
基于椭圆脐点突变模型的大熊猫生存状态研究
2018-08-09 08:31闫志刚李俊清
生态学报 2018年13期
闫志刚,李俊清,*,孙 立
1 北京林业大学林学院,北京 100083 2 中华人民共和国国务院参事室,北京 100006
得益于有力保护,大熊猫野生种群数量与栖息地面积近30年来一直处于增长态势。随着全国第四次大熊猫调查报告的发布,在2016版《世界自然保护联盟濒危物种红色名录》中,大熊猫受威胁等级由“濒危”降为“易危”[1]。但是值得关注的是,伴随着栖息地面积与野生种群数量的增长,栖息地破碎化与局域种群隔离问题也达到了历次调查之最;而且大熊猫的生存还受到诸如人为干扰、自然灾害等不利因素的影响[2]。生境的丧失与退化、人为干扰、自然灾害等因素能够相互强化,增加物种的灭绝风险[3];同时,IUCN在对物种受威胁等级的认定过程中,也未考虑物种的生物学习性,因此,IUCN提醒,大熊猫不再被列为濒危物种可能只是暂时的[1]。
生境的丧失与退化被视为导致物种濒危的首要原因[4]。濒危物种基于时空尺度,可以分为进化时间尺度中的濒危与生态事件尺度中的濒危[3]。大熊猫生存至今经历了漫长的历史进程,反映出这一物种具有顽强的生命力,其濒危原因更大程度上在于生态原因[5]。作为K对策生物,兼具高度特化,大熊猫更容易受到生境的丧失与退化的影响。因此,简单依据种群数量或栖息地面积单一指标的变化考察野生大熊猫生存状态是不严谨的。大熊猫自300万年前便形成了对以“森林+竹林”为特征的栖息地生态系统的生存依赖,其受威胁的主要原因在于生态系统的退化[6]。因此,从生态系统的视角出发,才能形成对大熊猫野生种群生存状态的正确认知。
生态系统由于非线性机制与反馈作用等原因,存在多稳态现象。生态系统发生稳态转换的点或区域称之为生态阀值,是生态系统本身能抗御外界干扰、恢复平衡状态的临界值[7]。生态系统的退化与生态系统的稳态转换有关。当生态系统临近阈值时,外界的微小干扰也会引发生态系统的剧烈响应,使之进入结构和功能截然不同的另一稳定状态,这种现象称为生态系统突变[8]。生态系统多稳态的存在是产生突变的主要原因[9]。生态系统突变属于分岔现象。按照动力学理论,在参数空间,当控制参数变化时,非线性系统的形态也将发生改变;形态的转变,意味着原有形态的稳定性发生了变化,即失稳。此时,系统会由一个定常状态(定态解)产生出多个定常状态,这就是所谓的“分岔”。分岔现象是一种重要的非线性现象,在自然界中有多种表现。系统失稳是发生分岔的前提,分岔之后,系统在不同状态间便发生了不连续的过渡,即“突变”。分岔和突变是对同一动力学现象从不同角度的解释。突变是控制参数变化到分岔点上出现的从一种定态向另一种定态的骤变[10]。作为系统科学中研究分岔与突变的数学理论,突变理论采用与动力学系统相关联的势函数来描述系统行为,其突出的优点是即使不了解系统的动力学方程,用几个状态变量就可以预测系统诸多的定性或定量性态[10- 11],因此其可以作为生态系统演化研究、状态评价的理论。
生态系统是由生物和环境构成的整体,两者之间相互影响、相互制约。生态系统是一个典型的复杂系统,复杂系统是指由大量相互作用的不同单元组成的自适应系统[12- 13]。复杂系统具有层次结构,层次即系统内部所具有的不同结构等级;系统内部处于同一层次的诸要素(子系统), 互相联结成一个整体,并表现出各要素都没有的新的性质[14]。复杂系统的这些特性在生态系统中的表现被称之为生态学的三大原理,即相互联系的整体性原理、生物组织层次理论以及涌现性原理[15]。系统科学是应对和解决复杂系统问题的有力工具,因此,系统科学的理论与方法适用于对生态系统的研究。基于系统科学,生态学的研究对象如个体、种群、群落、生态系统、景观等,都符合系统科学对系统的定义,均可视为系统科学意义上的“系统”[16]。对于生态系统而言,可以依据研究需要选择某一层次,并根据功能、结构或时空等,分解得到诸多相互依赖的子系统,子系统之间、子系统与外界环境之间进行着物质、能量、信息的交换,协同作用,共同进化,这种或强或弱的耦合作用,也是生态系统动力学现象的根源[17- 18]。
