
烟草甲两个羧酸酯酶基因的克隆及表达模式*
2018-08-08 05:10朱晓晔丁天波许抗抗周江李灿杨文佳
中山大学学报(自然科学版)(中英文) 2018年4期
朱晓晔 ,丁天波,许抗抗,周江,李灿 ,杨文佳
(1. 贵阳学院生物与环境工程学院∥贵州省山地珍稀动物与经济昆虫重点实验室,贵州 贵阳 550005; 2. 贵州师范大学生命科学学院,贵州 贵阳 550001; 3. 青岛农业大学植物医学学院,山东 青岛 266109)
烟草甲Lasiodermaserricorne(Fabricius),隶属鞘翅目Coleoptera、窃蠹科 Anobiidae,是一种世界性的仓储害虫,在中国大部分地区都有分布。该虫寄主范围广,对烟叶、粮食、茶叶、中药材等多种储藏物造成危害[1]。主要以幼虫潜居寄主体内进行蛀食,排出大量粪便、留下虫尸、产生异味,严重影响储藏物的产量和品质,其发生危害具有较强的隐蔽性[2]。目前烟草甲的防治主要以化学药剂熏蒸为主,不仅带来食品安全和人类安全隐患,而且导致其对杀虫剂产生了不同程度的抗药性,需要探索新的仓储害虫防治策略[3-4]。气调技术(利用高φ(CO2)和低φ(O2)协同作用)作为一种新的防治手段,近年来被广泛应用于仓储害虫的防治[5],但目前已在谷象Sitophilusgranaries、赤拟谷盗Triboliumcastaneum、粉斑螟Ephestiacautella、嗜卷书虱Liposcelisbostrychophila等发现其对CO2气调产生了不同程度的抗性[6-10]。前期研究对昆虫应对CO2气调胁迫适应性主要集中在生理学和生态学方面,而分子水平上的机制研究尚未见报道。
羧酸酯酶(carboxylesterase, CarE)是一种多功能家族酶,广泛存在于微生物、植物、昆虫和哺乳动物中。CarE属于丝氨酸水解酶家族,具有多种类型,有广泛的底物专一性和重迭性,大多数酶的反应机理相同,因为在其氨基酸序列上存在保守的催化三联体活性中心(Ser、His、Glu)。在昆虫体内,CarE的生理功能并不局限在对外源化合物的解毒代谢和对杀虫剂的抗药性形成中的作用[11-12],近年来研究发现它在昆虫信息素、激素降解以及调节昆虫行为中起着重要作用。此外,部分CarE与昆虫神经形成和发育调控密切相关。本课题组前期研究发现,高体积分数CO2气调胁迫后烟草甲体内CarE酶活性显著升高[17],推测CarE可能在害虫应对CO2气调胁迫过程中发挥着重要作用。
为了探究CarE基因在烟草甲体内的生理功能,本研究以转录组测序获得的CarE序列为基础,利用RT-PCR技术克隆获得烟草甲2条CarE基因的cDNA全长序列,并对其氨基酸序列、同源性和系统发育进行生物信息学分析,同时对其在不同发育阶段及CO2气调胁迫下的表达模式进行分析,旨在分析CarE基因与烟草甲生长发育及与CO2气调胁迫响应的相关性,为理解该虫生长发育调控及气调胁迫下的应激机制提供分子基础。
1 材料和方法
1.1 供试昆虫
供试烟草甲于2010年采自贵州省贵阳市,置于温度为(28±1) ℃,相对湿度为(75±5) %,全黑暗的人工气候箱内,以中药材当归Angelicasinensis为食料连续饲养繁殖40代以上。
1.2 主要试剂
