
拉萨河谷草地群落的数量分类与排序
2018-08-09 08:28姚帅臣王景升丁陆彬包小婷刘文婧李妍妍
生态学报 2018年13期
姚帅臣,王景升,丁陆彬,包小婷,李 超,王 彤,刘文婧,李妍妍
1 中国人民大学环境学院,北京 100872 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101 3 山东建筑大学测绘地理信息学院,济南 250101
青藏高原是我国乃至亚洲的江河源,向来有世界屋脊和第三极之称,是我国及东亚的重要生态安全屏障区。草地生态系统是青藏高原发挥其生态安全屏障作用不可或缺的组成部分,承载着区域牧业经济发展的重担[1]。“一江两河”(是指雅鲁藏布江、拉萨河、年楚河)地区是西藏南部和喜马拉雅山脉中段生态安全屏障区的一部分,是区域经济重点发展亚区,也是雅鲁藏布江中游地区河谷土地沙化以及水土流失的控制区[2]。近年来,受气候变暖、过度放牧和樵采等人类活动的多重干扰[3- 4],河谷地区天然草地退化形势严峻,致使水土流失日益加剧,生态环境破坏严重,从而破坏了生态和经济的良性循环[5- 7]。
青藏高原由于其独特的地理位置和自然条件吸引了很多学者的目光。但是,目前对“一江两河”地区草地群落的研究多集中于多样性、植被营养元素、生物量动态以及对干扰的响应等方面[8- 13]。对河谷草地群落的内部分类、空间分布和生态因子变化特征以及它们之间的相关性的系统研究较少。仅有李海涛等[14]、杨小林等[15]、拉琼等[16]、张燕杰等[17]分别在西藏拉孜县、拉萨半干旱河谷地区、雅鲁藏布江河岸以及拉萨河谷进行的分类研究。但由于拉萨河谷地理位置特殊、地形复杂,由东到西、由南到北气候差异比较明显,植被类型的分布也比较复杂[18]。同时,学者们的研究区域、环境和群落结构差异较大,因此对群落自身的生态特点、内在规律的认识也各有偏重。数量分类和排序分析方法能够系统研究草地群落的内部分类、群丛特征,并能科学地揭示植被-环境间的生态关系[19- 23]。因此,我们需要加强对拉萨河谷草地群落的数量分类和排序研究,使我们对相关区域植物群落的分布格局以及植物与环境之间生态关系的认识更清晰、全面。
植被数量分类主要应用数学的方法完成分类过程,以揭示植物之间以及植物与环境之间的生态关系,反映出一定的生态规律。双向指示种分析 (Two-Way Indicators Species Analysis, TWINSPAN)是一种多元等级分划方法,其每次划分过程都会应用指示物种进行修正,不仅给出样方的分类结果,也会给出物种的分类结果。也就是说,它能同时对样方和物种进行分类,并且反映出样方和物种与环境的关系。因此,TWINSPAN是现今最为成熟,应用最为广泛,使用最多的数量分类方法,国内外学者将其应用于各种群落类型的数量分类[24- 33]。排序是研究植被的连续变化的分析方法,它是现代植被分析的重要手段,通过将样方或植物物种排列在一定的空间,从而使排序轴可以反映出相关的生态梯度,进而解释植被分布以及植物物种的分布格局和它们与环境之间的相互关系。去趋势对应分析(Detrended Correspondence Analysis, DCA)是消除了对应分析中易于出现的弓形效应的排序方法,它在计算的过程中只需要样方和物种组成的数据矩阵。