
低温对羽衣甘蓝白鸽血红素含量及其合成基因表达的影响
2018-09-11 08:33李学来原佳乐刘晓冉孙堃峰史国安
河南农业科学 2018年8期
周 爽,马 超,李学来,原佳乐,刘晓冉,孙堃峰,史国安*
(1. 河南科技大学 农学院,河南 洛阳 471023; 2.洛宁县农业局,河南 洛宁 471700)
血红素(Heme)是一类重要的卟啉类化合物,广泛分布于动物的血液、肌肉以及一些植物组织中。在动物中,血红素主要存在于血液的红细胞中,与珠蛋白结合在一起共同构成血红蛋白;血红素还作为很多酶的辅基参与许多重要的生理功能。在植物中,血红素的结构和叶绿素非常相似,都是四吡咯化合物。在生物合成的前期阶段,从δ-氨基酮戊酸合成到原卟啉IX合成都是血红素和叶绿素所共有的。高等植物中的叶绿素生物合成从谷氨酸开始,整个途径可以分为3步:一是由谷氨酸合成δ-氨基酮戊酸。该过程由谷氨酸tRNA合成酶(HemA编码)、谷氨酸tRNA还原酶(GluTR编码)以及谷氨酸-1-半缩醛氨基转移酶(GSA编码)催化完成,是整个叶绿素生物合成途径的第1个限速步骤。二是由8个δ-氨基酮戊酸合成原卟啉IX。2个δ-氨基酮戊酸在δ-氨基酮戊酸脱水酶(ALAD编码)的作用下生成1个胆色素原,4个胆色素原在胆色素原脱氨酶(PBD编码)的作用下生成1个四吡咯,该四吡咯在尿卟啉原III合成酶(UroS编码)的作用下经过一系列反应生成尿卟啉原III,尿卟啉原III在尿卟啉原III脱羧酶(HemE编码)的作用下生成粪卟啉原III,粪卟啉原III在粪卟啉原III氧化酶(CPO编码)的作用下生成原卟啉原IX,原卟啉原IX在原卟啉原IX氧化酶(PPOX编码)的作用下生成原卟啉IX。三是由原卟啉 IX合成叶绿素。原卟啉IX是叶绿素和血红素生物合成途径中最后一个共同的中间代谢产物。在叶绿素途径,镁离子螯合酶(ChlI、ChlD、ChlH编码)催化镁离子插入原卟啉IX中形成镁离子原卟啉IX,随后镁离子原卟啉IX甲基转移酶(MTF编码)、镁离子原卟啉IX单甲基酯环化酶(MTC编码)、脱植基叶绿素a乙烯基还原酶(DVR编码)、NADPH-原脱植基叶绿素氧化还原酶(POR编码)、牻牛儿牻牛儿酯还原酶(ChlP编码)和叶绿素合酶(CS编码)共同催化合成叶绿素。目前,所有参与高等植物叶绿素生物合成途径的基因均已发现。在拟南芥中,叶绿素合成由26个基因编码的16种酶控制[1-3]。在血红素途径,这一过程由亚铁螯合酶催化,该酶催化亚铁离子插入原卟啉IX,生成血红素[1-3]。亚铁螯合酶由2个同工酶组成,分别由FC1基因及FC2基因编码,2个基因核苷酸差异极大,但功能相同。