
西藏拉萨越冬黑颈鹤家庭群的警戒同步性
2018-03-22 11:28李忠秋
生态学报 2018年4期
车 烨,杨 乐,李忠秋
1 南京大学生命科学学院,南京 210023 2 西藏自治区高原生物研究所,拉萨 850000
警戒行为是指个体抬头扫视环境,以侦查外部捕食者的潜在威胁或进行物种内部的社会性监视,能够起到反捕食或维护自身地位及资源等作用[1- 2]。Pulliam于1973年提出警戒行为的经典随机独立模型[3],目前关于动物警戒行为的研究大多以此为基础。然而,近期一些关于警戒行为同步性的理论和实证研究向经典模型发出了挑战。研究者发现个体能够利用周围同伴所表现出的行为、声音等预示着潜在捕食危险的警戒信息来调整自身的警戒行为,并且常常表现为模仿同伴的警戒状态[4- 9]。根据警戒行为同步性程度的不同,集体警戒可能存在基于模仿的同步警戒和避免重复的协调警戒两种模式。研究警戒行为的同步性,探究非随机警戒模式的产生机制与适用情况,不仅是对经典学说的补充,也为全面了解野生动物的警戒行为提供了新的研究方向,尤其为濒危物种的保护给予了重要的参考意义。
黑颈鹤是一种濒危鸟类,在以往研究中已证实群体大小、捕食脆弱性和空间位置等因素对其警戒行为的影响[10- 12],但尚未涉及警戒同步性。因此,研究了西藏拉萨雅鲁藏布江中游河谷越冬黑颈鹤家庭群的警戒行为,旨在验证由于年龄和是否携幼的差异将导致个体警戒水平的差异,探究3种家庭类型集体警戒是否存在差异性,并且预期越冬黑颈鹤家庭群与丹顶鹤家庭群、群体大小小于5的白头鹤[13- 14]集群类似,表现出同步警戒模式。
1 材料与方法
1.1 研究地点
研究地点位于中国西藏自治区拉萨市林周县的雅鲁藏布江中游河谷黑颈鹤国家级自然保护区(28°40′—30°17′N, 87°34′—91°54′E),海拔高度3500—4500m,面积约6143km2,所处区域为高原温带半干旱季风气候,日温差大,夏季无高温,干湿季明显,雨季降水集中,冬季偶有降雪,1月平均气温-4.7°C。区域内自然植被以砂生槐(Sophoramoorcroftiana)、伴生小角柱花(Ceratostigmaminus)、三刺草(Aristidatriseta)、固沙草(Orinusthoroldii)、白草(Pennisetumcentrasiaticum)和紫花针茅(Stipapurpurea)等高山草甸植被为主。除自然植被外,还有种植青稞(Hordeumvulgare)和冬小麦(Triticumspp.)的农田,黑颈鹤主要在这些农田中觅食,以遗留谷物为主要食物[12]。农田是越冬黑颈鹤行为观察和数据采集的主要区域。
1.2 研究对象
黑颈鹤是国家一级保护动物,全球濒危物种,是唯一一种繁殖期和越冬期均栖息于高原地区的鸟类。主要繁殖地包括青藏高原及其邻近地区,越冬区域主要分布在西藏中南部、云贵高原、印度和不丹等区域[12]。在雅鲁藏布江中游河谷黑颈鹤国家级自然保护区越冬的黑颈鹤12月初从繁殖地迁来,四月初迁离[15- 16]。黑颈鹤越冬栖息地的主要植被类型包括河谷灌丛、草原、湿地、农田等[15,17]。越冬期,黑颈鹤常见的集群模式有3种:家庭群、同种集群和混种集群。家庭群由两只成年鹤以及0—2只幼年黑颈鹤组成,同种集群由未繁殖个体或几个家庭共同组成,混种集群是指黑颈鹤与灰鹤、斑头雁、赤麻鸭等物种混群[17]。越冬黑颈鹤成鹤与幼鹤体型大小近似,但羽毛特征差异明显,因而非常容易分辨。
1.3 数据采集