典范对应分析(Canonical Correspondence Analysis, CCA)是一种基于单峰模型的排序方法,样方排序与对象排序对应分析,而且在排序过程中结合多个环境因子,因此可以把样方、对象与环境因子的排序结果表示在同一排序图上[19]。在实际的植被数量研究中,研究一般将数量分类和排序结合起来,以更好的了解植被之间,以及植被和环境之间 的生态关系。在目前的研究中学者们使用到了不同的分类和排序方法的组合进行植被的数量研究。学者们的研究表明,CCA排序轴的生态意义易于解释,并且能同时反映出各样方在物种组成和环境因子组成上的趋同性;DCA排序图中样方较为分散,群落间的界线比较清晰,与分类结合使用的效果要好[34- 37]。也就是说,将TWINSPAN分类和DCA、CCA排序的结合应用是非常科学合理的。
本文应用双向指示种分类法、典范对应分析和去趋势对应分析等方法,定量分析了环境因子对高原河谷草地植物群落分布和物种组成的影响。以期阐明高原河谷草地的地理分布格局,为合理保护利用当地草地资源提供参考,为西藏河谷区畜牧业可持续发展以及生态安全屏障建设提供科学依据。
1 研究区概况
研究区地处念青唐古拉山脉中段南翼,位于拉萨河上游澎波河流域,属于西藏自治区拉萨市林周县卡孜乡。该区域属高原温带季风半干旱气候区,平均海拔4500 m左右,河谷内气候温暖、干燥,年均温7.5℃,日温差大,最热月平均气温18.7℃,最冷月平均气温-1.7℃,多年极端最高气温29.6℃,极端最低气温为-16.5℃,分别出现在6月和1月;年均降水量440 mm左右,且降水主要集中在6—9月,无霜期110 d左右,气候干燥[38]。研究样地位于卡孜乡白朗村沟域的阴坡,天然草地类型主要包括亚高山草甸草原、亚高山灌丛草甸和高山草甸三大类,区内没有乔木树种的分布,灌丛高度一般不超过50 cm,盖度较小,草本层高度一般不超过20 cm,盖度较大。其中,亚高山草甸草原土壤类型为亚高山草甸草原土,草本优势种为垂穗披碱草(Elymusnutans)、草沙蚕(Tripogonbromoides)、青藏薹草(Carexmoorcroftii)和高山嵩草(Kobresiapygmaea)等,群落总盖度39%—50%;亚高山灌丛草甸土壤类型为亚高山灌丛草甸土,灌木层优势种为金露梅(Potentillafruticosa)、微毛樱草杜鹃(Rhododendronprimulaeflorum)、雪层杜鹃(Rhododendronnivale),草本优势种为高山嵩草,群落总盖度52%—90%;高山草甸土壤类型为高山草甸土,草本优势种为高山嵩草等,群落总盖度71%—87%。近年来由于畜牧业的快速发展,牲畜的啃食、践踏强度增加,加之人类活动对天然植被的干扰,原有植被发生一定程度的退化[10]。
2 研究方法
2.1 群落调查
研究采用样带和海拔梯度相结合的方法,以海拔4000 m为起点,每间隔海拔50 m左右布设一条样带,样带方向垂直于样地海拔等高线.在样带上随机选取5个1 m ×1 m的样方,至海拔5100 m处,共计样点23个,样方115个。记录每个样方中出现的植物种类,未能鉴定的植物采集标本并且拍照,记录其所在样方编号,带回实验室进一步鉴定。用卷尺测量每个物种的垂直高度、群落平均高,网格法测定物种盖度、总盖度等参数,同时使用GPS、罗盘仪测量记录海拔、坡度、坡向等生境因子。在每个样方中央和对角用环刀采取土壤样品,在实验室采用烘干法测定土壤含水量、土壤容重等数据。
2.2 数据处理
