
运动干预高血压小动脉平滑肌细胞表型转换中miR-143/145对蛋白激酶B信号的调节作用
2018-07-18 02:24廖静雯张琳张严焱叶芳曾凡星石丽君
中国运动医学杂志 2018年2期
廖静雯 张琳 张严焱 叶芳 曾凡星 石丽君
1北京体育大学(北京 100084)
2广州体育学院(广州 510500)
运动对血压的积极调控作用已经得到广泛的研究和认可,各级血管形态和功能都会随长期运动出现适应性变化,相比起大动脉、毛细血管和各级静脉,小动脉即阻力血管的重塑在这个过程中发挥了关键性作用[1,2]。小动脉平滑肌细胞(vascular smooth muscle cell,VSMC)作为动脉血管壁的主要细胞类型,是决定血管反应性和血管重构的重要因素,其增殖、迁移、凋亡以及基质成分合成是高血压、动脉粥样硬化等多种心血管疾病的共同病理基础;VSMC在体内外因素的影响下表现为不同的功能和分化阶段,均涉及VSMC的表型转换。查阅国内外文献发现,运动调控高血压小动脉VSMC表型转换尚未见报道,而多个研究发现运动可以改善动脉粥样硬化大动脉VSMC的功能并可能影响其表型[3-5]。在小动脉的运动适应性重塑过程中,VSMC表型调节可能发挥关键作用,即运动可能通过调节高血压动脉VSMC表型转换来实现对动脉形态和功能的积极影响进而调控血压。
微小RNA(mircroRNA,miRNA,miR)作为重要的表观调控机制之一,为探究先天和后天因素相互作用提供了新契机,对关键miRNA进行研究成为解释高血压的遗传因素与生活方式(如运动)相互作用的良好切入点。miR-143/145在血管壁尤其是VSMC中大量表达,并且miR-143/145在正常血管壁的VSMC及处于分化状态VSMC中表达最多[6];大量离体和在体实验已证实miR-143/145可维持VSMC收缩表型[7,8]。鉴于此,本研究假设长期有氧运动可能影响小动脉中miR-143/145的表达,且其变化趋势与运动后VSMC收缩表型成正相关,而与miR-143/145紧密相关的蛋白激酶B(Pro⁃tein kinase B,PKB,Akt)信号通路也可能在这个过程受到激活。
VSMC中Akt信号通路的激活也对收缩表型的VSMC起正向调控作用[9];肿瘤相关研究报道,miR-143/145靶向于Akt通路上下游多个蛋白的mRNA,两者间呈负向调控关系[10,11]。故本研究推测,miR-143/145可能通过靶向Akt信号通路上多个蛋白来调控VSMC表型转换;在特定的条件下(如高血压),刺激因素(如运动)是否通过miR-143/145引起该通路中某一信号分子变化进而使原本的调节机制被打破,成为本研究待解决的问题。
本实验对成年正常和高血压大鼠分别进行长期有氧运动干预,观察运动改善高血压症状的同时是否可以调节肠系膜动脉VSMC的表型;检测运动干预是否影响动脉血管中miR-143/145的表达,观察Akt信号通路主要蛋白的激活情况,并结合离体实验对肠系膜动脉VSMC分离培养及转染干预,以进行进一步的机制研究。
1 材料与方法
1.1 实验动物与分组
3月龄正常大鼠(Wistar Kyoto rats,WKY)和原发性高血压大鼠(Spontaneously hypertensive rats,SHR)购自北京维通利华实验动物中心,实验动物批号为11400700074310,使用许可证号为SCXK(京)2012-0001,SHR和WKY各分为安静对照组(WKY-C和SHR-C)和运动干预组(WKY-E和SHR-E),每组20只,每笼4只。国家标准啮齿动物饲料喂养,自由饮食和饮水。清洁级动物饲养房温度为20~25℃,相对湿度为45%~55%,昼夜12/12小时照明。
