
西非沿岸乌贼角质颚形态及生长特征*
2018-03-29 08:33瞿俊跃李建华陈新军
渔业科学进展 2018年2期
瞿俊跃 李建华,2,3① 陈 芃 陈新军,2,3 靳 楠
(1. 上海海洋大学海洋科学学院 上海 201306;2. 国家远洋渔业工程技术研究中心 上海 201306;3. 大洋渔业资源可持续开发教育部重点实验室 上海 201306)
乌贼(Sepia officinalisLinnaeus, 1758)是一种季节洄游性的浅海性底栖种,主要栖息在东大西洋波罗的海到南非沿岸200 m水深的砂质和烂泥质底层区域,而100 m以上的深度区域群体较为密集,大个体则分布在更深的水层(Bakhayokhoet al, 1982)。乌贼是大西洋最重要的商业性捕捞种类之一,同时也是我国西非过洋性渔业的重要捕捞对象。联合国粮农组织的捕捞统计数据显示,2004~2012年乌贼的产量不断增加,在2012年达到29589 t,近几年其产量虽有回落,但也保持在20000 t左右。
角质颚作为头足类的摄食器官,蕴含着许多生物学信息(刘必林等, 2010; 陈芃等, 2015),被认为是用来鉴定头足类尤其是蛸类年龄最可靠的硬组织之一;且角质颚比耳石大,具有容易提取、研磨方便等优点,在年龄与生长的研究中得到广泛的应用(方舟等,2014b)。角质颚有耐腐蚀的特点,因此,在摄食生态学中也有应用,例如,残留在大型海洋生物体内大量的角质颚可以用来分析头足类在食物网中的地位(Olsonet al, 2007)。此外,对角质颚的形态特征、色素沉着、稳定同位素和微量元素变化的分析有助于对头足类群体划分和资源动态等的理解(方舟等,2014b)。然而,国内对于角质颚的研究主要集中在柔鱼类,如柔鱼(Ommastrephes bartramii)(方舟等, 2014a;陈芃等, 2015)、茎柔鱼(Dosidicus gigas)(胡贯宇等,2015、2016)、太平洋褶柔鱼(Todarodes pacificu)(杨林林等, 2012)等。对乌贼目种类的角质颚研究还十分匮乏。为此,本研究根据2015年在西非毛里塔尼亚附近海域所获得的渔获样本,对乌贼角质颚样本进行分析,并通过角质颚的各项形态参数及其比值对乌贼的角质颚进行形态描述,探究不同胴长和不同性腺成熟阶段角质颚生长情况的差异,并利用角质颚的形态参数对个体大小进行估算,以期为东大西洋乌贼的形态学和生态学研究提供基础资料。
1 材料与方法
1.1 采集时间和地点
乌贼样本来源于2015年中国水产总公司渔船在西非毛里塔尼亚专属经济区海域底拖网作业获得的渔获,随机采集983尾。所获得的样本经冷冻保藏运回实验室。
1.2 角质颚采集
从乌贼头部口器中提取角质颚(图1),得到完整角质颚样本898对。将取出的角质颚进行编号并存放于装有75%乙醇溶液的50 ml离心管中。

图1 乌贼角质颚形态Fig.1 Common cuttlefish beaks
1.3 研究方法
1.3.1 基础生物学测量 将实验室中冷冻的样品取出并解冻后,对乌贼进行生物学测定,包括胴长(Mantel length, ML)、性别和性腺成熟度。测量胴长用皮尺进行,测定精确至1 mm。根据Lipinski等(1995)的分类方法,将性成熟度划分Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ共5期,本研究中,Ⅰ、Ⅱ期个体为性未成熟的个体,Ⅲ、Ⅳ期为性成熟个体,Ⅴ期为繁殖后(雄性为交配后,雌性为产卵后)的个体。