计算样方内各草本植物的重要值,其计算公式如下[39]:
草本植物重要值=(相对盖度 + 相对高度)×100/2
野外调查于2015年8月生物量高峰期完成,使用调查数据建立两个矩阵:由23个样点115个样方中的54种草本的重要值组成的植被矩阵;由23个样点和5个环境参数组成的环境属性矩阵。应用Wintwins 2.3中TWINSPAN进行植被数量分类,应用Canoco for Windows 4.5中的CCA和DCA进行样方和环境参数的排序。
3 结果与分析
3.1 样点的TWINSPAN数量分类
将23个样点进行TWINSPAN分类并依据植物群落分类和命名原则,结合调查结果的生态分析和群落生境特征的指示物种或优势物种命名群落类型,本区植物群落可分为8类,如图1,各群落主要特征见表1。

图1 拉萨河谷样点TWINSPAN分类树状图Fig.1 Dendrogram of the TWINSPAN classification of 23 plots in Lhasa River Valley指示物种:1.半球齿缘草; 2.矮火绒草; 8.独一味; 11.二裂委陵菜; 26.麦薲草
垂穗披碱草+草沙蚕群落,包括样点1,共1个样点。该群落分布于海拔3993 m左右的阴坡,坡度7°,土壤含水量42%,土壤容重1.17 g/cm3。群落总盖度39%,优势种为垂穗披碱草(Elymusnutans)和草沙蚕(Tripogonbromoides),主要伴生种有半球齿缘草(Eritrichiumhemisphaericum)、二裂委陵菜(Potentillabifurca)、高山嵩草、拉萨厚棱芹(Pachypleurumlhasanum)、木根香青(Anaphalisxylorhiza)、弱小火绒草(Leontopodiumpusillum)、丝颖针茅(Stipacapillacea)、酸模叶橐吾(Ligularialapathifolia)、狭囊薹草(Carexcruenta)、线叶嵩草(Kobresiacapillifolia)、云南棘豆(Oxytropisyunnanensis)等。
青藏薹草群落,包括样点2、3、4,共3个样点。该群落分布于海拔4053—4157 m的阴坡,坡度9°—13°,土壤含水量27%—47%,土壤容重1.17—1.36 g/cm3。群落总盖度56%,优势种为青藏薹草,主要伴生种有半球齿缘草、多裂委陵菜(Potentillamultifida)、二裂委陵菜、黄苞天南星(Arisaemaflavum)、青藏狗娃花(Heteropappusbowerii)、肉果草、丝颖针茅、酸模叶橐吾、狭囊薹草等。
高山嵩草+青藏薹草群落,包括样点5、6、7、8,共4个样点。该群落分布于海拔4200—4346 m的半阴坡,坡度16°—20°,土壤含水量66%—76%,土壤容重0.81—0.98 g/cm3。群落总盖度45%—50%,优势种为高山嵩草和青藏薹草,主要伴生种有钉住委陵菜、二裂委陵菜、高山豆(Tibetiahimalaica)、肉果草(Lanceatibetica)、丝颖针茅、酸模叶橐吾、无茎黄鹌菜(Youngiasimulatrix)等。