1.2 运动干预方案
各组大鼠先进行1周的适应性训练结束后,运动组进行8周的正式有氧跑台训练(速度为20 m/min,坡度为0,每周5天,每天60 min),运动方案参考文献[12,13]中递增负荷跑台训练及在预实验中对大鼠最大摄氧量的测定,训练强度相当于55%~65%VO2max。安静对照组大鼠于各笼内饲养8周,不进行任何运动干预。分别于正式运动干预前及取材前对各组大鼠进行血压和心率的测量,具体方法参照以往的研究[14]。
1.3 HEHE染色
大鼠麻醉后取肠系膜动脉(2~3级)剥离粘连组织后经固定脱水后溶于石蜡,切片脱蜡后用苏木素和伊红染色,封片并拍摄。每个样本组织切片随机选取10个视野,应用Image J软件进行测量。
1.4 Western blot blot
取肠系膜动脉充分裂解液,采用BCA测试方法对蛋白浓度进行测定。采用SDS-PAGE电泳对蛋白进行分离,将蛋白转膜至PVDF膜上,与一抗(抗体α-actin和OPN购自Abcam;抗体GAPDH和calponin购自San⁃ta Cruz;抗体Akt和p-Akt购自Cell signal)和二抗(均购自Jackson ImmunoResearch)先后结合,采用ECL发光试剂反应液与膜上的目的蛋白反应,凝胶成像系统(Bio-rad,ChemiDoc™ XRS+System)中进行拍摄。利用Image Lab软件对拍摄出的条带影像进行分析。
1.5 实时荧光定量PCRPCR
大鼠肠系膜动脉采用miRcute miRNA提取分离试剂盒(TIANGEN,DP501),按照说明书步骤进行操作,并运用酶标仪对总RNA的纯度和浓度进行检测。反转录采用RevertAid First Strand cDNA Synthesis Kit试剂盒(Thermo Scientific,K1622),按照说明书步骤进行操作。mRNA采用通用引物,miRNA及其对应的内参U6应用特异性茎环引物(miR-145:5’-GTCGTATC⁃CAGTGCAGGGTCCGAGGTATTCGCACTGGATA⁃CGAAGGGATTC-3’;miR-143:5’-GTCGTATCCAGTG⁃CAGGGTCCGAGGTATTCGCACTGGATACGATGAG⁃CTAC-3’;U6:5’-CTCGCTTCGGCAGCACATATACT-3’)进行反转录反应。cDNA扩增采用Power SYBR Green PCR Master Mix试剂盒(Applied biosystems,4367218)进行反应,按照说明书步骤进行操作。胰岛素样生长因子I受体(insulin-like growth factor I re⁃ceptor,IGF-1R)扩增引物为:5’-GATGGAGGAGGTGA⁃CAGGAA-3’和5’-GTGGAGGTGAAACGGAGAAC-3’;胰岛素受体底物(insulin receptor substrate,IRS-1)扩增引物为:5’-AGGCACCATCTCAACAATC-3’和5’-GTTTCCCACCCACCATACTG-3’;p70核蛋白体S6激酶(p70 ribosomal S6 kinase,p70S6K)扩增引物为:5’-GGTGGAGTCTGGGAGCATTA-3’和 5’-TGTGAGG⁃TAGGGAGGCAAAT-3’;GAPDH 扩增引物为:5’-TCCTGCACCACCAACTGCTTAG-3’和 5’-GAT⁃GACCTTGCCCACAGCCTTG-3’。