1.3.2 角质颚形态测量 将角质颚从离心管中取出,将其外部清洗干净后,使用数显游标卡尺对角质颚的上头盖长等12项形态参数进行测量(图2),测量结果精确至0.01 mm。
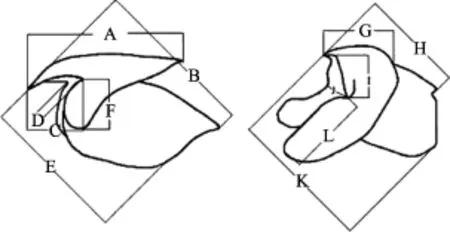
图2 角质颚外部形态测量示意(Fang et al, 2014)Fig.2 Scheme of morphometric measurement for beak
1.3.3 数据处理 角质颚形态描述按照 Clarke(1962、1986)的研究方法,分别计算上下角质颚喙长与头盖长(RL/HL)、喙长与脊突长(RL/CL)、头盖长与脊突长(HL/CL)、翼长与脊突长(WL/CL)、翼长与头盖长(WL/HL)和翼长与喙长(WL/RL)的比值,对角质颚的形态特征进行描述。
按照性别分别计算各角质颚形态参数的平均值和标准差,并利用t检验(李春喜等, 2008)检验不同性别各角质颚形态参数的差异。为校正样品规格差异对形态参数值的影响,t检验时,将其形态参数均除以胴长,即对 UHL/ML、UCL/ML、URL/ML、URW/ML、ULWL/ML、UWL/ML、LHL/ML、LCL/ML、LRL/ML、LRW/ML、LLWL/ML和LWL/ML这12个指标进行分析。
按胴长将乌贼分为 100~150、150~200、200~250和250~300 mm 4个胴长组,利用方差分析(ANOVA)(Gueorguievaet al, 2004)及多重比较(LSD法) (王斌会等, 2010)对不同胴长组及不同性成熟度间角质颚各项形态参数值的差异进行检验,以分析不同因子对角质颚生长的影响。
首先利用主成分分析(Tanget al, 2007),对乌贼角质颚的12个比例形态指标进行分析,得到累计方差贡献率达到 70%的主成分(陈芃等, 2015),并计算各个样本的各主成分得分。其次,利用线性回归法拟合胴长与各主成分之间的关系式作为利用角质颚估算乌贼个体大小的方程式。上述分析均采用 SPSS 19.0软件进行。
2 结果
2.1 角质颚外部形态特征
上颚特征:角质颚上颚的 HL较长,长度为CL的77.11%,上喙较平缓,下喙连接翼部。喙较弯曲,RL分别达到HL和CL的23.76%和18.29%。侧壁较宽大,其后端边缘和脊突弯曲,均呈一定的弧度。翼部连接到侧壁前端的底部,WL大于 RL,为 RL的194.28%,但相对于HL和CL仍旧较小,分别为HL和 CL的 45.39%和 34.95%(表1)。
下颚特征:角质颚下颚 HL的长度为 CL的46.04%,喙较长,RL分别达到HL和CL的49.97%和 26.63%。喙与翼部的夹角较上颚大,侧壁较上颚小,其后端边缘和脊突均比较平直。翼部较长,约为侧壁的2倍,WL显著大于HL和RL,分别为HL的228.01%,RL的 476.34%,WL和 CL相当,为 CL的 103.21%(表1)。

表1 乌贼角质上、下颚各长度指标的比值Tab.1 Relative ratios of the upper and lower beaks of common cuttlefish
2.2 不同性别乌贼个体角质颚形态特征比较
不同性别乌贼个体角质颚形态存在差异(表2),除上喙宽、下喙长、下喙宽外,雄性个体的角质颚形态参数的平均值均大于雌性(表1)。但t检验表明,将各项形态参数均除以胴长消除了样品规格差异的影响后,发现雄性个体在角质颚的各项形态参数上均极显著小于雌性(P<0.01)。
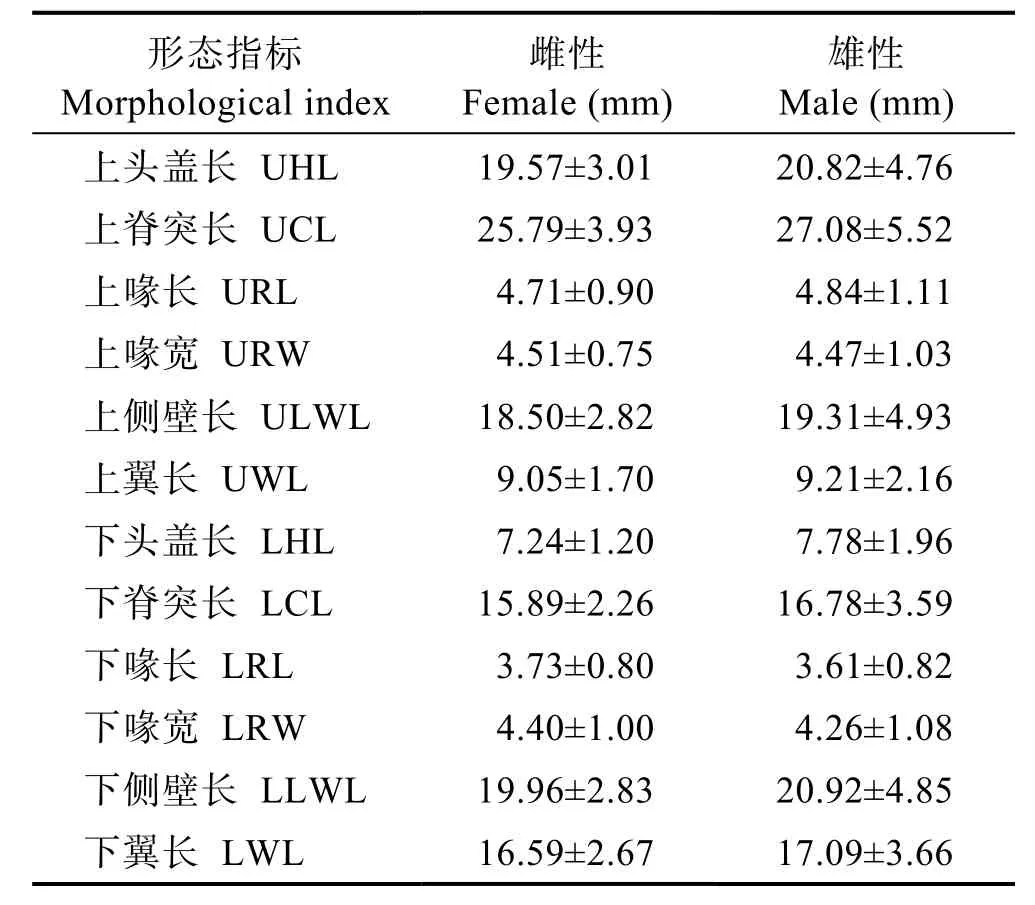
