
海拔梯度对塔尔巴哈台山地鸭茅群落物种组成及多样性影响
2018-03-09 09:12何盘星张鲜花朱进忠
草业科学 2018年2期
何盘星,张鲜花,朱进忠
(新疆农业大学草业与环境科学学院,新疆 乌鲁木齐 830000)
物种多样性是维持草地生态系统多样性和草地持续稳定利用的前提[1-2]。目前,人类过度利用导致天然草地植物种群栖息地破碎与丧失,从而出现不同程度的退化和沙化等,同时有毒有害植物侵入,导致适合当地生存的一些优势物种逐渐消失,物种多样性丧失,草地生态系统遭到破坏,草地经济和生态服务价值降低[3]。因此,为维护草地生态系统的稳定,草地植物群落物种多样性的研究显得尤为重要。已有研究证实,不同的生态因子会对物种多样性产生一定的影响,如海拔梯度就是一个综合的复杂的生态因子,与温度、水分、光照及地形等条件的改变有紧密相关性[4-7]。有研究发现,水热条件的变化导致地表异质性,进而整体改变了群落的外貌、结构特征及演替方向等[8]。
鸭茅(Dactylisglomerata)是一种重要的优良冷季型牧草,已作为建植高产优质栽培草地的主要牧草广泛种植[9-10]。鸭茅在新疆的天山、阿尔泰山、准噶尔西部山地等广泛分布,是山地草甸的主要优势种,占据海拔1 300-2 400 m,生境多样。近年来,由于天然草地不同程度的退化,以鸭茅为主的天然草地植物群落物种多样性逐渐降低,优良牧草数量大幅度减少,优良基因性状衰退,有的甚至濒临灭绝,直接导致草地生产力下降。本研究以塔尔巴哈台山地未受或很少受到人类活动影响的鸭茅种群集中分布区域的草地为研究对象,分析海拔梯度变化对鸭茅群落物种组成及群落数量特征的影响,探讨物种多样性水平及影响因素,以及多样性与海拔梯度等环境因素的相互关系,为合理利用与保护鸭茅种质资源、物种多样性提供科学依据。
1 材料与方法
1.1 研究区概况
塔尔巴哈台山地位于新疆西北部,属于准噶尔西部山地部分,在我国境内分布在塔城盆地北缘,地处47°03′37.3″-47°06′29.2″ N,82°59′33.9″-83°00′35.0″ E,海拔1 200-1 900 m地段(图1)。气候为中温带大陆性气候,夏季炎热短暂,冬季漫长酷冷。塔尔巴哈台山较高大的山体拦截了来自北冰洋和大西洋的湿润水汽,在山区的迎风坡形成丰沛的降水,年降水量在450 mm以上,年平均气温为5.5 ℃,极端最高气温38.5 ℃,最低气温-35.4 ℃。1 300-1 900 m处实地测量土壤含水量在26%~47%(图2)。土壤类型为山地黑钙土。草地类型主要为山地草甸草地,植物主要以禾草和杂类草为主,优良牧草种类多,禾本科植物占48.4%以上。研究区概况如表1所列。

图1 研究区采样点分布图Fig. 1 Sampling location distribution map
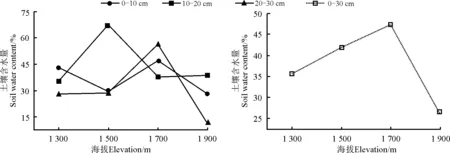
图2 土壤含水量的趋势图Fig. 2 Trend chart of soil water content