2015年12月至2016年1月,每天9:00至19:00沿不同的路线,采取步行或驾车的方式寻找黑颈鹤家庭群,并使用GPS定位,以避免重复取样。在发现目标家庭群后,使用摄像机(Sony cx270)进行拍摄,拍摄地点通常距离鹤群200—400m,以在保证视频清晰度的同时尽量减少可能对鹤群造成的干扰[18]。同时记录采样日期、时间、经纬度、生境类型、家庭组成类型(两成、两成一幼、两成两幼)等。连续拍摄30min,若鹤群出现群体大小改变、迁飞或周围环境出现明显的干扰,如牲畜冲撞使黑颈鹤受到惊吓、噪音过大的拖拉机驶过等情况时,提前停止拍摄。在强风、降雪等恶劣天气下不进行数据采集。
1.4 数据分析
使用拍摄时间超过20min的视频作为分析样本,回放视频并利用焦点动物法和扫描取样法统计个体和集体警戒水平,每次扫描间隔30s。对于家庭内每一只个体k,其个体警戒水平为扫描到该焦点个体处于警戒状态的次数占扫描总次数的比例,记为pk。利用焦点动物法时,每次仅始终观察视频中的一只个体,观察结束后再更换统计家庭内其他个体,直至完成对每只个体警戒行为的扫描取样。对于一个家庭,所观测到的集体警戒水平为扫描到该家庭内至少有一只个体处于警戒状态的次数占扫描总次数的比例,记为Vobs。警戒行为定义为当黑颈鹤停止觅食,脖子向上伸长且向周围张望[11- 12]。
为保证数据的正态性,对所得数据进行了平方根变换。使用独立样本t检验比较成鹤与幼鹤、携幼成鹤与非携幼成鹤的个体警戒水平的差异性。在检验个体警戒行为的发生是否受到家庭内其他个体的影响,即越冬黑颈鹤家庭群是否采用同步警戒或协调警戒模式时,在每个家庭内,比较了实际观察到的集体警戒水平数值Vobs与独立假设下计算出的预期集体警戒水平数值Vexp。预期值计算公式为Vexp=1-∏(1-pk),其中pk代表个体k(1≤k≤n)花费在警戒行为上的时间比例[13,19- 20],n代表群体大小。使用两种方式确定n值,一种衡量方式为仅考虑家庭内成年黑颈鹤的数量;另一种为家庭内全部个体的数量。相对应地,在统计集体警戒的实际观测值时,也分别以家庭内至少有一只成鹤或至少有一只个体处于警戒状态作为是否发生了集体警戒的标准,以保证在两种衡量方式下统计观测值与计算预期值时个体数量是一致的。通过配对t检验比较Vobs与Vexp,如果两者没有显著差异,则说明群体内个体是独立对周围环境进行扫视的,彼此之间不存在模仿和相互影响;如果观测值大于预期值,则说明个体倾向于形成非重叠的警戒片段,因而采取协调警戒模式;如果观测值小于预期值,则说明个体倾向于模仿同伴的警戒行为,因而采取同步警戒模式。此外,我们还随机选取家庭群中的一只成鹤与一只幼鹤,在计算其Vobs与Vexp后,同样使用配对t检验比较家庭内部成鹤与幼鹤之间的同步性。
所有数据分析使用SPSS 18.0软件完成,显著水平为P=0.05,文中数据以未正态校正前的实际平均值±标准误差(Mean±SE)的形式列出。
2 结果
2.1 采样统计
研究中共拍摄了63个越冬黑颈鹤家庭,合计时间1604.83min(时间范围:4.4—35.3min),平均时间(25.9±1.1)min。选择其中39段(61.9%)画面清晰、全程无遮挡且持续拍摄时间在20min以上的视频作为扫描样本,包括两成型家庭13个(33%)、两成一幼型家庭19个(49%)、两成两幼型家庭7个(18%),平均群体大小2.85。观察对象中成年黑颈鹤78只,幼年黑颈鹤33只,个体统计时间共计3121.5min,平均时间(28.2±0.5)min。
2.2 个体警戒