表2 不同性别乌贼个体角质颚形态参数均值Tab.2 Average values of morphological parameters of common cuttlefish beak in both genders
2.3 胴长和性腺成熟度与乌贼角质颚形态参数的关系
样本在 100~150、150~200、200~250 和 250~300 mm胴长组的比例分别为0.3%、5.76%、46.01%、26.1%和16.89%。ANOVA分析表明,乌贼群体不同胴长组和角质颚各项指标均存在显著差异(P<0.01)。LSD分析显示,随着胴长的增加,乌贼的各个角质颚的形态参数也极显著增加(图3),所以,可以根据角质颚的长度对乌贼的胴长及生长情况进行判定。
乌贼群体雌性样本的性腺成熟度在Ⅱ~Ⅴ期之间,所占比重分别为1.93%、12.08%、85.51%和0.48%。乌贼群体雄性的性成熟度在Ⅱ~Ⅳ之间,所占比重分别为13.39%、57.82%和28.79%。ANOVA分析表明,乌贼个体角质颚各项形态参数在不同性腺成熟度均存在显著差异(P<0.01)。LSD分析显示,角质颚形态参数在性成熟度Ⅱ~Ⅳ期显著增加(P<0.05),而性成熟度Ⅴ期时角质颚形态参数显著小于性成熟度Ⅱ~Ⅳ期(P<0.05)。
2.4 角质颚形态参数主成分分析及其与大小的关系
对乌贼12个角质颚形态参数指标进行主成分分析,结果显示(表3):前2个主成分的累计方差贡献率已达到76.5%,其中第一主成分负载绝对值最高的前2个指标为UHL和LCL;第二主成分负载绝对值最高的前2个指标为LRL和LRW。

图3 胴长与乌贼角质颚形态参数的关系Fig.3 The relationship between mantle length and beak shape parameters of common cuttlefish
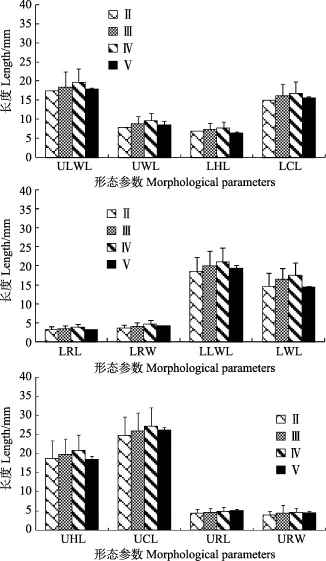
图4 性腺成熟度与乌贼角质颚形态参数的关系Fig.4 The relationship between morphological parameters and maturity stage in common cuttlefish beak
根据主成分分析的结果,选取前2个主成分,并利用各个角质颚的形态参数计算各主成分的得分,以此拟合乌贼胴长和各主成分得分的关系式,结果如下(表3):
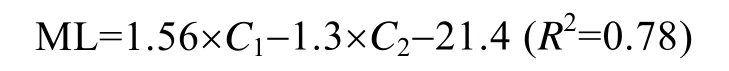
式中,ML为胴长,C1为第一主成分得分,C2为第二主成分得分。方差分析显示,胴长与这2个主成分得分间呈极显著的线性关系(F=1468.13,df1=2,df2=896,P<0.01)。验证结果表明(表4,图5),该方程式可作为利用角质颚估算乌贼个体大小的关系式。
3 讨论
3.1 角质颚的形态
本研究显示,乌贼角质颚下颚的长度比值波动较大,比较不稳定。除了 HL/CL外,其余的角质颚长度比值标准差均大于上颚(表1)。这可能是与上颚肌相较于侧肌和下颚肌较发达有关(Kear, 1994)。同时也表明,上颚相对于下颚可更好地作为种类划分的指标。上、下颚中,WW/RL比值的标准差远大于其他各项长度比值,可能是由于翼部与喙部相连且有一定的弯曲,导致测量喙长及翼长较困难,造成结果出现了一定的误差,造成比值波动较大。Arimoto等(1998)发现头足类角质颚的结构相对稳定,存在明显的特征,在种群鉴定上比其他的软体部位更为有效。而不同种类角质颚在长度比值上可能存在特异性,可以作为种类判别的指标。根据对其他乌贼类的报道,发现角质颚长度比值的稳定特性甚至可以用来判别不同的种类。例如,日本无针乌贼(Sepiella japonica)下颚WL/HL为1.67±0.28 (刘本伟等, 2016),该值小于毛里塔尼亚附近海域乌贼的2.28±0.53(表1),虽然置信区间略有重叠,但可以作为判别这2种种类的佐证。