金露梅+高山嵩草群落,包括样点9、10、11,共3个样点。该群落分布于海拔4393—4480 m的阴坡,坡度18—22°,土壤含水量87%—120%,土壤容重0.56—0.75 g/cm3。群落总盖度84%—90%,灌木层盖度6%,灌木层优势种为金露梅(Potentillafruticosa),草本层盖度78%—84%,草本优势种为高山嵩草,主要伴生种有矮火绒草(Leontopodiumnanum)、大花嵩草(Kobresiamacrantha)、钉柱委陵菜(Potentillasaundersiana)、独一味(Lamiophlomisrotata)、高山唐松草(Thalictrumalpinum)、禾叶点地梅(Androsacegraminifolia)、麦薲草(Elymustangutorum)、青藏薹草、丝颖针茅、微药羊茅(Festucanitidula)、狭囊薹草、羊茅(Festucaovina)、珠芽蓼(Polygonumviviparum)等。
微毛樱草杜鹃+高山嵩草群落,包括样点12、13、14,共3个样点。该群落分布于海拔4540—4650 m的阴坡,坡度22—28°,土壤含水量71%—95%,土壤容重0.69—0.82 g/cm3。群落总盖度66%,灌木层盖度11%,灌木层优势种为微毛樱草杜鹃,伴生种有金露梅和雪层杜鹃(Rhododendronnivale),草本层盖度55%,草本优势种为高山嵩草,主要伴生种有矮火绒草、垂穗披碱草、钉柱委陵菜、独一味、高山唐松草、禾叶点地梅、黑被薹草(Carexatrata)、横断山风毛菊(Pedicularissiphonantha)、冷地早熟禾(Poacrymophila)、木根香青、丝颖针茅、微药羊茅、狭囊薹草、线叶龙胆(Gentianafarreri)、羊茅、圆穗蓼(Polygonummacrophyllum)、云南棘豆、长梗秦艽(Gentianawaltonii)等。
微毛樱草杜鹃+雪层杜鹃+高山嵩草群落,包括样点15、16、17,共3个样点。该群落分布于海拔4700—4805 m的阴坡,坡度30—35°,土壤含水量95%—102%,土壤容重0.64—0.69 g/cm3。群落总盖度67%—75%,灌木层盖度22%—29%,灌木层优势种为微毛樱草杜鹃和雪层杜鹃,草本层盖度45%—56%,草本优势种为高山嵩草,主要伴生种有矮火绒草、钉柱委陵菜、冷地早熟禾、全萼秦艽(Gentianalhassica)、喜马拉雅嵩草(Kobresiaroyleana)、狭囊薹草、线叶嵩草、线叶龙胆、圆穗蓼等。
雪层杜鹃+高山嵩草群落,包括样点18、19、20,共3个样点。该群落分布于海拔4861—4954 m的半阴坡,坡度25°—34°,土壤含水量123%—155%,土壤容重0.47—0.57 g/cm3。群落总盖度52%,灌木层盖度4%,灌木层优势种为雪层杜鹃,伴生种有微毛樱草杜鹃,草本层盖度48%,草本优势种为高山嵩草,主要伴生种有矮火绒草、倒披针叶风毛菊(Saussureanimborum)、钉柱委陵菜、高山大戟(Euphorbiastracheyi)、高山唐松草、禾叶风毛菊(Saussureagraminea)、横断山风毛菊、麦薲草、扭盔马先蒿(Pedicularisdavidii)、全萼秦艽、西藏三毛草(Trisetumtibeticum)、喜马拉雅嵩草、线叶龙胆、线叶嵩草、颖毛早熟禾(Poahirtiglumis)、珠芽蓼等。