独立样本t检验的结果显示,成年黑颈鹤警戒时间比例(0.199±0.014)极显著(t109=6.495,P<0.001)高于幼鹤(0.074±0.011)。有幼鹤存在的家庭中,成鹤花费在警戒行为的时间比例为0.210±0.017,略高于家庭中没有幼鹤的成鹤(0.178±0.024),但未达到显著水平(t76=1.093,P=0.278)。
2.3 集体警戒同步性
对3种类型的家庭群集体警戒水平进行单因素方差分析的结果表明,无论是仅考虑家庭内成鹤(F2,36=1.712,P=0.195,图1A),还是包括幼鹤在内(F2,36=2.876,P=0.069,图1B),3种类型的越冬黑颈鹤家庭的集体警戒水平均不存在显著差异。

图1 三种类型的越冬黑颈鹤家庭群集体警戒水平比较Fig.1 Comparisons of collective vigilance in three family types of wintering black-necked crances(A)仅考虑家庭内成鹤的情况下;(B)考虑家庭内全部个体的情况下
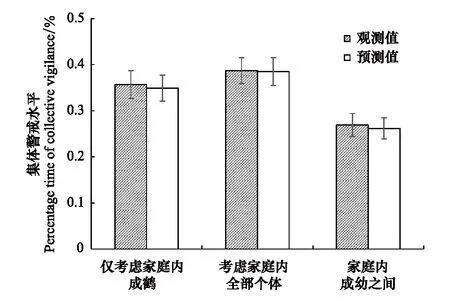
图2 集体警戒实际观测值与独立假设预测值的比较 Fig.2 Comparisons of observed and expected collective vigilance
利用实际观测到的集体警戒数值与独立假设下的预测值进行比较,不论是仅考虑家庭内的成鹤(Vobs=0.357±0.030,Vexp=0.349±0.028,图2),还是包括幼鹤在内的家庭中全部个体(Vobs=0.387±0.028,Vexp=0.385±0.030,图2),或者家庭内部的成鹤与幼鹤之间(Vobs=0.269±0.025,Vexp=0.262±0.023,图2),配对t检验的结果均无显著差异(仅考虑成鹤:t38=0.770,P=0.446;家庭内全部个体:t38=-0.760,P=0.452;家庭内成幼之间:t25=-0.734,P=0.470)。
3 讨论
3.1 个体警戒水平
与预期一致,在本研究中发现,家庭群中的幼年黑颈鹤花费在警戒行为的时间显著低于成年黑颈鹤。这与王凯等[21]对云南越冬黑颈鹤成幼行为模式的研究结果一致,也与丹顶鹤、白头鹤的研究结果相同[13,22- 23]。这与幼年个体缺乏足够的警戒和逃脱捕食者的能力[24],且食物获取效率较低,需花费更多时间寻找食物和进食,以保证生长需求[22]有关。
Xu等[10]研究了黑颈鹤捕食脆弱性与警戒强度的关系,结果表明由于幼鸟更易被捕食者攻击,因而携幼群体是高捕食脆弱性的[25],其群体内的成鹤比非携幼成鹤(低捕食脆弱性)明显更加警惕。但是研究没有发现家庭群内幼鹤存在与否对成鹤的警戒水平有影响。这一结果的不同,可以从幼鹤年龄和警戒需求来源这两方面来解释。首先,Xu等关于黑颈鹤捕食脆弱性与警戒强度的研究时间为九月份,此时黑颈鹤群体处于繁殖期,幼鹤年龄约3个月,体型较小,因此需要亲代的照顾,而本研究中的黑颈鹤越冬期,幼鹤年龄约6—7个月,体型大小已接近成鹤[12]。其次,Xu等的研究区域是人类干扰极低的保护区核心区,几乎不存在成年黑颈鹤的捕食者,但幼鹤面临较大的捕食风险,因此携幼成鹤与非携幼成鹤因幼鹤的存在与否形成了捕食脆弱性的差异,而我们的研究区域是有人类农业活动的农田,人类干扰是黑颈鹤最主要的警戒需求来源,而非捕食风险。并且这些人类干扰对不同类型的家庭来说是普遍存在且强度相似的,因此成鹤的警戒水平没有由于幼鹤的存在而表现出差异。