同样,不同种类的个体以及同一种类不同性别的个体角质颚形态参数存在特异性。主成分分析中,UHL和LCL在第一主成分中的绝对值最大,而第二主成分中,绝对值最大的是 LRL和 LRW(表3)。这与对前人对柔鱼(Ommastrephes bartramii)(陈芃等,2015; 方舟等, 2014a)、阿根廷滑柔鱼(Illex argentinus)的研究结果(方舟等, 2012)不尽相同,例如方舟等(2014a)通过主成分分析结果发现,柔鱼角质颚脊突、侧壁和喙长的快速生长为其摄食提供便利,这与乌贼的角质颚较为相似,但与柔鱼相比,上头盖长也可以作为代表乌贼角质颚的指标,可能是由于种类不同、物种摄食情况不同和栖息环境不同(乌贼为底栖种类,而柔鱼为中上层种类)导致的,这有待于今一步分析。不同性别上角质颚形态参数同样存在特异性。除以胴长后,雌性个体在角质颚的各项形态参数上均极显著大于雄性(表2)。这可能由于同一种群的雌性个体生长快于雄性,需求的食物量较大,造成角质颚形态参数的差异。
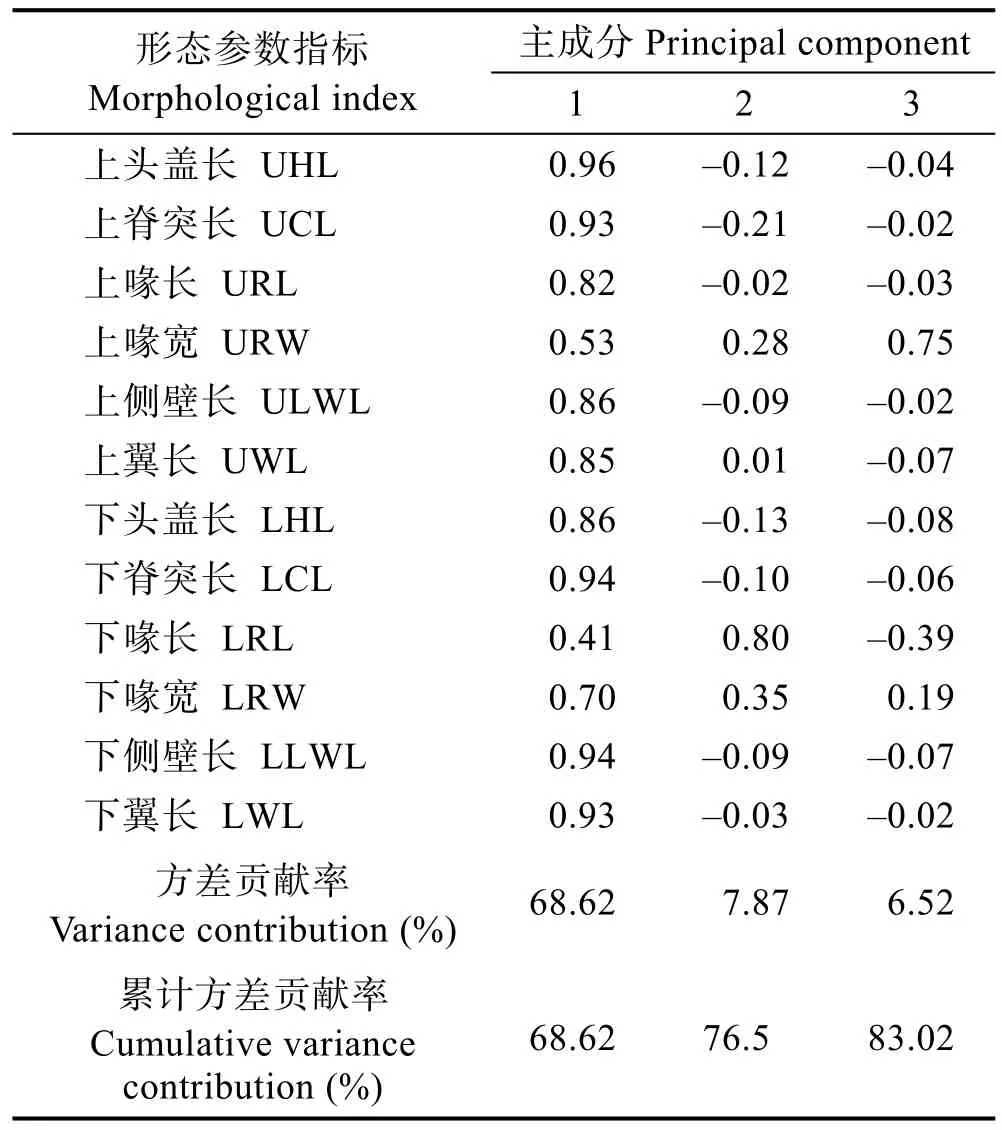
表3 乌贼角质颚形态参数指标的主成分分析结果Tab.3 Results of principal analysis of morphologicalindex of common cuttlefish beak

表4 回归分析及其检验结果Tab.4 Results of regression analysis
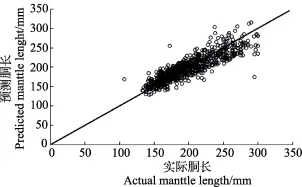
图5 利用角质颚预测乌贼胴长与实际胴长比较Fig.5 The comparison between actual and predicted mantle length based on the beak
3.2 角质颚的生长
研究显示,乌贼上、下颚各形态参数随着胴长组的增加而极显著增加,同时,利用主成分分析可以从12项形态参数中提取出形态差异(即前面角质颚个体大小与角质颚长度关系显著)的主要信息,因此,可以利用角质颚形态参数估算乌贼个体的大小(图5)。在性腺成熟度与乌贼角质颚关系的比较中发现,乌贼角质颚的形态参数随着性腺成熟度Ⅱ~Ⅳ期(即性未成熟到性成熟)增加而显著增加(图3),但是,性腺成熟度为Ⅴ期的个体的角质颚形态参数却显著小于前面几期,一种解释为,可能在毛里塔尼亚附近存在着乌贼的不同群体,Ⅴ期的个体所代表的群体,普遍个体较小且产卵早。这个假设有待于今后进一步验证。
3.3 角质颚形态特征的应用
目前,渔业资源学的研究重点主要集中在年龄生长、资源评估和食性分析上(Hernández-Lópezet al,2001)。角质颚因其自身的相对稳定的结构从而在这些方面具有较好的应用前景(Smithet al, 2000)。但对乌贼的生活史还无充分了解,作为硬组织的角质颚保存着大量信息,还可以通过其他方法,如角质颚研磨获得日龄情况(Hernández-Lópezet al, 2001; Rayaet al, 1998)、氨基酸(Miserezet al, 2007)及色素成分组成分析(García,2003)等来发现并判断其中信息所包含的意义。