大熊猫分布区指大熊猫存在和居住的区域;大熊猫栖息地是指大熊猫具体生活的生境或小生境,大熊猫只出现在分布区内适宜生活的栖息地上;大熊猫潜在栖息地是指与栖息地相连、具有适宜于或比较适宜于大熊猫生息繁衍的生境条件、以往存在或可能存在或从未存在过大熊猫分布、在调查中没有发现大熊猫实体或活动痕迹的地域,通过加强管护,在10—15年间大熊猫很有可能扩散到该区域[19]。全国第三、四次大熊猫调查(以下简称3调、4调)分别于1999—2003年、2011—2014年实施,调查数据反映了近期大熊猫种群与栖息地等变化情况。对比3调与4调数据可知,在过去的10年间,大熊猫野生种群数量增加了268只(增幅16.8%),栖息地增加约2716 km2(增幅11.8%),潜在栖息地增加约541 km2(增幅6.3%);同时由于干扰的加剧,栖息地破碎化与局域种群隔离呈现同步增强的态势,栖息地斑块较多,部分斑块质量较差,局域种群隔离严重,小种群复壮困难[2,19- 21]。一方面是栖息地面积与野生种群数量的总体增长,一方面是栖息地破碎化与种群隔离愈加严重,形成鲜明对比。因此基于种群或栖息地单一指标,无法形成对当前大熊猫生存状况的认知;而且,单项指标的消长也只是反映了生态系统某一方面的变化,无法解释其整体的状况。
突变理论作为一种成熟的系统分析理论和方法,可以不囿于单项指标的变化,从整体层面较好解决这一矛盾现象。大熊猫受威胁的主要原因是栖息地的退化,保护栖息地是保护大熊猫的关键。森林与竹子构成的复合栖息地生态系统,与大熊猫野生种群构成一个自然的协同进化体[6,22]。因此,对于大熊猫生存状态的研究,可以依据突变理论,选择种群、栖息地、潜在栖息地作为子系统(即状态变量),构建“大熊猫—栖息地”系统突变模型进行分析,将栖息地与种群作为整体来考虑,对大熊猫生存状态进行了研究,并结合生态学理论,提出有价值的保护策略,这即是本文拟解决的科学问题与所采用的技术方法。在保护生物学与大熊猫研究中,具有一定的独创性;同时,该研究方法作为一种基于系统分析的方法论,具有普遍适用性。
1 数据来源与研究方法
1.1 数据来源
本文数据来源于2006年出版的《全国第三次大熊猫调查报告》、2014年发布的《全国第四次大熊猫调查报告》(未公开出版)、2015年出版的《四川的大熊猫:四川省第四次大熊猫调查报告》[19- 21],以栖息地和潜在栖息地的面积、大熊猫野生种群数量作为基础数据,时间跨度为1999—2014年,空间区域为大熊猫分布区(秦岭、岷山、邛崃山、大相岭、小相岭、凉山山系)。由于技术方面的限制,种群数据不包括1.5岁以下大熊猫幼体数量。
1.2 研究方法
主要研究方法与理论:相关性分析与突变理论。
1.2.1 相关性分析
对3、4调各山系大熊猫种群数量与栖息地、潜在栖息地面积进行复相关分析。
1.2.2 突变理论
突变理论是研究客观世界非连续性现象(突变)的理论,依据数学模型来描述和预测事物的连续性中断的质变过程。突变理论并不给出产生突变机制的假设,而是提供一个合理的数学模型来描述现实世界中产生的突变现象,对它进行分类,使之系统化,其数学基础是奇点理论和分岔理论。在突变理论中,系统在任何时刻的状态可以由给定的n个状态变量xi的值确定,同时系统还受到m个控制变量uj的控制,即uj的取值决定了xi的值,所要解决的问题是控制变量在何种情况下,系统势函数临界点类型会发生变化。