miR-145扩增引物为:5’-GACCAGTCCAGTTTTCCCAG-3’和 5’-TCG⁃TATCCAGTGCAGGGTC-3’;miR-143扩增引物为:5’-ACGAGATGAGATGAAGCACT-3’和 5’-TCGTATC⁃CAGTGCAGGGTC-3’;U6扩增引物为:5’-CTC⁃GCTTCGGCAGCACATATACT-3’和5’-ACGCTTCAC⁃GAATTTGCGTGTC-3’。反应结束后导出数据,获得Ct值,计算△△Ct=(Ct实验组待测基因-Ct实验组参照基因)-(Ct对照组待测基因-Ct对照组参照基因),目的基因的相对表达量为2-△△Ct。
1.6 VSMCVSMC培养与转染
采用酶分离法分离雄性Wistar大鼠(8~12周龄)肠系膜动脉VSMC,方法参照文献[15]。将原代VSMC用含有10%胎牛血清和1%双抗(penicillin/streptomycin)的DMEM培养基培养,置于37℃和5%CO2的培养箱中。当培养皿中VSMC密度达80%~90%时用胰酶(0.25%Trypsin-EDTA)进行消化并传代,VSMC至4~7代且细胞密度达60%~80%时分别进行转染。采用Lipofectamine RNAiMAX 试剂(Invitrogen)将各miRNA干扰序列转染进入细胞内:miR-145 mimic组,miR-145 inhibitor组,空脂质体negative control(NC)组。miR-145 mimic(双链)序列为5’-GUCCAGUUUUCCCAGGAAUCCCU-3’和 5’-GGAUUCCUGGGAAAACUGGACUU-3’;miR-145 inhibitor(单链)序列为5’-AGGGAUUCCUGGGAAAACUGGAC-3’。 蛋白指标于转染后24 h,48 h,72 h和 96 h进行检测,各mRNA于转染后12 h、24 h、48 h和72 h进行定量。
1.7 VSMCVSMC免疫荧光染色与激光共聚焦成像
吸走培养皿中的培养基,VSMC于4%多聚甲醛中固定,用0.2%Triton X-100孵育细胞进行细胞膜穿孔,PBS漂洗后用5%BSA对非特异性结合抗体部位进行封闭。以PBS稀释一抗actin(1︰200),对细胞进行4℃孵育过夜,用PBS漂洗后以PBS稀释二抗(1︰1000)对细胞进行孵育,PBS漂洗后滴加封片剂以盖玻片封片。24 h内运用激光共聚焦成像系统进行信号捕捉与成像。
1.8 统计学分析
数据输入及计算采用SPSS16.0软件。得到的实验数据用平均数±标准差表示;在体实验组间比较采用单因素方差分析,各组间多重比较采用Tukey检验,干预前后各组血压数据比较采用配对样本t检验;离体实验对观测点的多重比较采用LSD方法分析。显著性检验水平以P<0.05表示差异具有统计学意义。
2 结果
2.1 运动干预前后大鼠血压和心率的变化
干预后,WKY-E组大鼠SBP显著小于WKY-C组(P<0.05);SHR-E组SBP非常显著小于SHR-C组(P<0.01);SHR-E组SBP与8周运动前比也显著下降(P<0.05);SHR-E组DBP和MAP也显著低于SHR-C组(P<0.05)。见表1。