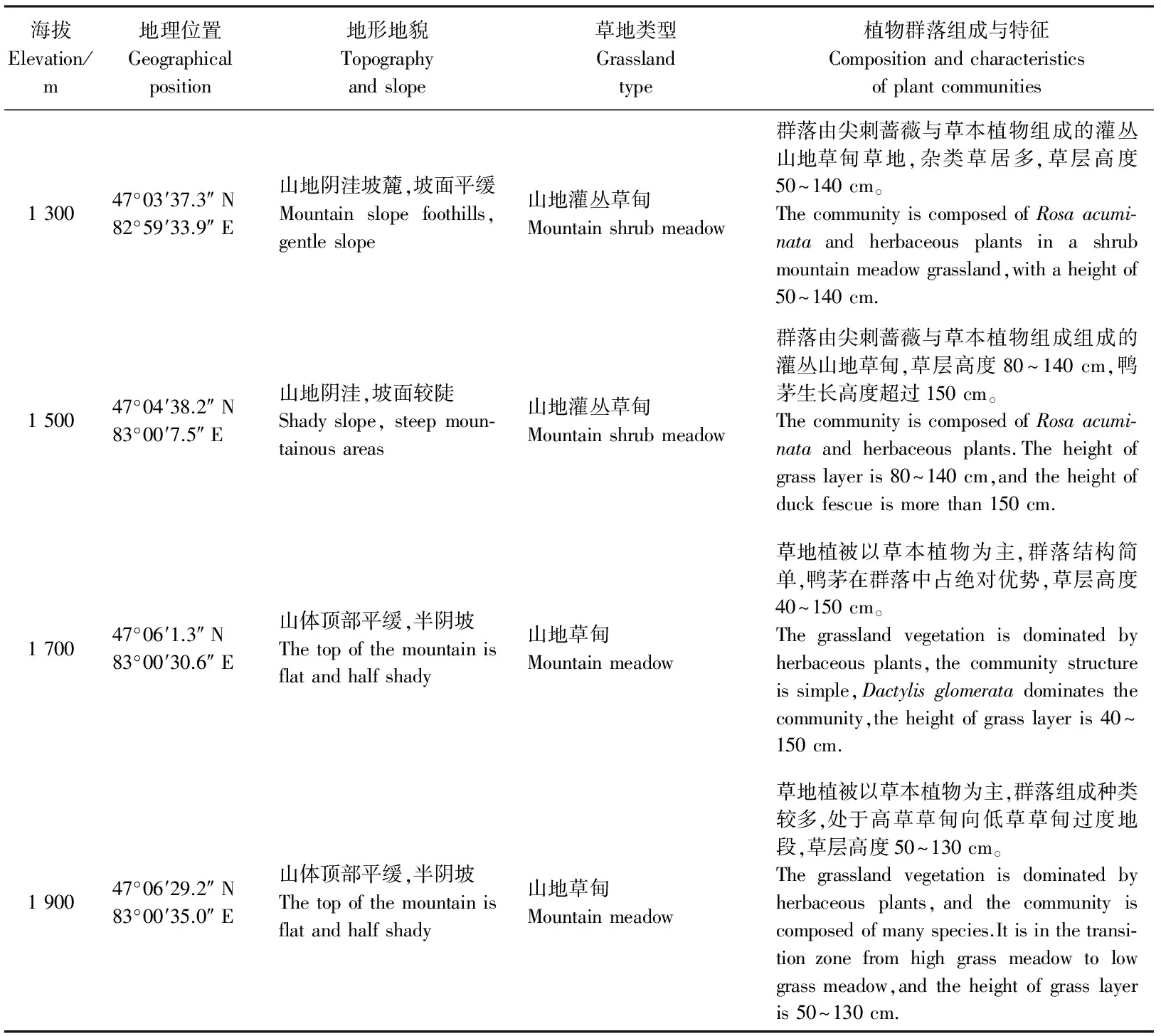
海拔Elevation/m地理位置Geographicalposition地形地貌Topographyandslope草地类型Grasslandtype植物群落组成与特征Compositionandcharacteristicsofplantcommunities130047°03′37.3″N82°59′33.9″E山地阴洼坡麓,坡面平缓Mountainslopefoothills,gentleslope山地灌丛草甸Mountainshrubmeadow群落由尖刺蔷薇与草本植物组成的灌丛山地草甸草地,杂类草居多,草层高度50~140cm。ThecommunityiscomposedofRosaacumi⁃nataandherbaceousplantsinashrubmountainmeadowgrassland,withaheightof50~140cm.150047°04′38.2″N83°00′7.5″E山地阴洼,坡面较陡Shadyslope,steepmoun⁃tainousareas山地灌丛草甸Mountainshrubmeadow群落由尖刺蔷薇与草本植物组成组成的灌丛山地草甸,草层高度80~140cm,鸭茅生长高度超过150cm。ThecommunityiscomposedofRosaacumi⁃nataandherbaceousplants.Theheightofgrasslayeris80~140cm,andtheheightofduckfescueismorethan150cm.170047°06′1.3″N83°00′30.6″E山体顶部平缓,半阴坡Thetopofthemountainisflatandhalfshady山地草甸Mountainmeadow草地植被以草本植物为主,群落结构简单,鸭茅在群落中占绝对优势,草层高度40~150cm。Thegrasslandvegetationisdominatedbyherbaceousplants,thecommunitystructureissimple,Dactylisglomeratadominatesthecommunity,theheightofgrasslayeris40~150cm.190047°06′29.2″N83°00′35.0″E山体顶部平缓,半阴坡Thetopofthemountainisflatandhalfshady山地草甸Mountainmeadow草地植被以草本植物为主,群落组成种类较多,处于高草草甸向低草草甸过度地段,草层高度50~130cm。Thegrasslandvegetationisdominatedbyherbaceousplants,andthecommunityiscomposedofmanyspecies.Itisinthetransi⁃tionzonefromhighgrassmeadowtolowgrassmeadow,andtheheightofgrasslayeris50~130cm.