设系统势函数为V,则定义系统的平衡曲面M为:
xV=0
(1)
平衡曲面M是由系统Λ的全部平衡点构成,定义奇点集S是由Λ的全部退化临界点组成,其为M的一个子集,满足:
(2)
det表示行列式,H(V)表示V的Hessen矩阵。S在控制变量空间C中的投影,称为分岔点集B,分岔点集B是C中所有使V的形式发生变化的点的集合。
当状态变量数目不超过2、控制变量数目不超过4时,只存在7种基本突变类型,对应着具有不同数量的状态变量与控制变量的系统[23- 24],如表1所示。

表1 基本突变模型的势函数
根据本文的研究内容,需选用椭圆脐点突变模型,其势函数表达式为:
(3)
x,y为状态变量;w,u,v为控制变量。对于Vx,y求偏导数并令其为零,得平衡曲面M为:
(4)
奇点集S除满足上式,还需满足
Δ=4w2-x2-y2=0
(5)
系统发生稳态转化的基本判据为:
w<0
(6)
即当w<0时,势函数的极值分布会发生突变,系统呈现不稳定状态。
在失稳的情况下,系统的临界点(x0,y0)可以由下式求解:
(7)
式中,Φ0=-uw2-0.25v2,Φ1=2w2+2uw,Φ2=-3w2-u
1.2.3 突变模型的构建
大熊猫作为典型的K对策生物,种群密度波动较小,其种群平衡点处在环境的K值附近。因此,可以认为“大熊猫—栖息地”系统存在着较为稳定的耦合关系,表达式如下:
Z=fX,Y
(8)
式中,Z为大熊猫野生种群数量(只),X为栖息地面积(102km2),Y为潜在栖息地面积(102km2)。
栖息地破碎化对于种群存在多种不利影响[25]。为了避免灭绝旋涡的发生,一个种群需要保有一定数量的栖息地面积(Minimum Area Requirements)才能保证长期生存,对于大熊猫野生种群而言,所需最小栖息地面积约为114.7 km2[26- 27]。本文选用了100 km2作为与大熊猫野生种群相对应的栖息地面积指标单位,既便于处理数据,又不会从实质上改变种群与栖息地之间的耦合函数关系,从而影响到模型构建的精度以及计算结果的准确性;潜在栖息地亦然。
为了求解系统势函数,可以认为fX,Y与其Taylor展开式之间同构,并设f是3确定的,有:
Z=a30X3+a21X2Y+a12XY2+a03Y3+a20X2+a11XY+a02Y2+a10X+a01Y+a00
(9)
系数aij,可依据六大山系大熊猫野生种群数量与栖息地、潜在栖息地的面积数据,利用SPSS软件进行非线性拟合得到,进而得到如下形式(求解步骤见附录A):
Z=z+K00
(10)
式中,K00为常数,与系统状态的判别无关,z即为所需椭圆脐点突变模型表达式:
(11)
2 计算结果
2.1 相关性分析结果
大熊猫野生种群数量与栖息地、潜在栖息地的面积复相关分析结果见表2。

表2 大熊猫野生种群数量与栖息地、潜在栖息地的面积复相关分析结果
根据复相关分析可知,无论3调与4调,大熊猫野生种群数量与栖息地、潜在栖息地的面积均呈高度显著正相关,支撑了前文中的观点,即大熊猫野生种群与栖息地构成协同进化体,保护栖息地是保护大熊猫的关键。同时,也反映了大熊猫野生种群数量与其栖息地系统之间存在着较为稳定的耦合关系,可以此作为构建突变模型的基础。
2.2 突变模型分析结果
按照前文方法,依据全国大熊猫调查数据,按照式(9)构建大熊猫野生种群数量与栖息地、潜在栖息地的面积耦合关系函数,相关系数见表3。