TRIzol试剂(TRIzol regent,Invitrogen公司);DNA酶(RQ1 RNase-Free DNase,Promega公司);反转录试剂盒(PrimeScript®RT Reagent Kit,TaKaRa公司);Taq酶(TaKaRa公司);胶回收试剂盒(Gel Extraction Kit,Omega公司);载体(pGEM-T Easy Vector,Promega公司);感受态细胞DH5α(北京全式金公司);定量试剂(GoTaq®qPCR Master Mix,Promega公司)。
1.3 试验方法
1.3.1 烟草甲RNA提取 分别收集烟草甲低龄幼虫、高龄幼虫、蛹和成虫,每个虫态设置4个生物学重复,每个重复40头试虫。按照TRIzol试剂说明书提取上述样品总RNA,利用Nanodrop 2000核酸浓度测定仪和琼脂糖凝胶电泳检测总RNA的浓度和完整性。
1.3.2 CO2气调处理 参照笔者实验室已发表的CO2气调毒理的生物测定数据[21],挑取高龄幼虫于2个亚致死体积分数LC10[φ(CO2)=30% +φ(空气)=70%]和LC30[φ(CO2)=70% +φ(空气)=30%],及致死中体积分数LC50[φ(CO2)=90%+φ(空气) =10%]的CO2气调处理6 h,挑取存活的试虫备用,相同条件下空气处理作为对照,每组处理设置4个重复。将处理后的试虫按照1.3.1方法分别提取总RNA。
1.3.3 第一链cDNA的合成 利用DNA酶处理总RNA,去除基因组DNA后,按照PrimeScript®RT Reagent Kit说明书合成第一链cDNA,作为全长验证PCR扩增和实时定量PCR的模板。
1.3.4 基因克隆 根据笔者实验室测序完成的烟草甲转录组数据中获得的2个CarE基因片段,BLAST分析后,采用Primer 5.0软件设计2对特异性引物(表1)用于开放阅读框的扩增。PCR反应体系包含10×PCR缓冲液2.5 μL,MgCl22.5 μL,dNTPs 2 μL,上下游引物各1 μL,cDNA模板2.0 μL,rTaq酶0.25 μL,ddH2O补足至25 μL。扩增条件为:95 ℃预变性3 min,然后95 ℃变性30 s,58 ℃ 退火30 s,72 ℃延伸2 min,共35个循环,最后72 ℃延伸10 min。PCR产物经w=1%琼脂糖凝胶电泳检测后,按照Gel Extraction Kit说明书回收目的条带,连接至pGEM-T Easy上,再转化至DH5α感受态细胞中,经蓝白斑筛选和PCR鉴定,最终将阳性克隆送往成都擎科梓熙生物技术有限公司进行测序。

表1 本研究中使用的引物Table 1 Primers used in this study
1.3.5 序列分析 利用DNAMAN 6.03(Lynnon Biosoft)软件对测序结果进行编辑和分析,推导的氨基酸采用BLAST工具(http:∥www.ncbi.nlm.gov/BLAST/)进行同源性比较分析。利用ProtParam(http:∥web.expasy.org/)、NetNGlys 1.0 server(http:∥www.cbs.dtu.dk/services/NetNGlyc/)和SignalP 4.1(http:∥www.cbs.dtu.dk/services/SignalP/)分析编码蛋白的理化性质、N-糖基化位点和信号肽。利用Scanprosite(http:∥prosite.expasy.org/scanprosite/)分析氨基酸的保守区域。利用MEGA 7.0软件中的邻接法(Neighbor-joining)构建系统发育树,各分支重复检验次数均为1 000[18]。