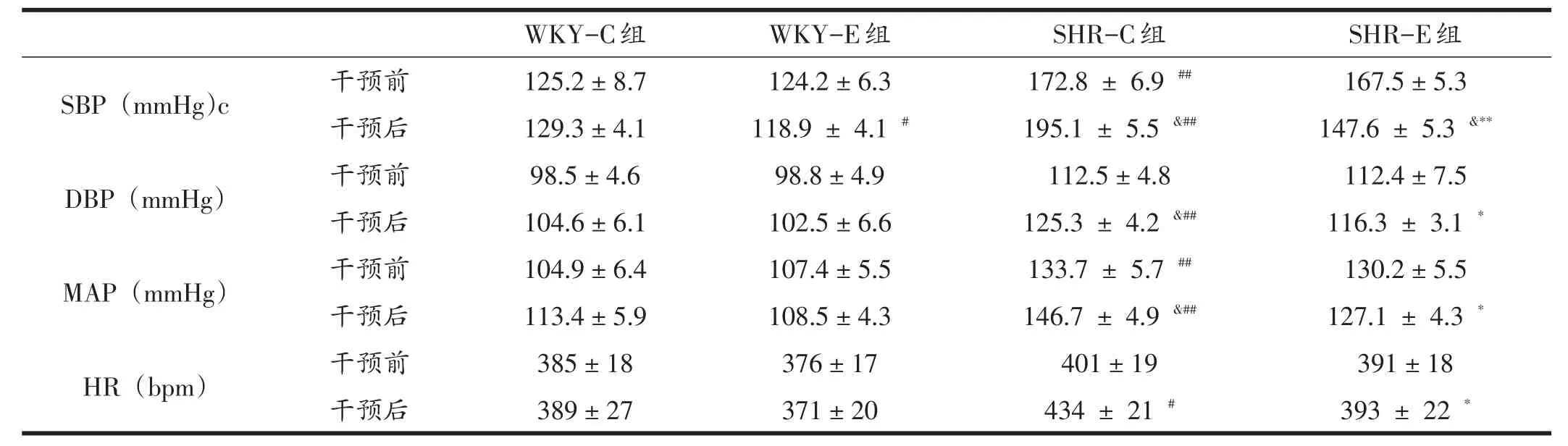
表1 各组大鼠血压和心率比较(n=20)
2.2 运动干预后大鼠肠系膜动脉形态学变化
WKY-E组与WKY-C组大鼠肠系膜动脉内径间无显著差异,SHR-E组动脉内径与SHR-C组相比有升高的趋势但无显著差异;WKY-E组与WKY-C组动脉壁厚无显著差异,SHR-E组动脉壁厚显著小于SHR-C组(P<0.05);动脉壁厚/内径比值,虽然SHR-E组相比SHR-C组下降但未见显著差异。见图1。
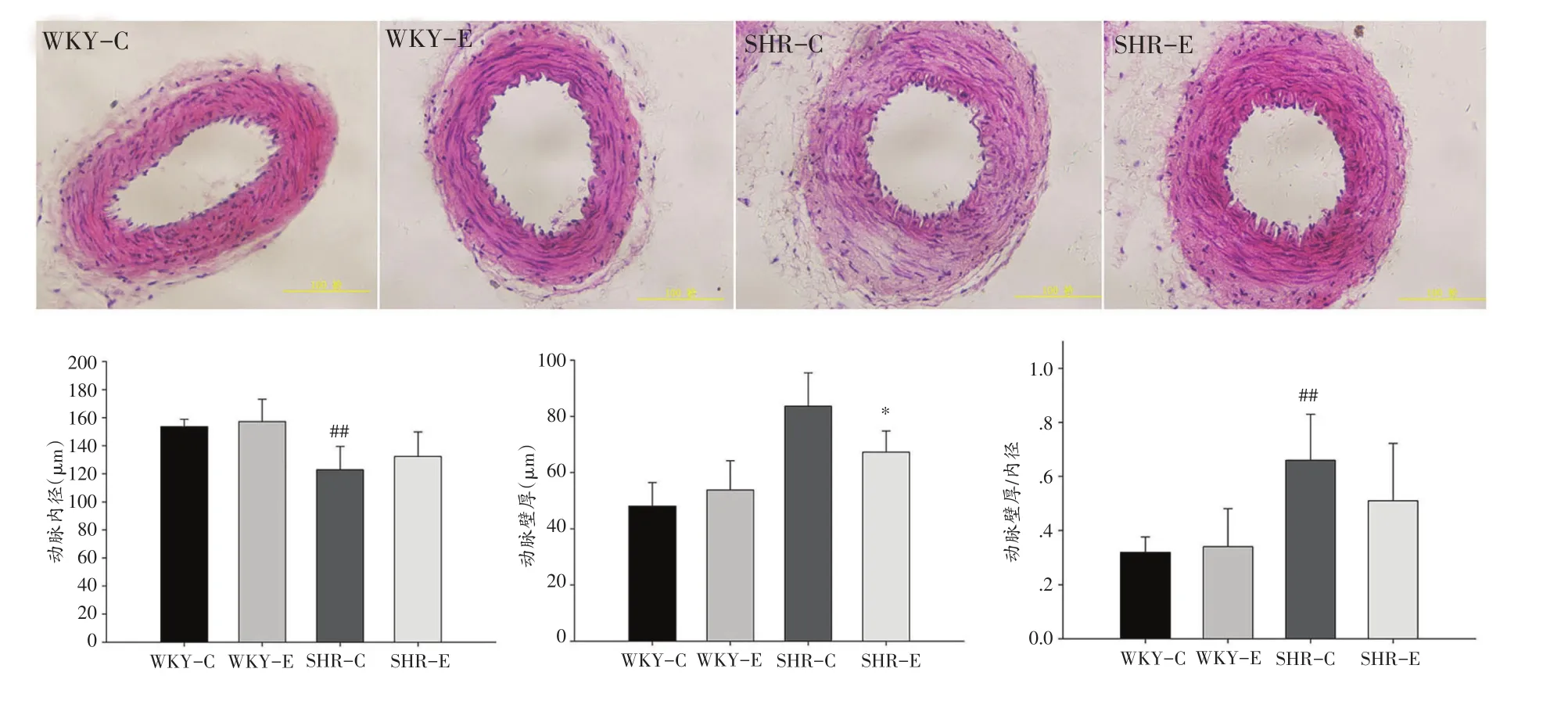
图1 运动对肠系膜动脉形态的影响(n=6)
2.3 运动干预后大鼠肠系膜动脉VSMCVSMC表型标志蛋白的变化
α-actin在WKY-C组和WKY-E组间无显著差异,SHR-C组α-actin表达显著低于WKY-C组,而SHR-E组α-actin表达与SHR-C比虽然有所升高但未见显著性差异(图2A)。WKY-E组calponin表达与WKY-C比未见显著性差异,SHR-C组与WKY-C组比其calponin表达显著降低,SHR-E组calponin表达显著高于SHRC组(P<0.05)(图2B)。WKY-C组和WKY-E组OPN呈现低表达且两组间未见显著性差异,SHR-C组OPN表达显著高于WKY-C组,SHR-E组OPN显著低于SHRC组(P<0.05)(图2C)。
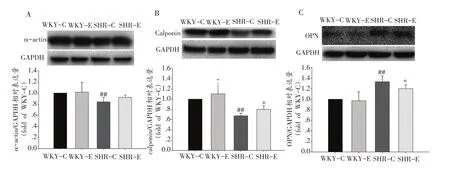
图2 运动对平滑肌表型标志蛋白的影响(n=12)
2.4 运动干预后大鼠肠系膜动脉miR-miR-143143和miR-miR-145145表达的变化
WKY-C组和WKY-E组miR-145表达无显著差异,SHR-C组miR-145表达显著低于WKY-C组(P<0.01),SHR-E组miR-145表达显著高于SHR-C组(P<0.01)。miR-143在WKY-C和WKY-E间也未见显著差异,SHR-C组miR-143表达显著低于WKY-C组(P<0.01)。见图3。
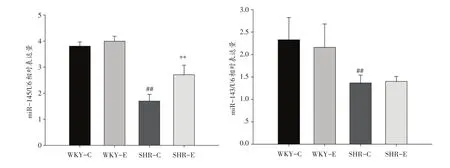
图3 运动对肠系膜动脉miR-145和miR-143表达的影响(n=12)
2.5 运动干预后大鼠肠系膜动脉AktAkt信号通路的激活
SHR-C组Akt磷酸化程度较WKY-C被显著抑制(P<0.01),SHR-E组Akt信号活性增强,与SHR-C比有显著性差异(P<0.05)(图4)。
2.6 运动干预后VSMCVSMC经转染后α -actinactin表达的变化
与原代分离VSMC相比,经过传代后的VSMC内α-actin表达减少且呈现的丝状结构也减少,细胞呈多边形;VSMC经转染miR-145 mimic后,miR-145过表达使VSMC中α-actin表达明显增多且细胞呈梭形,甚至出现了α-actin的丝状结构,表现在其收缩功能结构在形态上增强;VSMC经转染miR-145 inhibitor后VSMC依然保持较低的α-actin表达量,细胞结构和形态不明显,没有明显细胞骨架结构(图5A)。