按照附录A所示步骤,进一步转化为椭圆脐点突变模型,模型相关系数见表4。
由此可得系统突变模型的稳态判据,相关结果见表5。

表3 耦合关系函数的系数
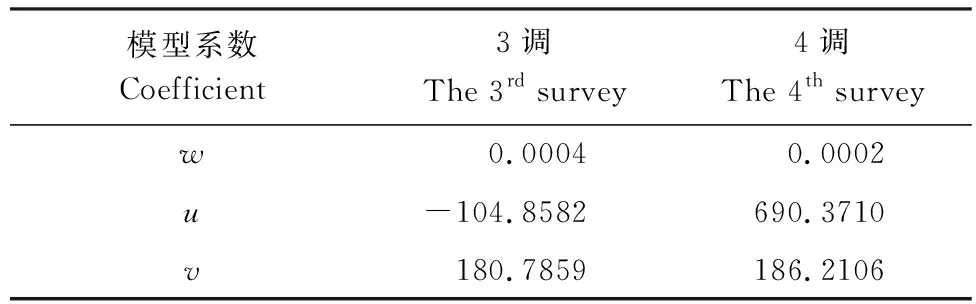
表4 椭圆脐点突变模型的系数

表5 系统突变模型的稳态判据
可见,4调期间,大熊猫野生种群数量与栖息地、潜在栖息地的面积虽然处于增长态势,但根据突变模型的分析结果,“大熊猫—栖息地”系统的稳定性却处于下降趋势,且均接近于临界状态,表明大熊猫赖以生存的生态系统具有较大的退化压力。
3 讨论
3.1 大熊猫生存状态的原因分析
由于突变理论不提供导致系统发生突变的机制,所以应用突变理论对生态系统稳定性与大熊猫生存状态的研究,实质是对事物发展“结果”的数学解读,是一种唯象理论(Phenomenology)的表现,因此需进一步结合生态学理论进行原因分析。总体上,保护与干扰的长期共存与博弈,构成了近期大熊猫保护的基本格局,分布区生态系统的稳定状态与大熊猫的生存状态可以视为保护与干扰相互作用的结果。分布区生态系统对大熊猫种群具有重要的影响,我国实施的一系列生态与物种保护工程,对于大熊猫种群的恢复发挥了重要作用;人为干扰作为历史上导致大熊猫种群衰退的重要原因,也是近期导致大熊猫栖息地破碎化与种群隔离的最重要影响因素,同时,趋于严重的栖息地破碎化与局域种群隔离又导致了大熊猫分布区生态系统稳定性的下降。
大熊猫栖息地生态保护力度的加大是野生种群持续增长的主要原因[19]。自1998年,大熊猫分布区内的天然林采伐已全面停止,并逐步实施了“天然林保护”、“退耕还林”、“长江防护林”等重大生态工程,这使得大熊猫生存所依赖的栖息地生态系统总体上得到了较好的保护与休养生息,大部分区域的植被及竹林得到了较好的自然恢复;同时,随着大规模森林采伐作业活动的停止,相应的人为干扰大幅减少,使得原本因采伐公路及运输车流而隔离的大熊猫种群,可以较顺畅的进行交流,降低了部分局域种群的灭绝风险,为种群数量增长创造了条件[28]。2001年启动的“全国野生动植物保护及自然保护区建设工程”,新建或扩建了相当数量的保护区,相对于3调,4调期间保护区面积增加了约11800 km2,增幅54.3%;尤其对于秦岭中部、岷山中部以及邛崃山中部的大熊猫核心种群,新保护区的建立及原有保护区保护管理能力的提升,使得这些大熊猫核心种群得到了更有效的保护,这些核心种群的数量占野生大熊猫总数量超过三分之二,对于大熊猫保护工作全局至关重要[2,19- 20]。
但是,在栖息地与种群持续恢复的同时,大熊猫保护仍面临重要的危机,主要表现为:经济社会发展带来长期且强烈的干扰,加剧了栖息地破碎化与种群的隔离,在一定程度上削弱了保护工作的成效,进而加重了大熊猫的生存危机[2]。
3.1.1 栖息地破碎化是导致生态系统稳定性下降的重要原因
目前,大熊猫栖息地总面积虽有所增加,但没有在宏观上改变大熊猫分布区整体狭长这一状况,并且大熊猫核心分布区栖息地整体破碎化程度有所加剧。新分布区与新种群的发现是4调栖息地面积与种群数量增长的主要因素,但对于大熊猫核心分布区的秦岭与岷山山系,4调栖息地面积仅增加了5.4%、1.2%,均未发现新的分布种群[20];3调时面积最大的两个栖息地斑块(岷山A栖息地、邛崃山系栖息地)在4调时已经破碎化,随之两个规模较大的局域种群也被隔离成几个较小的种群[19- 20];总体上,目前大熊猫栖息地被隔离成33个斑块,其中10个斑块的面积均小于114.7 km2,即30.3%的栖息地斑块不能满足种群的长期生存需要[20- 21,27],依据野生大熊猫最小存活种群数量为30只的标准[29- 31],33个局域种群中有22个存在灭绝风险,占比66.7%。
