,尤其是种子开始露白,也即真正萌发的阶段,在萌发中后期,酶活性急剧增加也在一定程度上说明了种子在萌发时糖类和脂类的代谢产生了大量的有害产物(如脂质过氧化物),会对细胞、生物体造成伤害,因此,这些有害产物就会诱导防御系统相关酶(POD、SOD、CAT)的合成增加,来清除有害产物,从而促进种子的萌发;同时Roberts等也曾指出,POD酶活性的增加导致HMP途径的加速,可能是萌发的重要原因[14],与本实验的结果一致;另一方面,种子由干燥休眠状态到吸胀状态,POD、SOD活性减小,表达量很低或几乎没有表达,说明干种子细胞膜受到活性氧自由基的影响较大,种子浸种后代谢减弱,营养物质被贮藏不被代谢,为萌发作准备,因此所产生的有害产物较少。EST同工酶浸种后a、b条带颜色均变浅,后又开始加深,并且在露白时期出现了新的条带,说明种子在浸种时代谢减弱,而萌发后代谢活跃,条带颜色加深并出现新的条带,与POD、SOD及CAT同工酶结果一致。阳春砂种子休眠与萌发过程中POD、SOD、CAT、EST同工酶条带的变化,在一定程度上表明了这些同工酶基因的表达导致了阳春砂种子休眠的打破和最终的萌发。
[1]国家药典委员会.中国药典(一部)[M].北京:中国医药科技出版社,2010:236.
[2]刘艳,杨锦芬,何瑞,等.不同发芽条件对阳春砂种子发芽的影响[J].广州中医药大学学报,2010,27(4):399-402.
[3]张丹雁,欧阳霄妮,徐志东,等.不同浸种处理对阳春砂种子发芽的影响[J].广州中医药大学学报,2010,27(1):62-64.
[4]阮英恒,刘军民,孙林霞,等.道地产区阳春砂种子的品质检验及其贮藏特性的研究[J].中药新药与临床药理,2014,25(2):222-226.
[5]于志忱,韩德聪,黄庆昌,等.春砂仁(AmomumvillosumLour.)种子发芽的试验研究[J].中山大学学报,1981(1):102-108.
[6]潘琳,徐程扬.种子休眠与萌发过程的生理调控机理[J].种子,2010,29(6):42-46.
[7]潘瑞炽.植物生理学[M].北京:高等教育出版社,2001.
[8]Kermode AR,Finch-Savage BE.Desiccation sensitivity in orthodox and recalc-itrant seeds in relation to development.In:Black M,Pritchard HW,eds.Desiccation and Survival in Plants,Drying Without Drying.Wallingford:CAB International,2002:149-184.
[9]葛超,赵成昊,胡凯,等.刺五加种子休眠解除过程中过氧化物酶和酯酶同工酶的变化[J].种子,2010,29(1):50-52.
[10]周化斌,徐根娣,刘鹏,等.大豆种子萌发中过氧化物酶同工酶的动态研究[J].种子,2002,21(1):9-11.
[11]刘国勇,彭丽娟,陈发菊.香果树种子萌发过程中酯酶和过氧化物酶同工酶分析[J].安徽农学通报,2009,15(6):30-31.
[12]梁宏伟,刘姝,陈发菊.银鹊树种子萌发过程中的过氧化物酶和酯酶同工酶的变化[J].种子,2006,25(5):38-40.
[13]伍贤进,宋松泉,田向荣,等.玉米种子吸胀萌发过程中抗氧化酶活性的变化[J].吉林农业大学学报,2004,26(1):6-9.
[14]Roberts C H.Oxidative Process and the control of seed germination in Seed Ecology[M].London:Butter-Worth,1973:189.
猜你喜欢
青年文学家(2021年4期)2021-03-18
防护林科技(2020年11期)2020-12-30
农民致富之友(2020年19期)2020-07-18
微生物学杂志(2020年6期)2020-03-05
家庭百事通·健康一点通(2019年11期)2019-09-26
中国食品学报(2019年3期)2019-01-13
天津农业科学(2018年3期)2018-03-24
江苏农业科学(2017年12期)2017-09-16
江苏农业科学(2016年8期)2017-02-15
河北渔业(2015年9期)2015-09-15