本研究阐述了乌贼角质颚的形态参数特征。通过对角质颚与胴长及性腺成熟度关系的分析,一定程度上可推断出个体的生长发育情况。并利用角质颚的形态参数建立了乌贼个体大小估算方程式。但是,本研究所得到的捕捞的样本在Ⅰ期和Ⅴ期的个体数量较少,可能会对研究结果带来误差,还需要后续的研究和补充。
Arimoto Y, Kawamura A. Characteristics of the fish prey of neon flying squid,Ommastrephes bartramii, in the central North Pacific. Heisei, 1998, 8: 70–80
Bakhaykho M, Drammeh O. Elements de biologie et d'identite des populations de seiches (Sepiaofficinalis hierredda)des cotes sene-gambiennes. Special Working Group on Cephalopod Stocks in the Northern Region of CECAF. Santa Cruz de Tenerife, Canary Islands (Spain). 27 Jan 1982
Chen P, Fang Z, Chen XJ. Population identification ofOmmastrephes bartramiibased on beak’s external morphological characters. Marine Fisheries, 2015, 35(1):1–9 [陈芃, 方舟, 陈新军. 基于角质颚外部形态学的柔鱼种群判别. 海洋渔业, 2015, 35(1): 1–9]
Clarke MR. The identification of cephalopod “beaks” and the relationship between size and total body weight. Bulletin of the British Museum (Natural History) Zoology, 1962, 8:419–480
Clarke MR. A handbook for identification of cephalopod beaks.Clarendon, Oxford, 1986, 273
Food and Agriculture Organization of the United Nations. Food and Agriculture Organization of the United Nations′ fisheries database-Global capture production from 1950 to 2014. http://www.fao.org/fishery/species/2711/en [联合国粮农组织.联合国粮农组织渔业统计数据––––––1950–2014年全球捕捞产量. http: //www.fao.org/fishery/species/2711/en]
Fang Z, Chen XJ, Lu HJ,et al. Morphological differences in statolith and beak between two spawning stocks forIllex argentinus.Acta Ecologica Sinica, 2012, 32(19): 5986–5997[方舟, 陈新军, 陆化杰, 等. 阿根廷滑柔鱼两个群体间耳石和角质颚的形态差异. 生态学报, 2012, 32(19): 5986–5997]