1.2 样地设置
在海拔1 300 m-1 900 m鸭茅集中分布区域,按照每隔200 m海拔梯度设置一个调查样点,每个样点设置3个样地,每个样地面积20 m×20 m,每个样地内按照十字交叉法每隔5 m设置1 m×1 m样方共9个,记录测定物种种类、高度、密度、盖度和地上生物量。
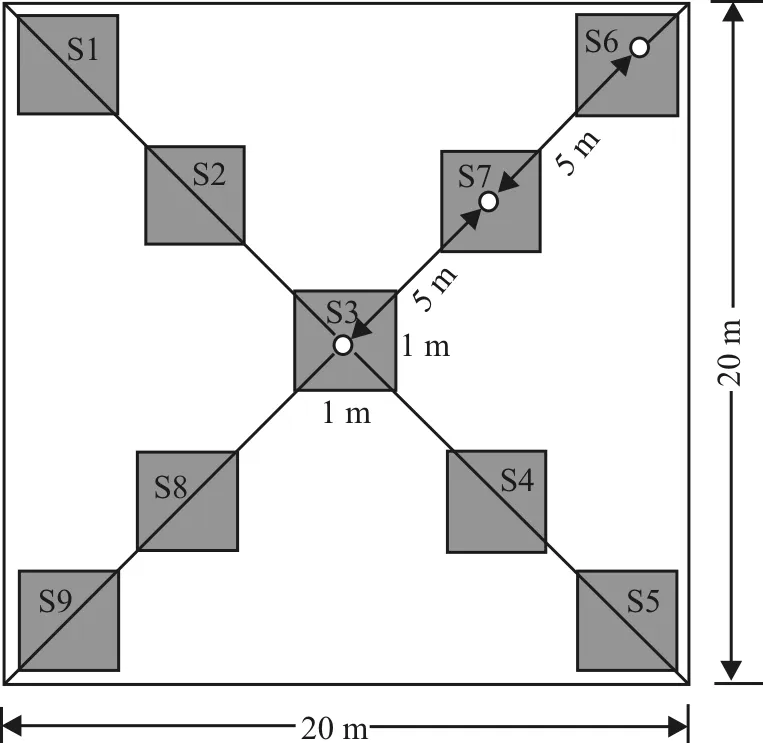
图3 样方设计示意图 Fig. 3 Design schematic of square
1.3 测定方法
1.3.1群落数量特征测定
1)高度(height):测定样方内所有植物的自然生长高度。
2)盖度(coverage):采用针刺法测定;
3)密度(density):分别测量每种植物的株丛数。
4)地上生物量(aboveground biomass):齐地面刈割测定每种植物鲜重。
5)植物重要值(important value,IV):
重要值=(相对高度+相对盖度+相对密度+相对生物量)/4。
1.3.2物种多样性测度 为分析不同海拔梯度下鸭茅群落物种多样性的空间分布格局,采用马克平等[11]生物多样性两个空间尺度的测度方法,对α多样性和β多样性进行分析。α多样性主要是关注局域生境下的物种数目,反映生境内的多样性;β多样性指沿环境梯度不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率,反映生境间的多样性。
(1)α多样性
Shannon-Wiener指数:
Simpson指数:
Pielou指数:
Esw=H/lnS。
校正Pielou均匀度指数:
Esi=(S-1)/(1-1/S)。
Margalef指数:
R=(S-1)/lnN。
物种丰富度:
N=S。
式中:Pi=Ni/N,Pi为第i个物种的相对重要值,Ni为各物种的重要值,N为全部物种个数之和,S为样方中物种总数。
(2)β多样性
Sφrensen β多样性指数(s):
Cs=1-2c/(a+b)。
Jaccard β多样性指数(j):
Cj=1-c/(a+b+c)。
式中:a、b为两群落的所有物种数,c为群落间共有的物种数量。
1.3.3数据处理 利用SPSS 17.0软件进行单因子方差分析和LSD显著性检验,并进行多样性指数间的相关性分析;采用Origin 8.0软件制图。
2 结果与分析
2.1 海拔梯度对鸭茅群落物种组成影响
根据对 4个不同海拔地段的植物组成数据资料的统计,发现塔尔巴哈台山地鸭茅群落生境中约有植物 52种,隶属于17科(表2)。其中,以菊科(Compositae)植物最多,为12种,其次为禾本科(Gramineae),8种,唇形科、十字花科和蔷薇科均为4种,豆科、牻牛儿苗科和伞形科均为3种,其余12种分属于8科。 其中,海拔1 900 m处植物种类数最多,为32种,海拔1 700 m处植物种类组成较为简单,海拔1 500 m处灌木类植物较多,灌丛在群落中所占比重较大,而海拔1 300 m处由于处于山坡坡脚处,灌木较海拔1 500 m处少,草本植物所占比重较大。
2.2 海拔梯度对鸭茅群落数量特征影响
塔尔巴哈台山的鸭茅群落高度、盖度、密度及地上生物量随海拔梯度变化呈现出一定的差异性,但未表现出一定的规律性(表3)。

表2 不同海拔地段鸭茅群落植物种类重要值Table 2 Dactylis glomerata community important value of plant species in different elevations
续表2
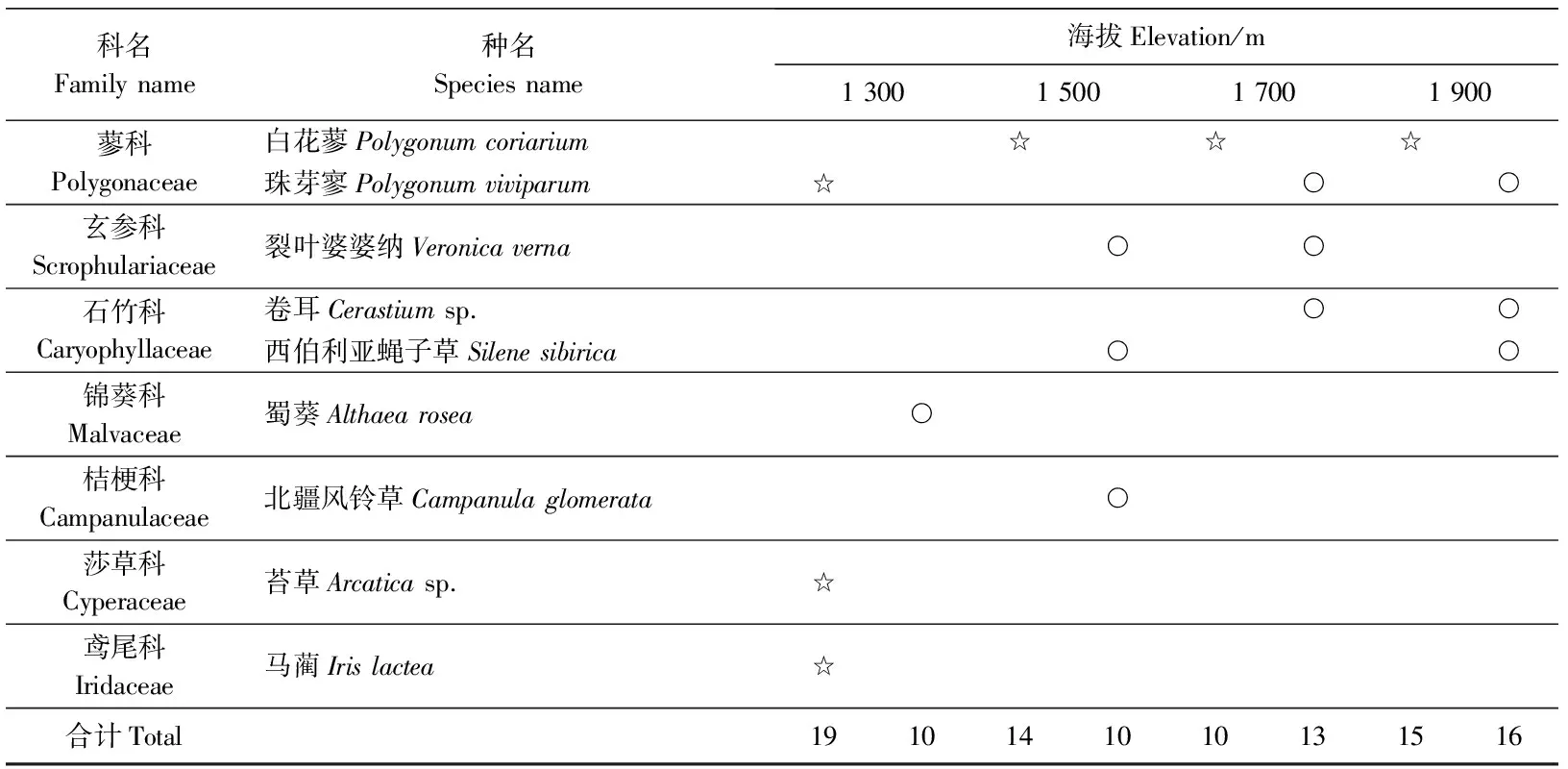
科名Familyname种名Speciesname海拔Elevation/m1300150017001900蓼科Polygonaceae白花蓼Polygonumcoriarium☆☆☆珠芽寥Polygonumviviparum☆○○玄参科Scrophulariaceae裂叶婆婆纳Veronicaverna○○石竹科Caryophyllaceae卷耳Cerastiumsp.○○西伯利亚蝇子草Silenesibirica○○锦葵科Malvaceae蜀葵Althaearosea○桔梗科Campanulaceae北疆风铃草Campanulaglomerata○莎草科Cyperaceae苔草Arcaticasp.☆鸢尾科Iridaceae马蔺Irislactea☆合计Total1910141010131516