高山嵩草群落,包括样点21、22、23,共3个样点。该群落分布于海拔4993—5104 m的阴坡,坡度7°—18°,土壤含水量96%—167%,土壤容重0.44—0.66 g/cm3。群落总盖度71%—87%,优势种为高山嵩草,主要伴生种有钉柱委陵菜、高山唐松草、麦薲草、扭盔马先蒿、全萼秦艽、西藏三毛草、线叶龙胆、线叶嵩草、伊凡苔草(Carexivanovae)、缘毛紫菀(Astersouliei)、圆穗蓼等。
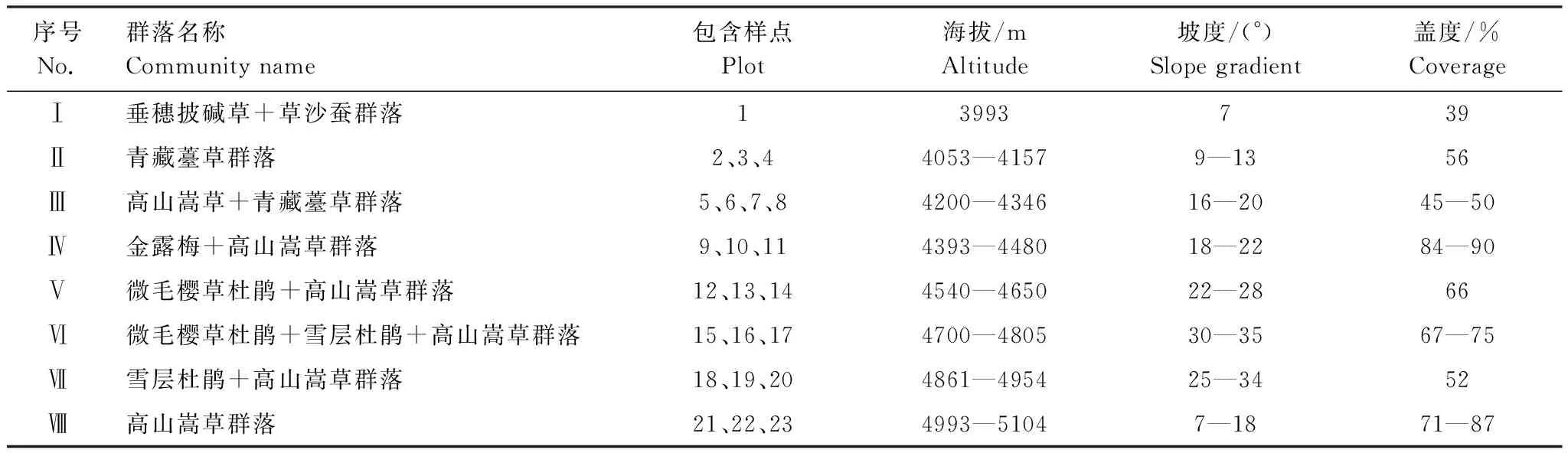
表1 拉萨河谷草地群落分类主要特征描述
3.2 样点DCA排序分析
采用DCA对研究区内的23个样点进行排序,结果如图2,前4个排序轴的特征值分别是 0.575、0.315、0.107、0.029。群落类型在DCA二维排序图上的分布一定程度上反映了群落空间变化的梯度和趋势,DCA排序的第一二轴特征值较大,说明这两个排序轴包含的生态信息量多,能准确的反映出各植物群落之间以及植物群落与环境因子之间的相互关系,具有重要的生态意义。从排序图可以看出,TWINSPAN的分类结果所指示的群落类型在二维排序图上基本都有自己一定的分布范围和界限,这说明DCA排序很好的反应了样方之间以及样方和环境之间的相对关系。从排序轴看,第一轴基本反映了海拔高度的变化梯度,从左到右,群落所在的海拔逐渐升高,最左边是分布在低海拔地区的垂穗披碱草+草沙蚕群落(样点1)、青藏薹草群落(样点2、3、4),最右边则是分布在高海拔的群落高山嵩草群落(样点21、22、23)。第二轴反映了坡向的变化,即第二轴从下往上,植物群落的坡向由阴坡向半阴坡的变化。可以看出,样点的DCA二维排序图反映的植物群落结构和生境的空间梯度,与TWINSPAN的分类结果相匹配。

图2 拉萨河谷样点DCA二维排序图Fig.2 Two-dimensional DCA ordination diagram of plots in Lhasa River Valley
3.3 样点CCA排序
为了进一步研究群落分布和环境因子之间的相互关系,我们对23个样点和5个环境因子进行典范对应分析CCA排序,所得结果如图3。沿CCA排序图第一轴,随着海拔的升高,群落从分布在较低海拔地区的垂穗披碱草+草沙蚕群落和青藏薹草群落过渡到分布在中间海拔的金露梅+高山嵩草群落、微毛樱草杜鹃+高山嵩草群落、微毛樱草杜鹃+雪层杜鹃+高山嵩草群落和雪层杜鹃+高山嵩草群落,最后是分布在高海拔地区的高山嵩草群落。排序轴的第一轴体现了海拔和土壤容重对植物群落分布的影响。沿排序轴第二轴从上向下,随着坡向的变化,群落的分布也表现出一定的差异。从排序结果可以看到,影响群落分布的主要环境因子是海拔,其次是坡向。也就是说,首先,海拔的差异决定着研究区内的植被被相应的分为3种类型:较低海拔的亚高山草甸草原、中海拔的亚高山灌丛草甸和高海拔的高山草甸,其次,在同一植被类型下,由于其他条件的不同,3种植被类型则又被划分为不同的群落。同时,在排序图中可以看出TWINSPAN划分的8类群落在CCA排序中也基本都有各自相应的分布范围和界限,这也在一定程度上证明了TWINSPAN的分类结果的准确性以及和样方CCA排序的一致性。

图3 拉萨河谷样点CCA排序图Fig.3 CCA ordination diagram of plots in Lhasa River ValleyAlt: 海拔, Altitude;Asp: 坡向, Aspect;Slo: 坡度, Slope;Sbd: 土壤容重, Soil bulk density;Sw: 土壤含水量, Soil water