1.3.6 实时定量PCR检测 采用实时定量PCR方法检测烟草甲LsCarEB1和LsCarEB2在不同发育阶段及CO2气调胁迫后的相对表达量。使用在线软件Primer 3.0(http:∥frodo.wi.mit.edu/)设计特异性表达引物,内参基因采用烟草甲LsEF1α基因(GenBank登录号:KY549658),引物序列信息如表1。实时定量PCR反应体系如下:GoTaq®qPCR Master Mix 10 μL、cDNA 1 μL、上下游引物各1 μL以及Nuclease-free Water 7 μL。扩增条件:95 ℃预变性5 min;然后95 ℃变性30 s,60 ℃退火30 s,共40个循环;最后在60~95 ℃ 进行熔解曲线分析。每个样品重复测定3次。利用2-△△CT方法计算不同发育阶段及CO2气调胁迫后的基因相对表达量[19],试验数据采用SPSS23.0软件中的单因素方差分析(ANOVA)进行差异显著性分析。
2 结果与分析
2.1 LsCarEB1和LsCarEB2的克隆及序列分析
利用RT-PCR技术扩增出烟草甲LsCarEB1和LsCarEB2的cDNA全序列,将其提交到NCBI核酸数据库,GenBank登录号分别为MG189601和MG189602。LsCarEB1开放阅读框长度为1 698 bp,编码565个氨基酸(图1),理论相对分子质量和等电点分别为64 100和6.40。LsCarEB2的开放阅读框为1 695 bp,编码564个氨基酸(图2),理论相对分子质量和等电点分别为64 000和5.32。经NetNGlyc 1.0 server预测,LsCarEB1可能存在3个N-糖基化位点,分别为N117、N250和N376;LsCarEB2存在2个N-糖基化位点,分别为N359和N371。 经ScanProsite分析表明,2个推导的蛋白均具有羧酸酯酶典型的两个保守结构域:LsCarEB1蛋白第199-214位含羧酸酯酶B型丝氨酸活性位点FGGNPNSVTLTGFSAG,第104-110位含二硫键形成位点EDCLYLN;LsCarEB2蛋白第197-212位含丝氨酸活性位点FGGNPDSITLTGMSAG,第104-110位含二硫键形成位点EDCLYVN。LsCarEB1蛋白催化活性中心是以丝氨酸(Ser212)为中心的催化三联体(Ser212、Glu346和His466),LsCarEB2蛋白的催化三联体为(Ser210、Glu339和His461)。LsCarEB2编码的蛋白N末端具有20个氨基酸组成的信号肽序列,推测其可能为分泌蛋白,而LsCarEB1无信号肽存在。
将LsCarEB1和LsCarEB2编码的氨基酸序列分别进行BLAST比对分析,结果发现,LsCarEB1与同为鞘翅目的赤拟谷盗T.castaneum(GenBank登录号:CAH64510)和黄粉虫Tenebriomoliter(AKZ17664)羧酸酯酶的同源性较高,分别为 51% 和44%。LsCarEB2与赤拟谷盗T.castaneum(KYB968215)和黄粉虫T.moliter(AKZ17669)羧酸酯酶的同源性分别为 56% 和53%。另外,LsCarEB1和LsCarEB2编码的蛋白质序列相似性为30.0%。
2.2 LsCarEB1和LsCarEB2的系统发育分析