VSMC经转染miR-145 mimic后收缩表型标志蛋白α-actin在12 h显著增加(P<0.01),在24 h达到峰值,与NC组比有非常显著性差异(P<0.01),在48 h后恢复正常水平;VSMC经转染miR-145 inhibitor后αactin表达在24 h显著降低(P<0.01)后逐渐恢复。VSMC经转染干扰miR-145表达后,其收缩表型标志蛋白α-actin表达均有明显地改变(图5B)。
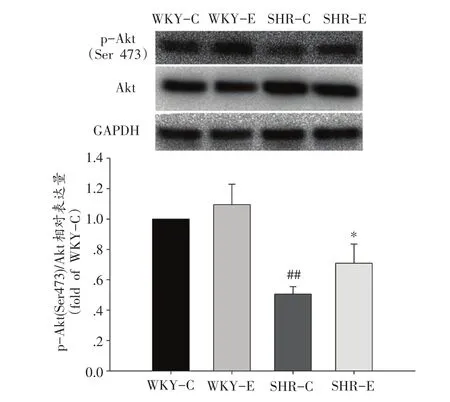
图4 运动对肠系膜动脉Akt磷酸化的影响(n=12)
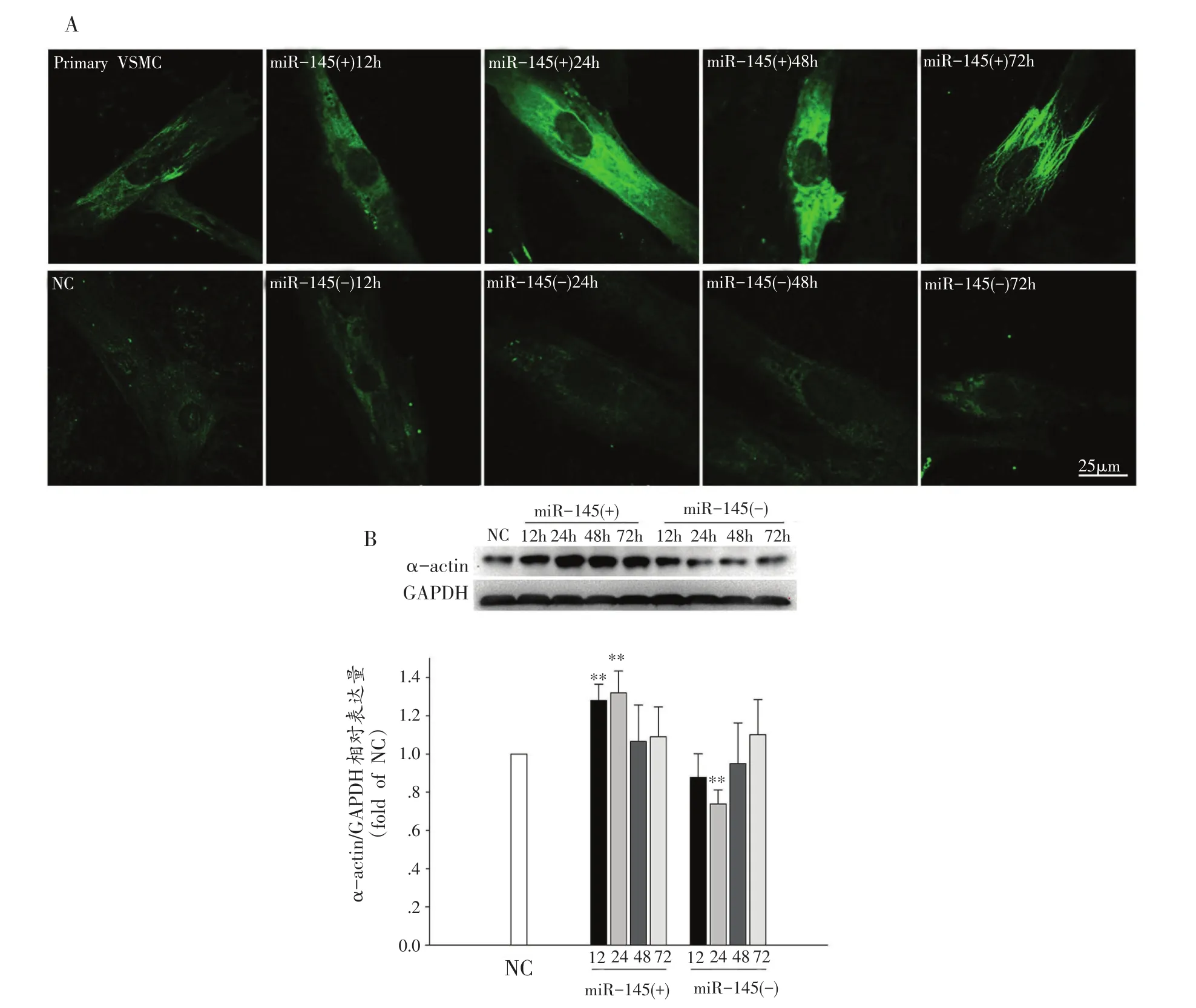
图5 VSMC经转染后α-actin的相对表达情况(n=3)
2.7 VSMCVSMC经转染后AktAkt磷酸化表达变化
VSMC经转染miR-145 mimic使之过表达后显著抑制了Akt磷酸化(P<0.01),使Akt在12 h即非常显著降低,在24 h达到最低值后抑制效果持续至48 h,在72 h Akt磷酸化程度恢复正常;VSMC经转染miR-145 inhibitor后,Akt受miR-145的抑制效果被显著解除,Akt活性在转染后12 h显著升高(P<0.05),持续到48 h其活性出现峰值后到72 h恢复正常水平。VSMC经转染干扰miR-145后,Akt活性具有显著性变化,miR-145与Akt呈明显的负向调控关系(图6A)。
2.8 VSMCVSMC经转染后AktAkt下游分子 mRNAmRNA表达变化
VSMC经转染miR-145 mimic后miR-145的过表达显著抑制了IGF-1R mRNA的含量,在12 h出现最低值后逐渐恢复,在48 h依然显著低于正常水平(P<0.01);IRS-1 mRNA表达在12 h受抑制显著降低(P<0.01),后逐渐恢复,在24 h达到正常水平;而p70S6KmRNA在48h出现显著下降并与NC比有非常显著差异(P<0.01),后恢复正常水平(图6B、C、D)。
VSMC经转染miR-145 inhibitor后,IGF-1R mRNA表达在24 h显著升高(P<0.01),IRS-1 mRNA表达显著升高(P<0.01),在24h显著高于NC并在72h达到峰值,p70S6KmRNA表达在72 h内未出现显著改变(图6)。
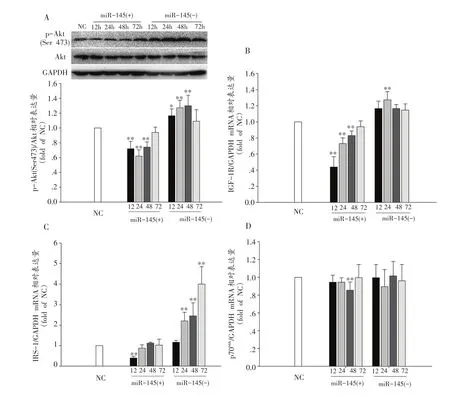
图6 VSMC经转染后Akt磷酸化及其下游信号分子mRNA表达情况(n=3)
3 讨论
3.1 运动对血压和小动脉血管形态的影响
中低强度长期有氧运动常作为干预高血压的方法,其有效性、安全性、低成本和可实施性在研究和实际运动中也最为广泛。本实验采用8周中等强度跑台训练对正常大鼠和高血压大鼠进行运动干预并观察其血压变化,结果显示运动干预组大鼠与运动前比SBP显著下降,与安静组比SBP、DBP和MAP均下降,即该有氧运动可以显著缓解SHR的高血压症状。
小动脉的重构在运动干预高血压中发挥了关键性的作用,长期运动后小动脉形态通过一系列适应性变化降低了外周阻力并增加了血管的反应性而最终起到降低血压的作用。本研究选取大鼠肠系膜动脉为组织材料进行研究,因其为小动脉肌性阻力血管,中膜平滑肌细胞丰富,符合本研究的实验目的。本结果中高血压运动组大鼠肠系膜动脉形态未出现明显适应性变化,其中动脉内径有所增大但并不显著,壁厚显著下降,动脉壁厚与内径的比值也未有显著差异,说明本实验中有氧运动可以使小动脉形态发生改变但并不显著。