3.2 集体警戒同步性
众多研究已经表明,群体内个体警戒行为的发生并非独立于其他个体,而是通过模仿或避免重复的形式调节自己的警戒行为[4- 9],因而在群体水平形成了同步警戒或协调警戒这两种集体警戒模式。同步警戒是指群体内个体具有模仿周围同伴警戒状态的倾向,在此模式下,集体警戒水平将低于独立假设下的预期水平,表现为存在更多的群体成员中没有任何一个个体警戒的空白期,预期会出现集体警戒波[8- 9,19,26- 27]。同步警戒虽然无益于集体警戒水平的增强,却有利于个体被捕食风险的降低,并且为群体成员共同评估风险提供可能[5- 6,28]。协调警戒是指群体内个体具有避免与周围同伴发生重复的警戒行为的倾向,在此模式下,集体警戒水平将高于独立假设下的预期水平,能够有效降低群体警戒空白期的比例[27,29- 30]。协调警戒是高效但代价巨大的,因为使集体警戒水平得以加强的同时,个体需要时刻监视周围同伴并采取相应的行为[31- 33]。
然而,本研究对越冬黑颈鹤家庭群集体警戒模式的研究中并未如预期一样发现同步倾向,原因可能是多方面的。
当食物可用性成为生存的关键因素时,此时能量的获取优先于安全保证,对食物的竞争或觅食的迫切需求将导致警戒水平的降低[34],继而也导致警戒同步性的降低。这一方面是因为警戒同步性的表现依赖于个体的警戒水平,冬季等食物紧缺情况下个体本来的警戒水平已经降低,警戒同步性也随之降低;另一方面,对食物获取的优先性也削弱了群体以降低捕食风险为功能的警戒同步性[5- 6,20]。由于收割期后的农业活动,如:耕作和牲畜放牧等,黑颈鹤作为食物的遗留谷粒是有限的[35],因此,缺乏食物的越冬黑颈鹤家庭群很有可能由于上述两个原因而没有表现出警戒行为的同步性。
警戒行为的首要作用是发现潜在威胁,避免成为捕食者的猎物,而不论是同步警戒亦或是协调警戒都是一种与特定环境以及被捕食风险相关的适应性的警戒模式。群体中较早察觉到捕食者的个体更容易逃脱,从而有效降低了自身的被捕食风险,而晚察觉或晚逃离的个体则容易被捕捉[28,36- 37]。基于监视和模仿产生的同步警戒,能够避免个体成为捕食者发起攻击时最后一个风险察觉者或群体逃离捕食者时最后一个响应的成员[14,30,38]。因此,当捕食者存在对落后个体的捕食倾向时(落后目标策略捕食者),预期警戒行为同步性将得到强化[8]。而本研究中的黑颈鹤是一种体型较大的鸟类,越冬期幼鹤个体大小已接近成鹤[12],其警戒来源主要是所觅食区域内人类活动的干扰,并不存在落后目标策略捕食者,因而也不存在其对警戒行为同步性的强化作用。此外,在青藏高原繁殖、越冬的黑颈鹤与在当地聚居的藏族人民世代生活在一起,已深深融入到当地的历史文化、古代传说、地域风俗习惯,甚至宗教信仰当中,黑颈鹤在藏族地区普遍被视为吉祥的象征,被称为神鸟而备受保护[39]。所以,虽然在农田觅食的黑颈鹤不可避免地会受到人类干扰,但横向比较其他研究物种在其他保护区内的游客干扰、狩猎风险[14,20],当地的人类干扰程度也许并不足以诱发黑颈鹤高强度的警戒行为同步性。
[1] Childress M J, Lung M A. Predation risk, gender and the group size effect: does elk vigilance depend upon the behaviour of conspecifics? Animal Behaviour, 2003, 66(2): 389- 398.