☆代表重要值大于0.01,是群落中的常见物种;○代表重要值小于0.01,是群落中的偶见种。
☆, the common species in the community, and the important value is greater than 0.01; ○, occasional species in the community, and the important value is less than 0.01.

表3 不同海拔鸭茅群落的数量特征Table 3 Community numerical characteristics of D. glomerata in different elevations
同列不同小写字母表示差异显著(P<0.05)。
Different lowercase letters within the same column indicate significant difference at the 0.05 level.
随海拔升高,群落高度呈现出先升高后降低的趋势,海拔1 500 m处显著高于其他海拔地段(P<0.05),海拔1 900 m处最低。随海拔梯度的变化,群落盖度在各海拔之间未呈现显著差异(P>0.05),且群落盖度均在85%以上。海拔1 300 m处群落密度显著高于其他海拔地段,且随海拔升高呈先降低后升高再降低的趋势,而在海拔1 500 m处最低,主要原因是灌丛在群落中占有的比重较大。群落生物量与群落密度呈现出一致的趋势,在海拔1 700 m处达到最大值,为1 209.82 g·m-2,但各海拔地段未呈现显著差异(P>0.05)。
鸭茅在群落中所占比重随海拔升高逐渐增加,且在海拔1 700 m处达到峰值49.2%,随后在1 900 m处海拔略有减少。
2.3 海拔梯度对鸭茅群落物种多样性的影响
2.3.1α多样性 α多样性指数随海拔变化表现出一定的规律性,如Shannon-Wiener指数和丰富度指数随海拔升高表现出先减少后增加趋势,呈现 “V”字型,在海拔1 700 m处显著低于其他海拔地段(P<0.05);而Margalef指数、Simpson指数却随海拔升高呈先增加后降低再增加的趋势,Margalef指数在海拔1 700 m处最低,Simpson指数在各海拔地段显示无显著差异(P>0.05); Pielou均匀度指数未呈现出一定的规律性,当海拔上升到1 900 m时,均匀度指数达到最小;校正后均匀度指数与海拔上升呈相反趋势(图2)。
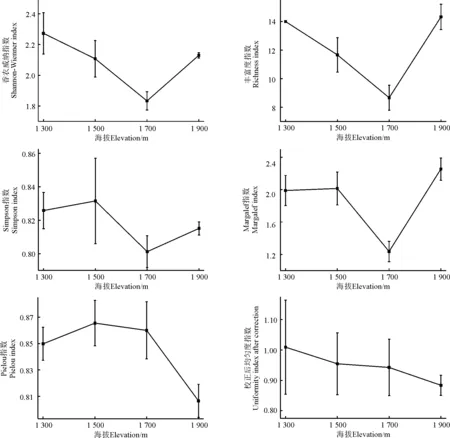
图4 不同海拔鸭茅群落多样性指数Fig. 4 α Diversity index of D. glomerata in different elevation
2.3.2鸭茅群落物种β多样性指数 一般情况下,控制β多样性因子为土壤、地貌及干扰等。Sφrensen β多样性指数结果显示,1 300~1 900 m的Sφrensen β多样性指数介于0.429~0.778,1 300 m与1 500 m处的β多样性指数最高,说明两处的共有种较少,1 700 m与1 900 m的β多样性指数最低,说明两处的共有种相对较多,但是4个海拔之间彼此差异不大。Jaccard β多样性指数也印证了同样的结果,随海拔高度的上升,Jaccard β多样性指数依次降低(表4)。
2.4 鸭茅群落物种多样性与海拔及地上生物量的关系
研究结果显示,海拔与多样性指数之间存在不同程度的负相关关系,即海拔的升高伴随着多样性指数的减少(表5)。地上生物量与Margalef指数、Simpson指数、Shanon-Wiener指数及物种丰富度指数为负相关,与Pielou均匀度指数、校正后均匀度指数为正相关,整体而言,地上生物量与物种多样性负相关,但指数与地上生物量之间的相关性均不显著(P>0.05)。

表4 不同海拔鸭茅群落Jaccard β多样性和Sφrensen β多样性(所有物种)Table 4 Jaccard β diversity and Sφrensen β diversity of D. glomerata in different elevations
加黑数值为Jaccard β多样性指数,未加黑数值为Sφrensen β多样性指数。
Values with bocd are Jaccard β diversity index, Values without bold are Sφrensen β diversity index.