栖息地破碎化对物种的生存会构成严重的威胁[32]。在人类活动的影响下,分布区域狭窄、数量稀少的大型哺乳类物种会面临很高的灭绝风险[33];局域种群隔离以及种群内个体数量的减少,会引起瓶颈效应、近亲繁殖、遗传漂变等不良效应;栖息地破碎化会引起边缘效应、隔离效应、面积效应等不良效应,对物种之间的相互作用也会产生不良影响,包括种间连接的数量与强度[34];多种不良效应的综合作用与相互影响,会引发一个相互作用、相互强化的生物与非生物过程,导致种群趋于灭绝[35- 36]。对于大熊猫,还存在着另一个严重的威胁——竹子开花,当竹子开花发生在完全隔绝的小生境中,该区域大熊猫会面临严重的生存危机。
栖息地破碎化对生物多样性也具有重要的影响,栖息地丧失与破碎化被认为是造成生物多样性下降的最主要原因,其产生的危害也最大[33- 34]。Haddad等学者根据目前世界上规模最大和历史最悠久的破碎化实验发现,栖息地破碎化会导致生物多样性的大幅降低与生态系统功能的丧失,该项实验地跨五大洲、时间跨度35年,涵盖了不同的生物群落与生态系统[37]。生物多样性的丧失对生态系统具有重要的影响。尽管存在争议,但大多数关于生态系统结构与功能的理论探讨和实验生态学研究成果表明,生物多样性与生态系统的稳定性呈正相关,栖息地破碎化会导致生态系统稳定性的降低[32- 33]。在另一项基于大熊猫局域种群与栖息地斑块的模型研究中,我们也发现,如果考虑栖息地破碎化的影响,虽然4调时大熊猫栖息地总面积与种群总数量相对于3调时有所增长,但趋于严重的栖息地破碎化与种群隔离导致了4调时分布区生态系统的稳定性,相对于3调时反而呈现更不稳定的现象[38]。
3.1.2 干扰对大熊猫栖息地的影响
人类的经济社会活动以及近年来频发的自然灾害,对大熊猫栖息地造成了较严重的干扰,加剧了栖息地的退化与破碎化。干扰因素基本上可以分为自然灾害和人为干扰两类,对栖息地的影响与作用形式有所区别。
大熊猫分布区位于青藏高原东缘的构造带,存在多条断裂地震带,近期发生的一系列地震对栖息地的影响显著。以2008年发生的汶川地震为例,依据3调统计数据进行灾害评估,地震造成大熊猫栖息地毁损面积约578 km2,占全国栖息地总面积的2.5%,造成潜在栖息地毁损面积约20 km2,占比0.2%,主要集中在岷山与邛崃山等大熊猫核心分布区[21]。地震及其次生灾害显著改变了大熊猫栖息地的地形、地貌、土壤特性及植被演替进程,栖息地结构和质量发生显著变化;震区大面积区域的增强型植被指数(EVI)显著下降,加剧了大熊猫栖息地的退化和破碎化进程,经过5年的恢复,4调时(2013年)发现多数区域EVI虽有明显改善,但尚未恢复到2007年的水平,大部分区域大熊猫栖息地植被仍在恢复中[21,39],后继发生的雅安、九寨沟地震,对大熊猫栖息地又构成了一定的影响。
同时,近年来,经济和社会的快速发展延伸到了大熊猫分布区,其深度与广度为历史最强,造成了强烈的冲击。公路、铁路及低等级的县域、乡村道路的不断增加,致使路网密度进一步增大;畜牧业、水电站建设与矿产开发呈现增加态势;作为山区经济增长点的旅游开发带动了更多旅游基础设施建设与游客数量的增长[20- 21]。这些以经济发展为主导的宏观社会变化,给大熊猫分布区带来了长久的人为干扰。人为干扰强度与经济社会发展水平有关,这决定了人为干扰在近年来趋于加剧;同时,值得注意的是,人为干扰作为普遍存在并且长期持续的影响因素,不会因经济社会的发达或落后而消失,但其多发类型与地区经济发展水平有关。在经济较为发达的中北部地区,主要以水电开发、基础建设、旅游活动等、由政府主导的经济建设活动所带来的大型干扰为主;在欠发达的南部地区(凉山山系),主要以采薪、放牧、割竹打笋等、由个人经济生活行为所带来的一般干扰为主[20,40]。