Fang Z, Chen XJ, Lu HJ,et al. Morphology and growth of beaks in two cohorts for neon flying squid (Ommastrephes bartramii)in the North Pacific Ocean. Acta Ecologica Sinica, 2014a,34(19): 5405–5415 [方舟, 陈新军, 陆化杰, 等. 北太平洋两个柔鱼群体角质颚形态及生长特征. 生态学报, 2014a,34(19): 5405–5415]
Fang Z, Chen XJ, Lu HJ,et al. Progress of beak in cephalopod I:Morphogenesis, biochemistry and growth. Marine Fisheries,2014b, 36(1): 78–89 [方舟, 陈新军, 陆化杰, 等. 头足类角质颚研究进展 I——形态, 结构与生长. 海洋渔业,2014b, 36(1): 78–89]
Fang Z, Liu B, Li J,et al. Stock identification of neon flying squid (Ommastrephes bartramii) in the North Pacific Ocean on the basis of beak and statolith morphology. Scientia Marina, 2014, 78(2): 239–248
García VH. Growth and pigmentation process of the beaks ofTodaropsis eblanae(Cephalopoda: Ommastrephidae). Berliner Palaobiol Abh, Berlin, 2003, 3: 131–140
Gueorguieva R, Krystal JH. Move over anova: Progress in analyzing repeated-measures data and its reflection in papers published in the archives of general psychiatry. Archives of General Psychiatry, 2004, 61(3): 310–317
Hernández-López JL, Castro-Hernández JJ, Hernandez-Garcia V.Age determined from the daily deposition of concentric rings on common octopus (Octopus vulgaris) beaks. Fishery Bulletin-National Oceanic and Atmospheric Administration,2001, 99(4): 679–684