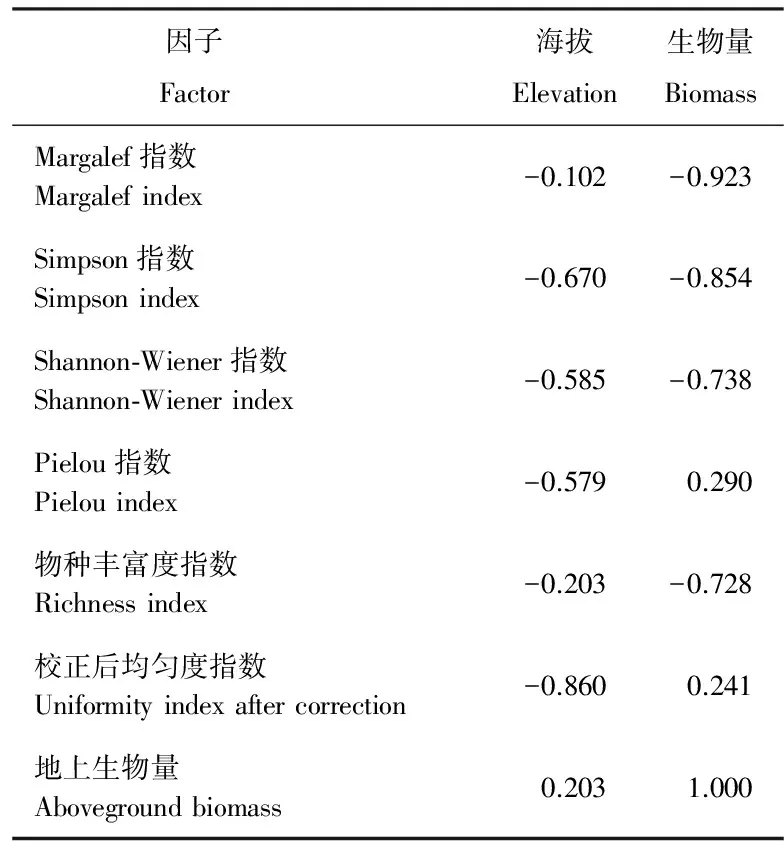
表5 物种多样性与海拔、生物量的相关性分析Table 5 Correlation analysis of species diversity with elevation and biomass
3 讨论与结论
新疆野生鸭茅种群主要分布在天山北坡、阿尔泰山及准噶尔西部山地的草甸草地,本研究选择准噶尔西部山地的塔尔巴合台山地未受人类活动干扰、保持自然状态的鸭茅群落。经研究证实,塔尔巴哈台山地是近代南北植物区系交汇的区域,具有南北植物区系共有的特征,物种丰富且生态系统复杂[12-13]。
3.1 物种组成变化
本研究针对塔尔巴哈台山天然草地鸭茅群落不同海拔地段物种组成变化的研究发现,从海拔1 300 m的山地灌丛草甸到海拔1 900 m山地草甸草地,鸭茅的重要值始终保持在28%~37%,且在各海拔地段均排在第1位,由此表明,鸭茅是该群落的优势种或建群种。而鸭茅群落的主要伴生种在组成上呈现出随海拔梯度的变化,伴随着地形、水分、坡向坡度等差异,主要伴生种由海拔1 300 m的山地灌丛草甸的蔷薇、早熟禾、千叶蓍等植物逐渐过渡到海拔1 900 m的山地草甸的草原糙苏、草原老鹳草等植物。同时,不同海拔地段共有物种数不同,海拔山地草甸1 700和海拔1 900 m共有物种数较多,而灌丛草甸海拔1 300与海拔1 500 m之间共有物种数却较少。山地草甸海拔1 700、1 900 m处均位于较为平缓的坡地,坡向一致,地形可能是造成共有物种数较多的主要原因,物种替代速率随海拔升高逐渐降低。而灌丛草甸海拔1 300 m地处山体阴坡沟谷地带,海拔1 500 m地处山体阴坡陡坡坡面,水热条件不同导致两海拔地段共有物种数较少。
综上所述,在同一性质的草地类型中,群落的物种组成变化与草地类型可能无较大关系,导致其差异最大,众多研究表明,群落物种组成变化可能是海拔、地形变化、水分、土壤理化性质、微地形等环境因子共同作用的结果[14-17]。如王继丰等[16]通过对三江平原湿地小叶章(Calamagrostisangustiflia)群落随土壤水分梯度变化的群落物种组成的研究发现,小叶章在不同生境下均为优势种,其伴生种组成分化明显,其原因可能是自身生物学特性、水分、地形及其他环境因子共同作用的结果。又如古尔班通古特沙漠中的短命植物,由于沙丘大小、坡向、坡位不同受到光照时间和强度及降水入渗的差异,导致物种组成发生波动变化[17],本研究结果与之相一致。
3.2 环境因子对物种多样性的影响
物种多样性是多种环境梯度(海拔梯度、纬度梯度、水分梯度等)综合作用的结果,其中海拔梯度是环境梯度中的主要梯度[18]。贺金生和陆伟烈[4]总结出物种多样性随海拔梯度的变化可分为5种情形,分别是:先升高后降低(中间高度膨胀型)、先降低后升高(V型)、单调降低(负相关)、单调升高(正相关)和无规律性(无关),自然界中最普遍的是中间高度膨胀型与单调降低的现象。如青海高寒草甸和滇西玉龙雪山地区发现物种多样性呈先增加后减少的趋势[19-21], 祁连山自然保护区和川西亚高山森林群落物种多样性随海拔的升高物种多样性减少[22-23]。此外,也有研究发现,岷江上游干旱河谷地段物种多样性受焚风效应的作用,随海拔的上升呈增加的趋势[24];沈泽昊等[25]发现贡嘎山海螺沟草本植物的物种多样性与海拔并无显著相关关系。本研究中,海拔1 300-1 900 m山地灌丛草甸到山地草甸的α多样性指数呈现先降低再逐渐升高的“V”格局,在海拔1 700 m处α多样性指数最低。分析其原因可能是,塔尔巴哈台山位于中纬度山地迎风坡,受到山体的抬升作用,大气降水随海拔的升高而增加,到一定程度后再减少[26-27],海拔1 700 m处恰好是塔尔巴哈台山的中山地带,土壤含水量最高,湿润的生境条件极大的满足了鸭茅的生存状况,导致海拔1 700 m处的鸭茅种群过度的对生境条件的占据,挤压了其他物种的生存空间,因而导致此处的物种丰富度较小,实地测量的土壤含水量也证实了这一点(图2)。