将LsCarEB1和LsCarEB2推导的氨基酸序列与GenBank登录的其它昆虫CarEs氨基酸序列进行比对,利用MEGA 7.0软件的邻接法构建进化树。结果表明,上述昆虫羧酸酯酶可分为6个簇(A、F、G、K、M和N),本文克隆获得的LsCarEB1和LsCarEB2基因均属于羧酸酯酶第三功能组(H-N)的M簇中,该簇主要为β酯酶(β-esterase)(图3)。进一步分析发现它们与同为鞘翅目的马铃薯甲虫Leptinotarsadecemlineate和赤拟谷盗T.castaneum的亲缘关系较近,与传统分类学的分类一致。
2.3 LsCarEB1和LsCarEB2在不同发育阶段的表达
利用实时定量PCR方法检测LsCarEB1和LsCarEB2在烟草甲不同发育阶段的表达情况。结果表明,LsCarEB1和LsCarEB2在烟草甲各发育时期均有表达,其中LsCarEB1在高龄幼虫、成虫及低龄幼虫中的相对表达量显著高于蛹期(P< 0.05),上述时期的mRNA表达量分别是蛹期的14.9、3.58和2.50倍。LsCarEB2在高龄幼虫、成虫期、低龄幼虫中的表达量分别是蛹期的6.92、2.76和1.21倍,且在高龄幼虫期的表达量显著高于其他时期(P< 0.05)(图4)。
2.4 不同φ(CO2)对LsCarEB1和LsCarEB2基因表达的影响
采用LC10、LC30和LC503个φ(CO2)处理烟草甲高龄幼虫,6 h后发现LsCarEB1在LC10的相对表达量与对照组相比无明显差异,而LC30和LC50处理组mRNA表达量分别为对照组的2.7倍和4.1倍,且显著高于对照组(P< 0.05)。LsCarEB2 mRNA的相对表达量分别为对照的1.66、1.89和2.65倍,其表达量随胁迫CO2的体积分数升高而上调,LC50气调胁迫下mRNA表达量显著高于对照、LC10和LC30处理组(P< 0.05)(图5)。
3 讨 论
羧酸酯酶作为昆虫体内重要的初级代谢酶系,在杀虫剂及外源物质的代谢、激素降解和神经发育过程中发挥着重要的作用[11-14]。随着昆虫基因组计划的开展、转录组测序技术不断普及,目前已有多种昆虫的羧酸酯酶基因被陆续克隆,如黑腹果蝇Drosophilamelanogaster[22]、冈比亚按蚊Anophelesgambiae[23]、赤拟谷盗T.castaneum[24]、家蚕Bombyxmori[25]、意大利蜜蜂Apismellifera[26]、豌豆蚜Acyrthosiphonpisum[27]、温室白粉虱Trialeurodesvaporariorum[28]、中华稻蝗Oxyachinensis[29]、橘小实蝇Bactroceradorsalis等[30]。根据其催化能力、细胞定位和系统进化,可将昆虫羧酸酯酶分为14个簇(命名为A-N簇),进一步可划分为A-C、D-G、H-N 3个功能组[31]。本研究在烟草甲转录组测序基础上,通过RT-PCR技术克隆获得2条CarE基因cDNA全序列,经与其它昆虫CarE氨基酸进行序列比对和聚类分析,发现LsCarEB1和LsCarEB2均属于M簇,为β酯酶。前期研究发现M簇CarEs基因参与昆虫体内多种生理途径,包括信息素的降解、生殖发育以及对外源物质的解毒代谢等[28]。