3.4 物种的CCA排序
对54个物种进行CCA排序分析,结果如图4。可以看出,物种在排序图中的分布格局与样点在DCA排序图中的分布格局相似。物种CCA排序第一轴反映了各物种分布海拔情况,黄苞天南星、多裂委陵菜、半球齿缘草、二裂委陵菜、草沙蚕、弱小火绒草、青藏狗娃花等常见的草甸草原植被分布在排序图右侧较低海拔区域,钉柱委陵菜、多刺绿绒蒿(Meconopsishorridula)、西藏三毛草、中亚早熟禾(Poalitwinowiana)等耐寒草甸植被则分布在排序图左侧较高海拔区域,高山嵩草、狭囊薹草等分布在中央,说明这几个植物种的生态适应性比较强,分布较为广泛。结合TWINSPAN分类结果和样点的DCA二维排序图,可以看出,在植物群落中,物种的分布格局与群落类型的分布格局之间具有很大的相关性。

图4 拉萨河谷物种CCA排序图Fig.4 CCA ordination diagram of 54 species in Lhasa River Valley1. 半球齿缘草;2. 矮火绒草;3. 草沙蚕; 4. 垂穗披碱草;5. 大花嵩草;6. 倒披针叶风毛菊;7. 钉柱委陵菜;8. 独一味;9. 多刺绿绒蒿;10. 多裂委陵菜;11. 二裂委陵菜; 12. 高山大戟;13. 高山豆; 14. 高山嵩草; 15. 高山唐松草;16. 禾叶点地梅;17. 禾叶风毛菊; 18. 黑被薹草;19. 横断山风毛菊;20. 厚边龙胆(Gentiana simulatrix);21. 黄苞天南星;22. 金露梅;23. 拉萨厚棱芹; 24. 拉萨黄耆(Astragalus lasaensis);25. 冷地早熟禾;26. 麦薲草;27. 木根香青;28. 扭盔马先蒿;29. 青藏狗娃花; 30. 青藏薹草;31. 全萼秦艽;32. 肉果草; 33. 弱小火绒草; 34. 丝颖针茅;35. 酸模叶橐吾;36. 樱草杜鹃;37. 微药羊茅;38. 无茎黄鹌菜;39. 西藏三毛草;40. 喜马拉雅嵩草;41. 狭囊薹草;42. 线叶龙胆; 43. 线叶嵩草;44. 雪层杜鹃;45. 羊茅;46. 伊凡苔草;47. 颖毛早熟禾;48. 圆穗蓼; 49. 缘毛紫菀;50.云南棘豆;51. 长梗秦艽;52. 中亚早熟禾;53. 珠芽蓼;54. 紫花针茅(Stipa purpurea)
4 结论与讨论
TWINSPAN数量分类将拉萨河谷的草地群落分为8种群落类型,其中青藏薹草群落属于亚高山草甸草原;金露梅+高山嵩草群落、微毛樱草杜鹃+高山嵩草群落、微毛樱草杜鹃+雪层杜鹃+高山嵩草群落、雪层杜鹃+高山嵩草群落属于亚高山灌丛草甸;高山嵩草+青藏薹草群落和高山嵩草群落属于高山草甸。样点DCA排序第一轴基本反映了海拔高度的变化梯度,第二轴基本反映了坡向的变化。TWINSPAN分类所划分的各群落在DCA排序图上都有各自的分布范围和界限,说明DCA排序能较好的反应各群落与其环境资源之间的关系,同时,TWINSPAN的分类结果也在排序图上得到较好的印证。样点CCA排序表明,影响群落分布的主要环境因子是海拔,其次是坡向。CCA排序进一步阐明了拉萨河谷草地群落分布决定于海拔和坡向等环境因子,并间接验证了TWINSPAN的分类结果。物种CCA排序和TWINSPAN分类结果表明:植物群落中物种的分布格局与植物群落类型的分布格局存在一定的相似性,物种的分布格局在很大程度上影响着群落的分布格局。