此外,坡度与坡向也是影响海拔1 700 m处物种丰富度较少的原因,位于山顶平缓地带,地形起伏不大,部分阴坡生长的植物消失,杨元合和朴世龙[28]研究也发现,物种丰富度与降水和温度呈显著正相关。鸭茅作为4个海拔梯度的优势种,具有较大的生态位宽度,在海拔1 700 m处的典型山地草甸的地形、水分等的综合条件无疑是鸭茅最佳的生态位生境,种群生长迅速,在群落中占绝对优势,海拔1 700 m处是 4 个海拔梯度中生境多样性最低的一个类型,造就了部分优势物种的最适生境,优势种占据一定生态位的能力,获取水分与热量的能力较强,挤压了其他物种的生存空间,导致群落偶见种和稀有种出现的几率减少。生境的异质性为不同生境要求物种的存活提供了可能,这里的结论与 “中间膨胀理论”或负相关理论并不冲突,特殊的原因是水热等环境因子的变化对群落物种多样性产生了影响。
综上所述,物种多样性随海拔梯度呈现出“V”型趋势,但各指数与海拔梯度整体上为负相关关系,这从一定程度上反映出随海拔的升高,天然草地生态系统逐渐变得更加脆弱和敏感、抗干扰或稳定性差,即使没有人类过度放牧或强烈干扰,群落组成和物种多样性也很容易发生改变[29]。因此,应采取合理有效的保护措施,尽量减少人类对天然草地的利用强度与频率,为保护好优质牧草的种质资源提供天然的庇护地,同时也可保证山地草地生态系统可持续发展和物种多样性的维持。
References:
[1] 章家恩,徐琪.现代生态学研究的几大热点问题透视.地理科学进展,1997(3):31-39.
Zhang J E,Xu Q.Perspective of hot problems in contemporary ecology.Progress in Geography,1997(3):31-39.(in Chinese)
[2] 杨利民,韩梅,周广胜,王玉辉.草地群落物种多样性维持机制的研究Ⅲ.物种分布格局.吉林农业大学学报,2002,24(1):58-61.
Yang L M,Han M,Zhou G S,Wang Y H.Studies on the maintaining mechanism of species diversity of grassland community Ⅲ.Species distribution pattern.Journal of Jilin Agricultural University,2002,24(1):58-61.(in Chinese)
[3] 刘兴元,龙瑞军,尚占环.草地生态系统服务功能及其价值评估方法研究.草业学报,2011,20(1):167-174.
Liu X Y,Long R J,Shang Z H.Evaluation method of ecological services function and their value for grassland ecosystems.Acta Prataculturae Sinica,2011,20(1):167-174.(in Chinese)
[4] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征.生态学报,1997(1):91-99.