图1 烟草甲LsCarEB1核苷酸序列及推导的氨基酸序列(起始密码子ATG和终止密码子TAA加粗;*表示终止密码子;下划虚线为信号肽序列;下划线为糖基化位点;催化三联体用阴影表示;方框为保守氨基酸序列)Fig.1 Nucleotide and deduced amino acid sequence of LsCarEB1 in Lasioderma serricorne (The start codon is indicated with bold and the stop codon is indicated with bold and an asterisk; N-linked glycosylation is underlined; Putative catalytic triad is shaded; The conserved amino acids are boxed)
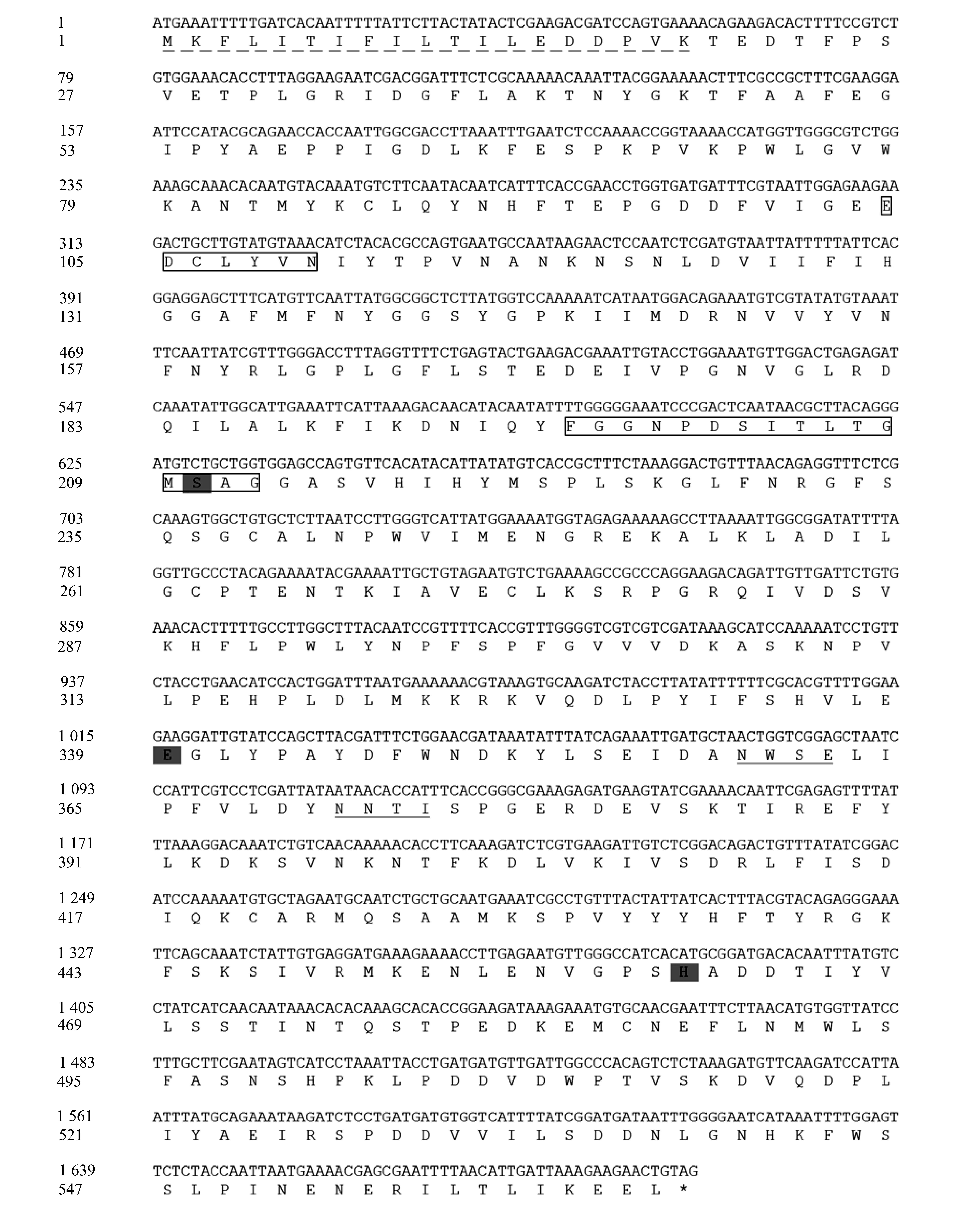
图2 烟草甲LsCarEB2核苷酸序列及推导的氨基酸序列(起始密码子ATG和终止密码子TAA加粗;*表示终止密码子;下划虚线为信号肽序列;下划线为糖基化位点;催化三联体用阴影表示;方框为保守氨基酸序列)Fig.2 Nucleotide and deduced amino acid sequence of LsCarEB2 in Lasioderma serricorne(The start codon is indicated with bold and the stop codon is indicated with bold and an asterisk; The predicated signal peptide is dotted underlined; N-linked glycosylation is underlined; Putative catalytic triad is shaded; The conserved amino acids are boxed)
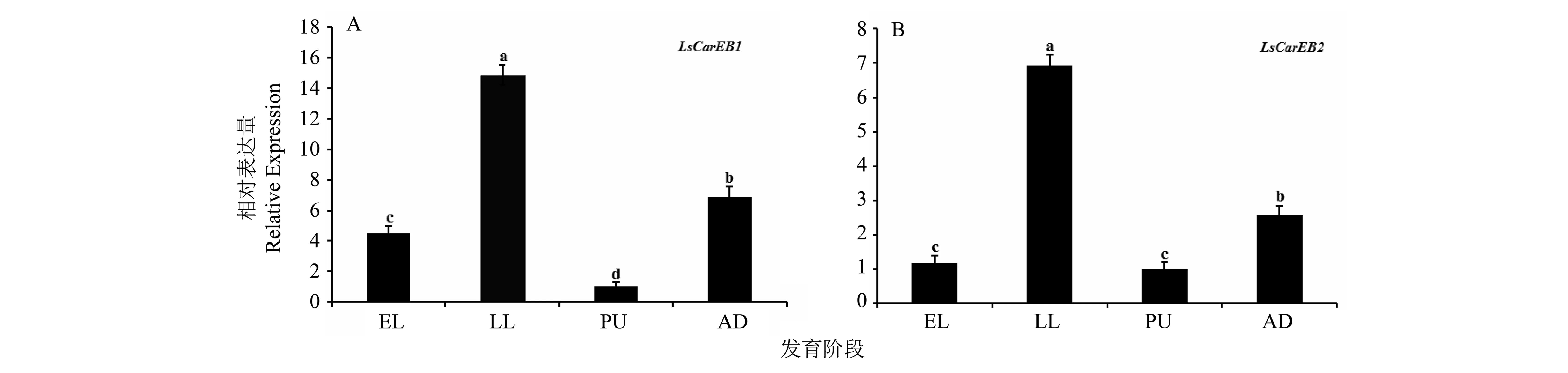
图4 烟草甲不同发育阶段LsCarEBs的相对表达量(柱状图上的不同小写字母表示在P < 0.05水平差异有统计学意义;EL、LL、PU、AD分别代表烟草甲的低龄幼虫、高龄幼虫、蛹和成虫A:LsCarEB1;B:LsCarEB2)Fig.4 Relative expression levels of LsCarEBs in different development stages of L. serricorne (Different lowercase letters above bars indicate statistically differences at the 0.05 probability level;EL: early larvae;LL: lately larvae;PU: pupae; AD: adults. A: LsCarEB1; B: LsCarEB2)

图5 不同φ(CO2)诱导下烟草甲高龄幼虫体内LsCarEBs的相对表达量(柱状图上的不同小写字母表示在P < 0.05水平差异有统计学意义;Control:对照;1:LC10(φ(CO2)=30% + φ(空气)=70%);2:LC30(φ(CO2)=70% + φ(空气)=30%);3:LC50(φ(CO2)=90% + φ(空气)=10%);A:LsCarEB1;B:LsCarEB2)Fig.5 Relative expression levels of LsCarEBs in lately larvae of L. serricorne after exposure to different volume fraction of CO2(Different lowercase letters above bars indicate statistically differences at the 0.05 probability level;Control; 1: LC10 (φ(CO2)=30% + φ(air)=70%); 2: LC30 (φ(CO2)=70%+ φ(air)30 %); 3: LC50 (φ(CO2)=90%+ φ(air)10%);A: LsCarEB1; B: LsCarEB2)