Hu GY, Chen XJ, Fang Z. Effect of individual growth on beak morphometry of jumbo flying squid,Dosidicus gigasoff the Peruvian Exclusive Economic Zone. Journal of Fisheries of China, 2016, 40(1): 36–44 [胡贯宇, 陈新军, 方舟. 个体生长对秘鲁外海茎柔鱼角质颚形态变化的影响. 水产学报, 2016, 40(1): 36–44]
Hu GY, Chen XJ, Liu BL,et al. Microstructure of statolith and beak forDosidicus gigasand its determination of growth increments. Journal of Fisheries of China, 2015, 39(3):361–370 [胡贯宇, 陈新军, 刘必林, 等. 茎柔鱼耳石和角质颚微结构及轮纹判读. 水产学报, 2015, 39(3): 361–370]
Kear AJ. Morphology and function of the mandibular muscles in some coleoid cephalopods. Journal of the Marine Biological Association of the United Kingdom, 1994, 74(4): 801–822
Li CX, Shao Y, Jiang LN. Biostatistics. Beijing: Science Press,2008, 85–110 [李春喜, 邵云, 姜丽娜. 生物统计学. 北京:科学出版社, 2008, 85–110]
Lipiński MR, Underhill LG. Sexual maturation in squid:Quantum or continuum. South African Journal of Marine Science, 1995, 15(1): 207–223
Liu BL, Chen XJ, Zhang CZ. Grey relationship analysis for the environmental factors affecting theNoctiluca scintillansdensity in Qinhuangdao coastal area. Progress in Fishery Sciences, 2010, 31(1): 8–14 [刘必林, 陈新军, 张长征. 印度洋西北海域鸢乌贼角质颚长度分析. 渔业科学进展,2010, 31(1): 8–14]
Liu BW, Pan GP, Zhou WY,et al. CulturedSepiella japonicabeak morphology study. Fisheries Science and Technology Information, 2016, 43 (4): 190–194 [刘本伟, 潘桂平, 周文玉, 等. 养殖日本无针乌贼角质颚形态研究. 水产科技情报, 2016, 43(4): 190–194]
Miserez A, Li Y, Waite JH,et al. Jumbo squid beaks: Inspiration for design of robust organic composites. Acta Biomaterialia,2007, 3(1): 139–149