He J S,Chen W L.A review of gradient changes in species diversity of land plant communities.Acta Ecologica Sinica,1997(1):91-99.(in Chinese)
[5] 潘红丽,李迈和,蔡小虎,吴杰,杜忠,刘兴良.海拔梯度上的植物生长与生理生态特性.生态环境学报, 2009,18(2):722-730.
Pan H L,Li M H,Cai X H,Wu J,Du Z,Liu X L.Responses of growth and ecophsiology of plants to altitude.Ecology and Environmental Sciences,2009,18(2):722-730.(in Chinese)
[6] 田青,李宗杰,王建红,宋玲玲,韩蓉,陈博.摩天岭北坡东南部不同海拔梯度草本植物群落特征.草业科学,2016,33(4):755-763.
Tian Q,Li Z J,Wang J H,Song L L,Han R,Chen B.The characteristics of herbaceous plant communities in the southeast of Motianling northern slope at different altitudes.Pratacultural Science,2016,33(4):755-763.(in Chinese)
[7] 白晓航,张金屯,曹科,王云泉,Sehrish Sadia,曹格.小五台山亚高山草甸的群落特征及物种多样性.草业科学,2016,33(12):2533-2543.
Bai X H,Zhang J T,Cao K,Wang Y Q,Sadia S,Cao G.Community characteristics and species diversity of subaipline meadows in Xiaowutai Mountain.Pratacultural Science,2016,33(12):2533-2543.(in Chinese)
[8] 乌云娜,张云飞.锡林郭勒草原植物物种多样性的水分梯度特征.内蒙古大学学报(自然版),1998(3):407-413.
Wu Y N,Zhang Y F.The characteristic of species diversity along water gradients in Xilinguole Steppe.Acta Scientiarum Naturalium Universitatis Nei Mengol,1998(3):407-413.(in Chinese)
[9] 张鲜花,朱进忠,李海琪.海拔梯度对天山北坡天然草地鸭茅种群分布格局的影响.草业科学,2016,33(12):2551-2558.
Zhang X H,Zhu J Z,Li H Q.Effects of scale on the regional distribution patterns ofDactylisglomerataat different altitudes in the northern Tianshan Mountains.Pratacultural Science,2016,33(12):2551-2558.(in Chinese)
[10] 彭燕,张新全.鸭茅种质资源多样性研究进展.植物遗传资源学报,2003,4(2):179-183.
Peng Y,Zhang X Q.Progress in studies on the genetic diversity ofDactylisglomerataL. Journal of Plant Genetic Resources,2003,4(2):179-183.(in Chinese)
[11] 马克平,黄建辉,于顺利,陈灵芝.北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数.生态学报,1995(3):268-277.
Ma K P,Huang J H,Yu S L,Chen L Z.Plant community diversity in Dongling Mountain.Beijing.China.Ⅱ.Species richness.evenness and species diversities.Acta Ecologica Sinica,1995(3):268-277.(in Chinese)
[12] 张鲜花.天山北坡东段与西段野生鸭茅的种群分布、形态特征及生理生化特性研究.乌鲁木齐:新疆农业大学博士学位论文,2015.
Zhang X H.Study on population distribution,morphological characteristics and physicological-biochemical characteristics ofDactylisglomeratain east and west section of Tianshan Mountain north slope.PhD Thesis.Urumqi:Xinjiang Agricultural University,2015.(in Chinese)
[13] 黄俊华,杨昌友,向其柏.中国萨吾尔-塔尔巴哈台山地种的地理成分分析.云南植物研究,2004,26(1):12-18.
Huang J H,Yang C Y,Xiang Q B.Geographical composition analysis of species in Saur-Tarbagatai Mountains in China.Acta Botanica Yunnanica,2004,26(1):12-18.(in Chinese)
[14] 段敏杰,高清竹,郭亚奇,万运帆,李玉娥,干珠扎布,旦久罗布,韦兰亭,西饶卓玛.藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局.草业科学,2011,28(10):1845-1850.
Duan M J,Gao Q Z,Guo Y Q,Wan Y F,Li Y E,Ganzhuzhabu,Danjiuluobu,Wei L T,Xiraozhuoma.Species diversity distribution paterrn of alpine grassland communities along an altitudinal gradient in the northern Tibet.Pratacultural Science,2011,28(10):1845-1850.(in Chinese)
[15] 张巧凤,刘桂香,于红博,包玉海.锡林郭勒草原主要植被群落变化.草业科学,2015,32(9):1381-1391.
Zhang Q F,Liu G X,Yu H B,Bao Y H.The changes of main plant communities in Xilinggol grassland.Pratacultural Science,2015,32(9):1381-1391.(in Chinese)
[16] 王继丰,韩大勇,王建波,付晓玲,朱道光,刘赢男,曹宏杰,黄庆阳,谢立红,钟海秀,隋心,倪红伟.三江平原湿地小叶章群落沿土壤水分梯度物种组成及多样性变化.生态学报,2017,37(10).
Wang J F,Han D Y,Wang J B,Fu X L,Zhu D G,Liu Y N,Cao H J,Huang Q Y,Xie L H,Zhong H X,Sui X,Ni H W.Variations in plant species composition and diversity ofCalamagrostisangustifliacommunity along soil water level gradient in the Sanjiang Plain.Acta Ecologica Sinica,2017,37(10).(in Chinese)
[17] 刘忠权,刘彤,张荣,陈辉煌.古尔班通古特沙漠南部短命植物群落物种多样性及空间分异.生态学杂志,2011,30(1):45-52.