[2] Gaynor K M, Cords M. Antipredator and social monitoring functions of vigilance behaviour in blue monkeys. Animal Behaviour, 2012, 84(3): 531- 537.
[3] Pulliam H R. On the advantages of flocking. Journal of Theoretical Biology, 1973, 38(2): 419- 422.
[4] Fernández-Juricic E, Siller S, Kacelnik A. Flock density, social foraging, and scanning: an experiment with starlings. Behavioral Ecology, 2004, 15(3): 371- 379.
[5] Sirot E. Social information, antipredatory vigilance and flight in bird flocks. Animal Behaviour, 2006, 72(2): 373- 382.
[6] Roth II T C, Cox J G, Lima S L. The use and transfer of information about predation risk in flocks of wintering finches. Ethology, 2008, 114(12): 1218- 1226.
[7] Beauchamp G. Sleeping gulls monitor the vigilance behaviour of their neighbours. Biology Letters, 2009, 5(1): 9- 11.
[8] Pays O, Goulard M, Blomberg S P, Goldizen A W, Sirot E, Jarman P J. The effect of social facilitation on vigilance in the eastern gray kangaroo,Macropusgiganteus. Behavioral Ecology, 2009, 20(3): 469- 477.
[9] Beauchamp G, Alexander P, Jovani R. Consistent waves of collective vigilance in groups using public information about predation risk. Behavioral Ecology, 2011, 23(2): 368- 374.
[10] Xu F, Ma M, Yang W K, Blank D, Ding P, Zhang T. Vigilance in Black-necked Cranes: effects of predation vulnerability and flock size. The Wilson Journal of Ornithology, 2013, 125(1): 208- 212.
[11] Kuang F L, Li F S, Liu N, Li F Q. Effect of flock size and position in flock on vigilance of black-necked cranes (Grusnigricollis) in winter. Waterbirds, 2014, 37(1): 94- 98.
[12] Yang L, Zhuom C J, Li Z Q. Group size effects on vigilance of wintering black-necked cranes (Grusnigricollis) in Tibet, China. Waterbirds, 2016, 39(1): 108- 113.
[13] Ge C, Beauchamp G, Li Z Q. Coordination and synchronisation of anti-predation vigilance in two crane species. PLoS One, 2011, 6(10): e26447.
[14] Li C L, Beauchamp G, Wang Z, Cui P. Collective vigilance in the wintering hooded crane: the role of flock size and anthropogenic disturbances in a human‐dominated landscape. Ethology, 2016, 122(12): 999- 1008.
[15] Bishop M A, Zhuoma C, Song Y L, Harkness J, Gu B Y. Winter habitat use by black-necked cranesGrusnigricollisin Tibet. Wildfowl, 1998, 49: 228- 241.
[16] 仓决卓玛, 杨乐, 李建川. 西藏黑颈鹤越冬期昼间行为的时间分配. 野生动物, 2008, 29(1): 15- 20.
[17] 仓决卓玛, 杨乐, 李建川, 央金卓嘎. 西藏黑颈鹤的保护与研究现状. 四川动物, 2008, 27(3): 449- 453.
[18] 李忠秋. 丹顶鹤行为观察的适宜距离: 简评观察者效应(英文). Chinese Birds, 2011, 2(3): 147- 151.
[19] Pays O, Renaud P C, Loisel P, Petit M, Gerard J F, Jarman P J. Prey synchronize their vigilant behaviour with other group members. Proceedings of the Royal Society B: Biological Sciences, 2007, 274(1615): 1287- 1291.
[20] Podgórski T, De Jong S, Bubnicki J W, Kuijper D P J, Churski M, Jędrzejewska B. Drivers of synchronized vigilance in wild boar groups. Behavioral Ecology, 2016, 27(4): 1097- 1103.
[21] 王凯, 杨晓君, 赵健林, 余红忠, 闵龙. 云南纳帕海越冬黑颈鹤日间行为模式与年龄和集群的关系. 动物学研究, 2009, 30(1): 74- 82.