以往研究中,高血压动物模型经过长期运动后动脉血管形态有明显改变或血管并未发生重构均有报道[13],其原因可能为:运动所致的血管形态变化是一个循序渐进积累的过程,细胞功能的改变可能早于血管的整体形态;研究表明,运动可有效改变肠系膜动脉的收缩功能[16,17]。另外,由于肠系膜动脉由腹腔动脉分支后继续分支为各级动脉,这一因素使样本取材过程中存在一定的误差,这也是导致各研究结果差异的原因。尽管大部分研究支持运动员动脉血管内径与缺少体力活动的人有差异[18,19],然而不同的运动模式(如耐力运动、力量练习或两者结合锻炼)是否会影响动脉的形态变化尚不清楚,这可能是由于不同的动脉血管检测部位及不同的运动模式造成的。各级血管对于长期运动的适应性重构具有较大差异;并且,血管重构更易发生于外周小动脉及常暴露于血流和剪切力变化的血管中。
3.2 运动对小动脉VSMCVSMC表型转换的影响
机体各级动脉的结构特性相差很大,中小动脉为肌性动脉,管壁内弹性胶原逐渐减少而VSMC较多,细胞外基质相对较少,发挥了控制血压和调节血液分配的作用。小动脉因体内外因素发生重构的过程中,VSMC形态和功能的变化(如表型转换)对血压的调节可能发挥了重要作用。本研究显示,运动对正常血压大鼠肠系膜动脉VSMC表型未见显著性影响,正常和高血压大鼠动脉中VSMC表型有显著差异,而运动可以有效调节VSMC表型,促进SHR大鼠动脉VSMC维持收缩表型。查阅文献发现,运动调节高血压小动脉VSMC表型尚未见报道;在相关心血管病症的运动干预后,运动确实调节了动脉VSMC表型。Jordao等研究发现,12周低强度跑台训练可以使SHR胸主动脉I型和III型胶原纤维mRNA表达降低,防止血管内弹性膜破裂;运动还使大动脉的形态、α-actin、弹性蛋白和胶原蛋白mRNA表达趋于正常,使VSMC在血管中定向分布[4]。Wamhoff等[5]对雌性小型猪进行16周递增跑台实验,发现长期的有氧运动可以减弱有丝分裂原引起的冠状动脉VSMC表型转换即保持具有维持血管收缩功能的收缩表型。Simmons等[20]对高胆固醇血症的猪实验模型进行16~20周的中等强度有氧训练,其通过对Akt、HSP90、Catalase等进行蛋白含量检测结合判断VSMC表型,发现其冠状动脉中VSMC表型未见显著性改变。Bowles等[21]证明运动可以改善冠状动脉VSMC中钙离子依赖的基因转录并激活细胞膜钾离子通路,维持VSMC稳定的收缩表型。以上结果的差异可能是由于所选择的判定蛋白的不同所导致的;另外,不同动物模型及干预方式不具备可比性且检测血管部位的不同都是造成结果差异的因素。综上所述,运动对高血压小动脉VSMC表型具有调节作用,维持其具有功能性的收缩表型。
3.3 运动干预VSMCVSMC表型中miR-miR-145145对AktAkt信号通路的调控关系
miRNA已在高血压和VSMC表型间建立了联系,但文献中对于miRNA在运动调节高血压血管功能中的作用鲜有报道,运动对VSMC表型转换中关键miR⁃NA的影响尚属空白。miR-143和miR-145作用共享一个初始转录本,发挥类似的作用,故许多研究中也把两者miR-143/145放在一起讨论。动物和离体实验均证明miR-143/145可以使VSMC维持收缩表型,促进其分化并抑制其增殖[22,23]。本研究显示,运动可以改变高血压大鼠肠系膜动脉中miR-145的表达并促进其增加,而miR-143表达未明显受到运动的影响,miR-145对于运动的适应性表达变化可能比miR-143更加灵敏。