结合4调可以发现,在大熊猫中北部分布区,一方面,诸如水电开发等大型干扰呈增强态势,水电站在邛崃山分布众多,采矿、旅游及其相关基础设施建设对岷山、邛崃山、秦岭的栖息地影响突出;另一方面,诸如放牧等一般干扰的发生数量与强度在这些区域呈明显下降[20]。大型干扰会对大熊猫及其栖息地造成持久性或永久性的影响,加剧了栖息地破碎化与种群的隔离[21]。一般干扰在南部分布区却有着强烈的影响,放牧、采笋、采薪等活动对大相岭、凉山山系影响明显[20- 21]。
总体上,在大熊猫栖息地内,干扰呈现集中分布,并具有密度大、强度高、时间长的特点,对生态系统构成了巨大压力,导致了栖息地的质量下降与破碎化,最终影响了大熊猫的分布格局;同时,人为干扰多发,但增强类型有所不同,与当地经济发展水平有关,应注意因地制宜,制定差异化的保护政策。
3.2 当前大熊猫保护工作的具体对策
应对栖息地破碎化与种群隔离的威胁是当前大熊猫保护工作的首要内容,应基于两个方面开展工作,一是通过科学合理的规划与实施野生动物走廊建设,以应对栖息地破碎化的直接威胁,二是根据各山系干扰的多发类型,采取具有地区差异化的保护政策,有针对性的采取减缓人为干扰的不利影响,从根源上遏制栖息地退化与破碎化进程。
3.2.1 针对栖息地破碎化的对策分析
栖息地破碎化对物种生存的影响首先体现在种群水平上,种群的消失速度比一个物种的灭绝速度要快得多[33],栖息地斑块面积的大小对生物多样性、物种灭绝阈值等也具有重要的影响[34],因此,应对栖息地破碎化危害的首要任务在于对隔离小种群的关注。同时,生物一般也具有一定的灭绝延迟能力,即使栖息地发生了严重破碎化,该区域物种的灭亡在时间上往往存在着几十年甚至几个世纪的延迟,即存在灭绝债务现象[33],这无疑为物种拯救行为提供了宝贵的时间窗口。
栖息地斑块的总量、质量、连接程度以及干扰的影响,对于破碎化的栖息地维持生物多样性和生态系统服务的能力具有决定性作用[37]。根据灭绝旋涡理论,在生境破坏初期,剩余适宜生境仍有较好的连通性,种群主要受生境损失的影响;随着生境破坏的日趋严重,斑块之间的连接将减弱,会引发物种对栖息地丧失的非线性反应,导致种群密度急剧下降;同时,一系列的反馈作用会增加生态系统的不稳定性和区域生态系统的随机性[41]。如果生态系统继续退化,一旦发生系统突变,未来的生态恢复将会非常困难。有研究表明,种群扩散率对于大熊猫小种群的复壮非常重要,即使扩散率低至1%也能有效降低局域种群的灭绝风险[42]。因此,基于大熊猫栖息地斑块分布的空间格局,采取诸如野生动物走廊等栖息地斑块的连接措施具有重要意义。以野生动物走廊作为促进小种群复壮措施,其实施效果主要取决于所连接种群的绝对规模。在小种群之间进行连接,对于小种群的增长与持久意义有限[42]。野生动物走廊的早期规划与建设应首先以面积较大的生境斑块与规模较大的种群为中心,向周边的小斑块与小种群进行辐射,这将形成一个“星型”生态网络;长期规划可设计和建造更多的走廊以连接更多的栖息地,将早期的星型生态网络转化为复杂生态网络。根据复杂网络理论可知,这种复杂生态网络具有更好的鲁棒性(Robust)和抗干扰能力,对于大熊猫野生种群而言,加强基因交流,也将有效地扩大基因库,可以更好抵御各种随机变化的影响[43- 44]。
3.2.2 针对干扰因素的对策分析