血红素可以作用于叶绿素生物合成过程的限速酶——谷氨酸-tRNA还原酶,从而抑制δ-氨基酮戊酸合成,并进一步抑制叶绿素的合成[4-5];在拟南芥突变体中的研究表明,由FC1催化合成的血红素可以充当逆行信号,调控一系列与光合作用相关的核基因(Photosynthesis-associatednucleargenes,PhANGs)的表达,如LHCB,RBCS(RuBisCO小亚基)等,从而影响光合作用的发生[6]。而由FC2催化合成的血红素则没有这样的功能[6]。FC1还可以受到各种外界环境的诱导而表达[7]。因此可以推测FC1可能是植物体中响应外界环境变化的重要调控位点,而FC2则主要是组成型表达。血红素合成及叶绿素合成构成了一个复杂的调控网络,因此,研究植物中的血红素合成情况对研究叶绿素代谢调控体系非常重要。
羽衣甘蓝白鸽是十字花科芸薹属甘蓝种中的一个变种,20世纪80年代引入我国,主要作为冬季观赏植物。目前对白鸽的研究主要集中在其观赏价值及栽培方法上。羽衣甘蓝白鸽是一种典型的叶绿素低温突变体,其植株在常温下呈绿色,低温条件下中心部位的嫩叶不能正常合成叶绿素而变为纯白色,是一种优良的适宜深入研究叶绿素代谢过程的突变体材料。前期的研究显示,低温条件下生长的白鸽嫩叶中叶绿素含量远低于常温条件下生长的嫩叶[8]。为明确血红素途径在叶绿素合成、调控中的重要作用,研究了低温对白鸽血红素含量及血红素、叶绿素生物合成相关基因表达情况的影响,探讨血红素合成对叶绿素合成的调控作用,以期为叶绿素代谢调控相关研究提供理论依据。
1 材料和方法
1.1 试验材料
羽衣甘蓝白鸽购自重庆天子林业有限公司。成年植株分别置于温室(平均温度20 ℃)及室外(平均温度5 ℃)培养,分别视为常温和低温处理。15 d后取样。
丙酮、乙醚、盐酸等化学试剂均购自重庆东玻化工有限公司。
引物由华大基因合成,相关分子生物学试剂购自TaKaRa公司及Promega公司。
1.2 血红素提取及测定
准确称量待测植株的新鲜叶片1 g,置于预冷的研钵中,加液氮研磨至粉末状,转入50 mL离心管中。加入10 mL丙酮、蒸馏水混合溶液(V丙酮∶V蒸馏水=99∶1),用力振荡均匀,4 000 r/min离心10 min,弃去上清。重复以上步骤直到离心得到的沉淀物变为无色,再在沉淀物中加入5 mL丙酮、蒸馏水混合溶液(V丙酮∶V蒸馏水=80∶20),用力振荡均匀,4 000 r/min离心10 min,弃去上清。重复这一步骤直到离心得到的沉淀物变为无色为止。此时血红素在沉淀中。在沉淀中加入5 mL盐酸、丙酮、蒸馏水混合溶液(V盐酸∶V丙酮∶V蒸馏水=5∶80∶15),剧烈振荡15 min,4 000 r/min离心10 min,取上清,然后向沉淀中再次加入5 mL盐酸、丙酮、蒸馏水混合溶液(V盐酸∶V丙酮∶V蒸馏水=5∶80∶15),剧烈振荡15 min,4 000 r/min离心10 min,取上清。