对拉萨河谷的草地群落分类结果和群落特征表明,拉萨河谷的草地群落分布呈现明显的垂直地带性分布格局,随着海拔的升高,群落类型有亚高山草甸草原逐渐变为亚高山灌丛草甸,最后变为高山草甸,这也与群落的实际分布状况相符。但是,可以看到,分布在海拔4000 m左右的群落被划分为一个单独的群落:垂穗披碱草+草沙蚕群落,该群落的优势种为垂穗披碱草,共优种为草沙蚕,这说明该群落有草原化的趋势,而这恰好是草甸草原退化的表现,我们认为这种退化在很大程度上是由于人类的频繁活动和过度放牧造成的。有学者的研究表明:放牧和人类的干扰会对植物的功能性状产生影响[40];放牧会影响群落的物种组成以及功能群之间的关系和相对优势度,植物的功能性状对放牧活动的响应具有非线性特征[41- 42];长期过度放牧会对植被的空间分布产生较大的影响,导致山地植被空间结构的紊乱,并进一步干扰群落数量分析结果的准确性[43]。他们的研究从群落的物种组成和多样性的角度证明了放牧和人类的干扰对植物的功能性状的影响,本研究从群落结构和分布的角度验证了他们的研究成果。
对于DCA排序轴的生态解释,不同学者的研究结果有所不同,有些学者认为,DCA第一轴反映水分环境梯度,第二轴反映热量环境梯度[44- 47]。部分学者则认为DCA第一轴反映(海拔)热量环境梯度,第二轴反映水分环境梯度[48- 49]。还有学者认为,DCA排序第一轴反映了群落海拔高度的梯度变化,第二轴反映了群落坡度的梯度变化或者坡向的梯度变化[50- 52]。还有一些学者的研究得出了其他的结论。综合学者们的研究,我们可以看出,DCA排序轴的生态解释在不同的研究地点、研究对象和尺度,有可能得出不同的研究结论。在降水比较多地区,水分相对充足,水分条件不会成为植被生长发育的禁锢,或许热量会在较大程度上影响着群落的物种组成和空间分布。在一些比较干旱的地区,水分条件就极有可能影响植被的群落结构和空间分布格局。在干旱荒漠地区,水分极为匮乏,土壤养分也会成为影响群落结构和空间分布的主要因素[46]。对拉萨河谷草地群落的DCA分析结果表明,影响拉萨河谷草地群落物种组成和分布格局的主要环境因子是海拔和坡向。拉萨河谷地区是山原河谷区,山体连绵,由于河谷的特殊地形,使得研究区气候较为干旱,水分条件相对较差。在低海拔地区,气温较高,蒸发量大,土壤含水量低,因此分布群落多为喜温耐旱的草甸草原;在高海拔地区,随着海拔升高,温度降低,水分条件转好,植被发展为灌丛草甸和高寒草甸。海拔反映了温度和水分综合梯度,坡向的不同也反映不同的热量条件,正是热量和水分的综合作用影响着群落的分布格局。因而,影响拉萨河谷草地群落物种组成和分布格局的主要生态因子是海拔和坡向是可靠的。
综上,本文采用TWINSPAN分类结合DCA和CCA排序的研究方法,对青藏高原河谷地区草地群落的数量分析取得了良好的分类结果,并初步揭示了河谷草地植物群落及环境因子的分布格局。可为河谷区草地的保护和利用,以及相关的植被群落研究提供理论依据。
猜你喜欢
湖北植保(2022年4期)2022-08-23
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
土壤(2021年1期)2021-03-23
林业调查规划(2020年3期)2020-06-03
Zoological Research(2019年3期)2019-06-28
乡村地理(2018年4期)2018-03-23