对于近期频发的自然灾害干扰——地震的影响,灾区大熊猫栖息地的恢复目标应以实现森林生态系统功能的修复为主。依据森林演替规律与自然进程,利用天然植被的自然恢复或人工重建的方式,通过对受损天然林的保育措施,实现森林面积与郁闭度的恢复,从而实现森林与竹林的协调更新;人工修复措施不应作为森林植被主要的修复手段,但对于受地质灾害影响严重的区域,应尽快引种乡土树种,以混交林形成保护[39]。
对于人为干扰的影响,应结合各山系生态系统特点、经济社会发展水平与多发干扰类型采取有针对性的措施。
秦岭、岷山、邛崃山山系作为大熊猫核心分布区,具有生态系统容量大、保护工作开展较好的特点,大熊猫种群呈稳定、缓慢的增长态势,反映了该区域大熊猫及生态保护工作趋于成熟。同时,这些地区的干扰呈现出大型干扰增强与一般干扰下降的发展趋势。在保护工作中应积极利用大熊猫交流廊道建设、扩建保护区等措施,注重对现有栖息地与潜在栖息地质量的提升。这一地区也是大熊猫国家公园主要所在地,应切实落实保护政策,稳步提升生态系统的规模与质量。大、小相岭山系的栖息地与大熊猫种群一直保持较好的恢复态势,同时又存在生态系统规模较小、人为干扰严重等不利因素,大熊猫种群灭绝风险较高。4调时,两山系栖息地增幅分别达到51.6%、48.8%,说明山系保护工作仍有较大潜力与改善空间,因此,一方面应继续加强保护,争取有效降低人为干扰;另一方面,考虑到种群规模较小,在实现小种群互通与栖息地斑块连接的基础上,应结合大熊猫放归,促进遗传交流,提高基因多样性,对于大熊猫的生存与延续尤为重要。凉山山系的大熊猫保护工作最为薄弱,是3调时唯一出现种群数量负增长的山系(增幅为-25.8%),4调时有所改善(增幅为7.8%),与其呈爪形的狭长地理格局以及经济发展落后有密切关系。狭长而分散的地理格局,造成种群隔离程度较高且局域种群规模过小,落后的地区经济,使得放牧、割竹打笋等与个人经济生活相关的人为干扰,一直非常严重。经济发展与生态保护并重,是当前凉山地区保护工作首要内容,实现生态保护、扶贫开发与地方经济发展的有机结合,改进产业结构,推进可替代生计培训等措施,减少经济对农牧业的依赖,并落实社区居民利益共享与补偿机制,以必要的经济手段提高共同参与保护的积极性,实现人与自然和谐共存。。
总体而言,经济社会的快速发展已深入大熊猫分布地区,其深度与广度为历史最强,由此引发的人为干扰也是近三十年来最为强烈,由此加重了大熊猫分布区栖息地破碎化和种群隔离,对生态系统造成了严重影响。人类干扰活动对大熊猫生境的影响具有协同增大效应,应结合各山系特点,确定对其他人为干扰具有放大作用的主导人为干扰类型,采取有针对性的措施,以期最优成效。
3.3 突变理论在生态学研究中的应用价值
3.3.1 突变模型在大熊猫研究中的应用
自然界虽然具有高度的复杂性,但很多时候,通过少量相关变量(关键因子)信息就足以构建一个有效的模型[15]。生态学模型构建的思想是基于事物发展与其主要矛盾之间的辩证关系。一个生态学模型应该包括三个要素:①空间因素,即系统边界的划分;②在所有功能单元中最重要的子系统(组分);③时间间隔[15]。因此,对于一个现实的生态系统,其模型构建的关键在于,根据研究问题确定相应的研究尺度,由此选取最重要的子系统(组分)作为关键因子(模型的变量),进而实现模型的构建。
突变理论的优势在于,即使不了解所研究系统的动力学方程,用几个状态变量即可预测系统诸多的定性或定量性态,并提供了数学模型来描述现实世界中产生的突变现象。对于生态学突变模型的构建,关键因子的选取十分重要。由表1可知,变量数目一定的突变模型,其表达式的形式是确定的,所以所选取的关键因子对于模型所蕴含的信息具有重要的影响作用。对于大熊猫种群,栖息地的重要性毋庸置疑,而潜在栖息地对于栖息地的增长、栖息地的联通等具有重要作用[45],因此,对于大熊猫的生存、种群复壮与扩散具有重要意义。所以本文在进行突变模型构建时,将潜在栖息地作为一项关键因子加以考虑,以保证模型蕴含信息的全面性、分析结果的准确性;同时,3调与4调在调查技术方法上具有一致性,也保证了数据来源的准确性,以及计算结果的可比性。