运动后miR-145表达的升高与小动脉中VSMC收缩表型标志蛋白的升高一致,说明运动引起的miR-145的改变可能参与了VSMC收缩表型的维持。以往研究也显示,miR-143在调节VSMC表型转换中的作用不如miR-145明显,主要表现为miR-143基因敲除小鼠较miR143/145及miR-145基因敲除小鼠血管结构和收缩功能改变并不明显[24,25];另外,miR-145在诱导多能干细胞向VSMC分化过程中发挥重要调节作用,而miR-143并没有明显作用[26]。
Akt是一种丝氨酸/苏氨酸蛋白激酶,参与调节细胞增殖、分化、凋亡;Akt位于PI3K/Akt信号转导通路的枢纽部位,是该通路下游的一种重要效应分子。大量研究已证明,VSMC在保持分化状态即维持其收缩表型中依赖于Akt通路,且其丝氨酸473位点(Ser 473)磷酸化程度决定了其激活状态并与VSMC分化标志蛋白成正相关[9];高血压动物实验中也证明Akt信号的激活可以使VSMC保持分化,即维持VSMC的收缩表型[27]。在Akt信号通路的上下游存在着反馈调节,如在胰岛素敏感的组织中(如骨骼肌和脂肪细胞),Akt下游mTOR和p70S6K可以通过抑制Akt上游IRS-1磷酸化来对该通路进行反馈调节[28,29],用雷帕霉素抑制mTOR和p70S6K的表达可以释放其对IRS-1的抑制从而激活Akt信号,这样的调节机制在糖尿病患胰岛素抵抗[30]和VSMC表型转换[31]中都得到了证实。长期运动影响血管中Akt表达在文献中只有少量报道,Li等[32]发现12周抗阻训练可以激活大鼠主动脉中Akt信号的活性,起到对抗氧化应激和保持血管的作用;Wang等[33]在心肌缺血所致的外周血管阻力增大模型中发现,8周跑台运动可以使大鼠肠系膜动脉中PI3K、Akt等信号激活,使该通路活性趋于正常。然而,Akt在长期有氧运动后对小动脉VSMC的作用尚未见报道。本研究显示,高血压大鼠肠系膜动脉中Akt信号活性被明显抑制,而运动可以激活动脉中Akt信号的活性,高血压小动脉中Akt信号在运动后被激活与VSMC收缩表型的维持和miR-145表达的提高相一致。然而,在癌症肿瘤相关研究中,miR-145与Akt信号通路存在负调控关系[34-36],故在各组织中miR-145与Akt信号通路可能形成网络式调节来发挥重要的生理与病理功能。
在心血管疾病中,miR-143/145对VSMC表型及Akt的研究只有个别报道,如Riches等离体培养VSMC发现,转染pre-miR-143/145可以使Akt上游IGF-1R蛋白表达增加,然而抑制miR-143/145并不会使IGF-1R减少[8],推测miR-143/145对IGF-1R的影响可能受到其它反馈调节的控制。在不同病理情况下的病理组织中(如癌症高血压、动脉粥样硬化、糖尿病等),miR-143/145可能呈现截然不同的表达及调节机制。在VSMC中miR-143/145与Akt信号间尚未明确建立调节关系,由于miRNA的多靶点效应和Akt信号通路的网络式反馈调节使得两者在VSMC生理和病理过程中的相互作用更为复杂;miR-143/145可能同时作用于Akt上下游信号分子的mRNA,而Akt信号的一个效应分子的激活或抑制可能被其它效应分子的作用所掩盖或代偿。