Liu Z Q,Liu T,Zhang R,Chen H H.Species diversity and spatial differentiation of ephemeral plant community Gurbantungut desert.Chinese Journal of Ecology,2011,30(1):45-52.(in Chinese)
[18] 何艳华,闫明,张钦弟,张直峰,苗艳明,毕润成.五鹿山国家级自然保护区物种多样性海拔格局.生态学报,2013,33(8):2452-2462.
He Y H,Yan M,Zhang Q D,Zhang Z F,Miao Y M,Bi R C.Altitudinal pattern of plant species diversity in the Wulu Mounitain Nature Reserve,Shanxi,China.Acta Ecologica Sinica,2013,33(8):2452-2462.(in Chinese)
[19] 王长庭,王启基,龙瑞军,景增春,史慧兰.高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究.植物生态学报,2004,28(2):240-245.
Wang C T,Wang Q J,Long R J,Jing Z C,Shi H L.Changes in plant species diversity and productivity along an elevation gradient in an alpine meadow.Acta Phytoecologica Sinica,2004,28(2):240-245.(in Chinese)
[20] 刘哲,李奇,陈懂懂,翟文婷,赵亮,徐世晓,赵新全.青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响.生物多样性,2015,23(4):451-462.
Liu Z,Li Q,Chen D D,Zhai W T,Zhao L,Xu S X,Zhao X Q.Patterns of plant species diversity along an altitudinal gradient and its effect on above-ground biomass in alpine meadows in Qinghai-Tibet Plateau.Biodiversity Science,2015,23(4):451-462.(in Chinese)
[21] 冯建孟,王襄平,徐成东,杨元合,方精云.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局.山地学报,2006,24(1):110-116.
Feng J M,Wang X P,Xu C D,Yang Y H,Fang J Y.A ltiudinal patterns of plant species diversity and community structure on Yulong Mountains,Yunnan,China.Journal of Mountain Science,2006,24(1):110-116.(in Chinese)
[22] 孟好军,刘建泉,刘贤德,赵维俊.祁连山青海云杉群落物种组成及α多样性垂直分布.生态环境学报,2011,20(3):435-440.
Meng H J,Liu J Q,Liu X D,Zhao W J.Species composition and vertical distribution pattern of α diversity of thePiceacrassifoliacommunity in Qilin Mountains of Gansu,China.Ecology and Environmental Sciences,2011,20(3):435-440.(in Chinese)
[23] 孟兆鑫.川西亚高山森林群落物种多样性与群落结构研究.四川农业大学学报,2007,25(4):441-446.
Meng Z X.A study on community structure and species diversity of subalpine forests in western Sichuan.Journal of Sichuan Agricultural Uninversity,2007,25(4):441-446.(in Chinese)
[24] 林勇明,王道杰,周麟,陈婷婷,田兵伟,苏凤环.岷江干旱河谷植物群落分布特征及梯度效应.福建林学院学报,2009,29(4):332-335.
Lin Y M,Wang D J,Zhou L,Chen T T,Tian B W,Su F H.Distribution characters and gradient effect of species diversity of the arid valley in the upper reach of Minjiang River.Journal of Fujian College of Forestry,2009,29(4):332-335.(in Chinese)
[25] 沈泽昊,刘增力,方精云.贡嘎山海螺沟冷杉群落物种多样性与群落结构随海拔的变化.生物多样性,2004,12(2):237-244.
Shen Z H,Liu Z L,Fang J Y.Altitudinal changes in species diversity and community structure of abies fabric communities at Hailuo valley of Mt.Gongga,Sichuan.Biodiversity Science,2004,12(2):237-244.(in Chinese)
[26] 秦翔,崔晓庆,杜文涛,任贾文,陈记祖.祁连山老虎沟冰芯记录的高山区大气降水变化.地理学报,2014, 69(5):681-689.
Qin X,Cui X Q,Du W T,Ren J W,Chen J Z.Variations of the alpine precipitation during 1960-2006 recorded in Laohugou ice core in western Qilian Mountains,China.Acta Geographica Sinica,2014,69(5):681-689.(in Chinese)
[27] 傅抱璞.地形和海拔高度对降水的影响.地理学报,1992(4):302-314.
Fu B P.The effects of topography and elevation on precipitation.Acta Geographica Sinica,1992(4):302-314.(in Chinese)
[28] 杨元合,朴世龙.青藏高原草地植被覆盖变化及其与气候因子的关系.植物生态学报,2006,30(1):1-8.
Yang Y H,Piao S L.Variations in grassland vegetation cover in relation to climatic factors on the Tibetan plateau.Journal of Plant Ecology,2006,30(1):1-8.(in Chinese)
[29] 王国宏,任继周,张自和.河西山地绿洲荒漠植物群落多样性研究 .Ⅱ/ 放牧振动下草地多样性的变化特征.草业学报,2002,11(1):31-37.
Wang G H,Ren J Z,Zhang Z H.Studies on the population diversity of plant community in Hexi mountain-oasis-desert area,Gansu,China.Ⅱ.Impacts of grazing pressure on species diversity in steppe.Acta Prataculturae Sinica,2002,11(1):31-37.(in Chinese)
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
散文诗(2021年22期)2022-01-12
小哥白尼(军事科学)(2019年5期)2019-08-27
环球时报(2019-05-23)2019-05-23
活力(2019年21期)2019-04-01
乡村地理(2018年3期)2018-11-06
乡村地理(2018年4期)2018-03-23
河北林业科技(2016年5期)2016-11-08
郑州大学学报(工学版)(2014年6期)2014-03-01