[22] Li Z Q, Wang Z, Ge C. Time budgets of wintering red-crowned cranes: effects of habitat, age and family size. Wetlands, 2013, 33(2): 227- 232.
[23] Li C L, Zhou L Z, Xu L, Zhao N N, Beauchamp G. Vigilance and activity time-budget adjustments of wintering hooded cranes,Grusmonacha, in human-dominated foraging habitats. PLoS One, 2015, 10(3): e0118928.
[24] Weathers W W, Sullivan K A. Juvenile foraging proficiency, parental effort, and avian reproductive success. Ecological Monographs, 1989, 59(3): 223- 246.
[25] Wang Z, Li Z Q, Beauchamp G, Jiang Z G. Flock size and human disturbance affect vigilance of endangered red-crowned cranes (Grusjaponensis). Biological Conservation, 2011, 144(1): 101- 105.
[26] Pays O, Sirot E, Fritz H. Collective vigilance in the greater kudu: towards a better understanding of synchronization patterns. Ethology, 2012, 118(1): 1- 9.
[27] Pays O, Jarman P J, Loisel P, Gerard J F. Coordination, independence or synchronization of individual vigilance in the eastern grey kangaroo? Animal Behaviour, 2007, 73(4): 595- 604.
[28] Quinn J L, Cresswell W. Escape response delays in wintering redshank,Tringatotanus, flocks: perceptual limits and economic decisions. Animal Behaviour, 2005, 69(6): 1285- 1292.
[29] Rodríguez-Gironés M A, Vásquez R A. Evolutionary stability of vigilance coordination among social foragers. Proceedings of the Royal Society B: Biological Sciences, 2002, 269(1502): 1803- 1810.
[30] Sirot E, Touzalin F. Coordination and synchronization of vigilance in groups of prey: the role of collective detection and predators′ preference for stragglers. The American Naturalist, 2009, 173(1): 47- 59.
[31] Ward P I. Why birds in flocks do not coordinate their vigilance periods. Journal of Theoretical Biology, 1985, 114(3): 383- 385.
[32] Lima S L. Back to the basics of anti-predatory vigilance: the group-Size effect. Animal Behaviour, 1995, 49(1): 11- 20.
[33] Ruxton G D, Roberts G. Are vigilance sequences a consequence of intrinsic chaos or external changes? Animal Behaviour, 1999, 57(2): 493- 495.
[34] Beauchamp G, Ruxton G D. Changes in vigilance with group size under scramble competition. The American Naturalist, 2003, 161(4): 672- 675.
[35] Bishop M A, Li F S. 李凤山. 农业耕作活动对西藏越冬黑颈鹤食性及食物可获得性的影响. 生物多样性, 2002, 10(4):393- 398.
[36] Hilton G M, Cresswell W, Ruxton G D. Intraflock variation in the speed of escape-flight response on attack by an avian predator. Behavioral Ecology, 1999, 10(4): 391- 395.
[37] Bednekoff P A, Lima S L. Re-examining safety in numbers: interactions between risk dilution and collective detection depend upon predator targeting behaviour. Proceedings of the Royal Society B: Biological Sciences, 1998, 265(1409): 2021- 2026.
[38] Sirot E. Negotiation may lead selfish individuals to cooperate: the example of the collective vigilance game. Proceedings of the Royal Society B: Biological Sciences, 2012, 279(1739): 2862- 2867.
[39] 李筑眉, 李凤山. 黑颈鹤研究. 上海: 上海科技教育出版社, 2005: 13.
猜你喜欢
廊坊师范学院学报(自然科学版)(2021年2期)2021-09-10
学苑创造·C版(2019年8期)2019-08-09
学苑创造·C版(2019年5期)2019-06-14
意林·少年版(2019年2期)2019-01-16
小哥白尼(军事科学)(2018年9期)2018-12-08
小哥白尼(军事科学)(2018年8期)2018-09-12
小哥白尼(军事科学)(2018年6期)2018-09-10
小哥白尼(军事科学)(2018年1期)2018-05-25
医学研究杂志(2015年11期)2015-06-10
医学研究杂志(2015年11期)2015-06-10