故本研究选择在离体培养的VSMC中对miR-145进行干扰,明确其与Akt信号的调节关系。转染miR-145 mimic使miR-145在增殖至4~7代的VSMC中过表达,VSMC中收缩表型标志蛋白表达增加而合成表型标志蛋白表达减少,且转染后的细胞呈梭形;而转染miR-145 inhibitor后VSMC中α-actin表达减少,VSMC向合成形态转换。α-actin作为重要的VSMC表型标志蛋白和细胞骨架功能蛋白,其高表达对于肌细胞保持收缩能力是必须的;Xin等[24]证明,敲除Dicer的小鼠由于全身性缺乏mRNA使其VSMC中actin整体骨架蛋白严重受损,而miR-145在小鼠内过表达后可以明显缓解其VSMC中骨架蛋白的受损,这个过程与miR-145参与actin的动态调节密切相关。
本研究结果还显示,在VSMC中miR-145过表达分别抑制了IGF-1R和IRS-1 mRNA,同时Akt磷酸化程度降低;当VSMC中miR-145表达减少时,IGF-1R和IRS-1 mRNA的抑制作用解除并出现高表达,同时Akt活性也被激活。说明miR-145可分别靶向于Akt信号通路上游IGF-1R和IRS-1 mRNA,通过抑制其表达进一步抑制Akt信号的激活;然而,运动干预后高血压动脉VSMC中miR-145和Akt均被激活,推测运动后miR-145维持VSMC的收缩表型可能并不经过Akt通路的激活。另外,研究证实Akt在调节VSMC表型的过程中其上下游间存在着反馈调节的关系,即Akt上游IGF-1R和IRS-1被磷酸化激活后可以使Akt活性增强进而激活mTOR/p70S6K,然而p70S6K的过度活化可以反过来抑制IGF-1R和IRS-1信号向下转导,故而形成一个反馈环[31]。在有限的VMSC相关研究中,Riches等对从二型糖尿病患者隐静脉分离的VSMC进行培养发现,转染pre-miR-145可以使IGF-1R蛋白表达增加,然而抑制miR-145并不会使IGF-1R减少[8]。人主动脉平滑肌细胞株研究中,Akt通路的阻断剂并不能减轻牵张力所致的miR-145抑制,即牵张力并不通过Akt通路发挥其对miRNA的抑制作用[23]。结合本研究结果及以往研究报道中miR-145对Akt通路的靶向关系可以推测,miR-145与Akt通路的相互关系可能受到复杂反馈调节的控制。在不同的体内外因素刺激下(如高血压、动脉粥样硬化、糖尿病、运动等),二者可能呈现截然不同的表达及调节机制,此方面的研究需要进一步探索。
4 结论
运动改善高血压大鼠血压症状的同时使小动脉结构适应性变化;运动有助于维持小动脉中VSMC收缩表型。miR-143/145可能参与了运动导致的VSMC表型适应性变化。miR-145对Akt呈负向调节,可能参与了VSMC表型转换过程中Akt信号通路的反馈调节。运动后Akt的激活并不是miR-145过表达导致的,提示运动可能通过其它途径激活Akt信号并促进VSMC向收缩表型转换。
猜你喜欢
山东医药(2022年3期)2023-01-06
河北果树(2021年4期)2021-12-02
天津医科大学学报(2021年1期)2021-01-26
医药前沿(2020年20期)2020-11-10
医学新知(2019年4期)2020-01-02
河北农业科学(2019年6期)2019-03-21
中国实用乡村医生杂志(2019年7期)2019-01-08
健康必读·下旬刊(2018年5期)2018-06-06
腹腔镜外科杂志(2016年10期)2016-06-01
腹腔镜外科杂志(2016年10期)2016-06-01