研究发现CarEs基因在昆虫不同发育阶段的表达存在一定的差异。如东亚飞蝗Locustamigratoria的LsCseF1在卵期和低龄若虫期表达较低,在4龄若虫表达量上调达到峰值,且一直持续到成虫期第5天[32]。而橘小实蝇BdCAREB1主要集中在3龄幼虫期高表达,进一步分析发现,幼虫发育至3龄幼虫进入暴食期,随着食物摄入的外源毒素显著增加,因此BdCAREB1的表达量相应升高[33]。本研究发现烟草甲LsCarEB1和LsCarEB2在不同发育阶段均有表达,高龄幼虫期的表达量显著高于低龄幼虫、蛹和成虫期,这可能与2个基因在高龄幼虫期发挥重要作用相关,其具体的功能解析还有待于进一步研究。
CO2气调技术是一种环保安全的新型害虫防治技术,它主要利用高φ(CO2)和低φ(O2)协同作用来提高对害虫的控制效果[5]。在同一昆虫中,不同发育阶段对CO2气调的耐受能力存在显著的差异,如粉螟C.cautella各发育时期的试虫经φ=99% CO2处理后,发现幼虫、蛹和成虫的致死时间分别为99.9、78.1和37.5 h,幼虫对CO2气调胁迫的耐受性最强;而赤拟谷盗经φ=99% CO2处理后,其幼虫的致死时间明显高于成虫,说明该虫的幼虫阶段具有更强的耐受性[7]。基于此,选择耐受性相对较差的发育阶段作为控制害虫的关键时期,将为有效地应用CO2气调防治害虫提供便利。本课题组前期研究发现,烟草甲高龄幼虫对CO2气调具有较强的耐受能力,进一步分析发现其体内羧酸酯酶的活性显著高于对照[18],基于羧酸酯酶在外源物质代谢方面的重要作用,推测高龄幼虫期CO2高耐受性可能与羧酸酯酶的活性升高密切相关。本研究发现,CO2气调处理烟草甲6 h后,LsCarEB1和LsCarEB2表达量均显著高于对照组,且mRNA的相对表达量随着胁迫φ(CO2)的升高而逐渐上调,我们推测这2个基因可能共同参与了烟草甲对CO2气调胁迫的应激响应。
本研究成功克隆烟草甲LsCarEB1和LsCarEB2的全长cDNA,据软件预测这2个基因编码的蛋白质均属于β酯酶;LsCarEB1和LsCarEB2在烟草甲各发育阶段均有表达,且在高龄幼虫期的表达量最高,均能被CO2气调胁迫诱导表达,说明它们是烟草甲生长发育的重要参与者,并在抵御CO2气调胁迫过程中起着关键作用。本研究为阐明CarEs基因在昆虫抵抗气调胁迫的分子机制奠定基础,同时为利用CO2气调防控害虫提供新的思路。
猜你喜欢
轻工学报(2022年3期)2022-06-22
食品科学(2022年6期)2022-03-30
农业技术与装备(2021年8期)2021-12-05
建材发展导向(2021年7期)2021-07-16
建材发展导向(2021年24期)2021-02-12
中国果菜(2020年9期)2020-12-16
建材发展导向(2019年5期)2019-09-09
世界农药(2019年2期)2019-07-13
分析化学(2017年12期)2017-12-25
船舶(2015年6期)2015-11-17