Olson RJ, Young JW. The role of squid in open ocean ecosystems.Report of a GLOBEC–CLIOTOP/PFRP workshop, 16–17 November 2006, Honolulu, Hawaii, USA. GLOBEC Report,2007, 24: 1–94
Raya CP, Hernández-González CL. Growth lines within the beak microstructure of the octopusOctopus vulgarisCuvier, 1797.South African Journal of Marine Science, 1998, 20(1): 135–142
Smith SC, Whitehead H. The diet of Galapagos sperm whalesPhyseter macrocephalusas indicated by fecal sample analysis. Marine Mammal Science, 2000, 16(2): 315–325
Tang QY, Feng MG. DPS data processing system: Experimental design, statistical analysis and data mining. Science, Beijing,2007
Wang BH. Multivariate statistical analysis and R language modeling. Guangzhou: Jinan University Press, 2010,157–158 [王斌会. 多元统计分析及R语言建模. 广州: 暨南大学出版社, 2010, 157–158]
Yang LL, Jiang YZ, Liu ZL,et al. Variation analysis on partial morphometric measurements of beak ofTodarodes pacificusinhabiting East China Sea. Periodical of Ocean University of China(Natural Science), 2012, 42(10): 51–57 [杨林林,姜亚洲, 刘尊雷, 等. 东海太平洋褶柔鱼角质颚的形态学分析. 中国海洋大学学报(自然科学版), 2012, 42(10):51–57]
Yang Y, Pan BH, Sun XF. Function ofLactobacillus planetariumand its application in animal production. Feed Research,2013(2): 36–37 [杨艳, 潘宝海, 孙笑非. 植物乳杆菌的功能及其在动物生产中的应用. 饲料研究, 2013(2): 36–37]
猜你喜欢
航天工业管理(2020年9期)2020-12-28
军事文摘(2020年14期)2020-12-17
儿童故事画报·智力大王(2020年1期)2020-04-28
航天工业管理(2020年1期)2020-04-20
凤凰生活(2018年6期)2018-06-25
学苑创造·B版(2018年12期)2018-03-04
装甲兵工程学院学报(2017年4期)2017-09-16
家教世界·创新阅读(2016年12期)2017-01-09
阅读与作文(小学高年级版)(2016年1期)2016-03-04
母子健康(2015年8期)2015-12-13