突变理论也有其局限性,即不能给出所研究问题产生突变的机制,也不能给出系统的动力学方程,因此,我们无法直观获悉模型参量与计算结果之间直接而明确的逻辑关系。应用突变理论进行模型构建以及对计算结果的解读,需要结合所应用领域的相关理论加以实现。
3.3.2 突变理论在生态学领域具有广泛的应用价值
2013年,英国生态学会在其成立100周年纪念之际,发布了100个极具研究价值的生态学问题,认为关于“生态系统的评估”以及“生态系统临界状态的判别”相关研究具有重要现实意义,同时还强调了将“其他学科已运用的理论运用于生态学”以及“发展和采用模型系统来理解生态系统”的重要性[46]。
当前地球生态系统面临前所未有的压力,科学评估生态系统的状态,并对生态系统重大突变进行预测与预警,对于地球系统与人类社会的可持续发展至关重要。但由于生态系统对外界的响应具有滞后性,同时,长时间、高分辨率数据的缺乏以及对现实生态系统内在复杂性的认识不足,使得生态系统的突变预测成为一项艰难的任务[8,47]。
突变理论作为一种成熟的系统科学理论,用形象的数学模型描述了系统连续性行为突然中断与质变发生的过程。突变理论所提供的理论与模型是对系统的描述,而非对机制的解释。如果所研究系统的机制未知或者过于复杂,根据奥卡姆剃刀原则(Occam′s Razor),应先假定系统具有与观察相吻合的最简单的机制,突变理论即提供了系统“最简单机制”的数学描述与模型,即以“降低描述的任意性”实现将“种种不同的观察统一于一个系统”[48]。突变理论研究的是系统稳定态跃迁的现象和规律。它认为系统存在稳定态和非稳定态,稳定态是指系统在微小扰动作用下,仍然能够保持原来状态;而一旦受到微小扰动就迅速离开原来状态,则是非稳定态;稳定态之间的转化,是以突变形式发生的;非线性是系统存在突变现象的重要原因。突变理论作为研究系统有序演化的有力数学工具,在生态学研究方面有着广阔的应用前景。
4 结论
(1) 大熊猫野生种群的数量与栖息地、潜在栖息地的面积呈高度显著正相关,且存在着较为稳定的耦合关系。大熊猫对栖息地生态系统具有高度的生存依赖,基于生态系统的视角,才能正确认知大熊猫野生种群的生存状态。保护栖息地是保护大熊猫的关键,大熊猫作为高度特化的K对策生物,更容易受到栖息地破碎化与种群隔离的影响。
(2) 采用生态系统的关键指标构建突变模型,可以从系统层面对大熊猫生存状态进行研究。近年来,大熊猫栖息地与野生种群总体增长的同时,栖息地破碎化和局域种群隔离也有所加剧。基于椭圆脐点突变模型分析可知,“大熊猫—栖息地”系统的稳定性处于下降趋势,且接近于临界状态,表明生态系统的仍具有较大的退化危险,大熊猫的生存危机依然严峻。
猜你喜欢
今日农业(2022年15期)2022-09-20
小哥白尼(野生动物)(2021年7期)2021-11-20
娃娃乐园·综合智能(2019年10期)2019-12-07
汉语世界(The World of Chinese)(2018年3期)2018-10-22
红土地(2018年7期)2018-09-26
学苑创造·A版(2018年7期)2018-08-07
厦门航空(2018年4期)2018-04-25
天天爱科学(2017年12期)2017-08-15
当代畜禽养殖业(2014年10期)2014-02-27
西南学林(2011年0期)2011-11-12