将两次得到的上清合并,大约有10 mL。在提取液中加入10 mL乙醚,混合均匀后加入分液漏斗中。将分液漏斗倒置几下后静置分层。血红素位于上层,收集备用。在下层溶液中加入10 mL乙醚,再次提取。将两次得到的上层溶液合并在一起,倒入分液漏斗,加入等体积的pH值>4的蒸馏水洗涤。静止分层后取乙醚层,加入旋转蒸发仪中,浓缩至5 mL左右,倒入干净的50 mL离心管中,用氮气吹干。在提取好的干燥样品中依次加入7.44 mL蒸馏水及0.48 mL 5 mol/L NaOH,搅拌至样品溶解后再加入12 mL蒸馏水和4.08 mL pyridine,再次混合均匀。测量时在样品中加入几滴dithionite和50 mmol/L的ferricyanide,测定574.5 nm处的吸光值,依据参考文献[9]的方法计算血红素浓度。每个样本用3株独立植株检测。
1.3 RNA提取及cDNA合成
使用RNAiso Plus(TaKaRa)提取常温及低温处理的羽衣甘蓝白鸽嫩叶RNA。以总RNA为模板,使用M-MLV反转录酶(Promega)合成cDNA第1链。相关操作按照Promega公司说明书进行。
1.4 半定量RT-PCR分析
所检测的相关酶及其编码基因如表1所示。羽衣甘蓝和甘蓝、拟南芥同为十字花科芸薹属,基因序列同源性非常高。根据GenBank上登陆的甘蓝的HemA1、GSA、ALAD、PBD、UroS、HemE1、HemE2、CPO、PPOX、FC1、FC2、ChlI、ChlD、ChlH、MTF、MTC、DVR、ChlP、CS、ACTIN保守序列,以及拟南芥PORB、PORC保守序列,利用Primer Premier 5.0设计半定量引物,详见表2。以肌动蛋白基因ACTIN作为内参基因,用半定量RT-PCR检测白鸽嫩叶中血红素和叶绿素生物合成相关基因在常温及低温条件下的表达情况,重复3次。利用Quantity one v 4.62分析条带亮度,利用Origin 7.5处理数据,利用Microsoft Excel进行单因素方差分析检验。半定量RT-PCR主程序为94 ℃高温变性5 min;PCR循环程序为94 ℃高温变性30 s,54 ℃退火30 s,72 ℃延伸1 min;根据不同基因循环数分别设置26~30个循环。
1.5 数据处理
采用Excel 2007进行单因素方差分析及作图。
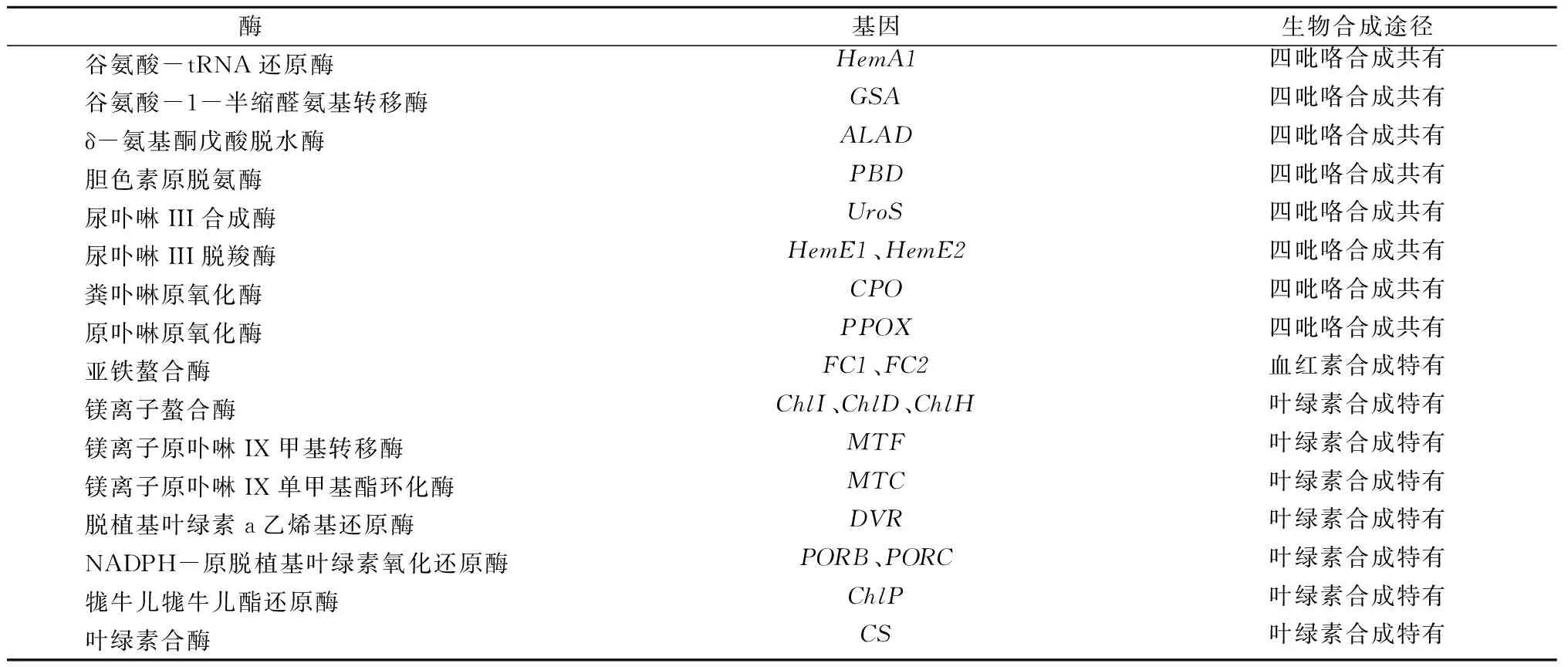
表1 半定量RT-PCR检测的相关酶及其编码基因
2 结果与分析
2.1 温度对羽衣甘蓝白鸽叶色的影响
温度对羽衣甘蓝白鸽叶色影响很大。如图1所示,常温条件下,羽衣甘蓝白鸽嫩叶表现为正常的绿色;低温条件下,羽衣甘蓝白鸽中心嫩叶变为纯白色。羽衣甘蓝白鸽的成熟叶和老叶在常温及低温条件下均为绿色(图1)。
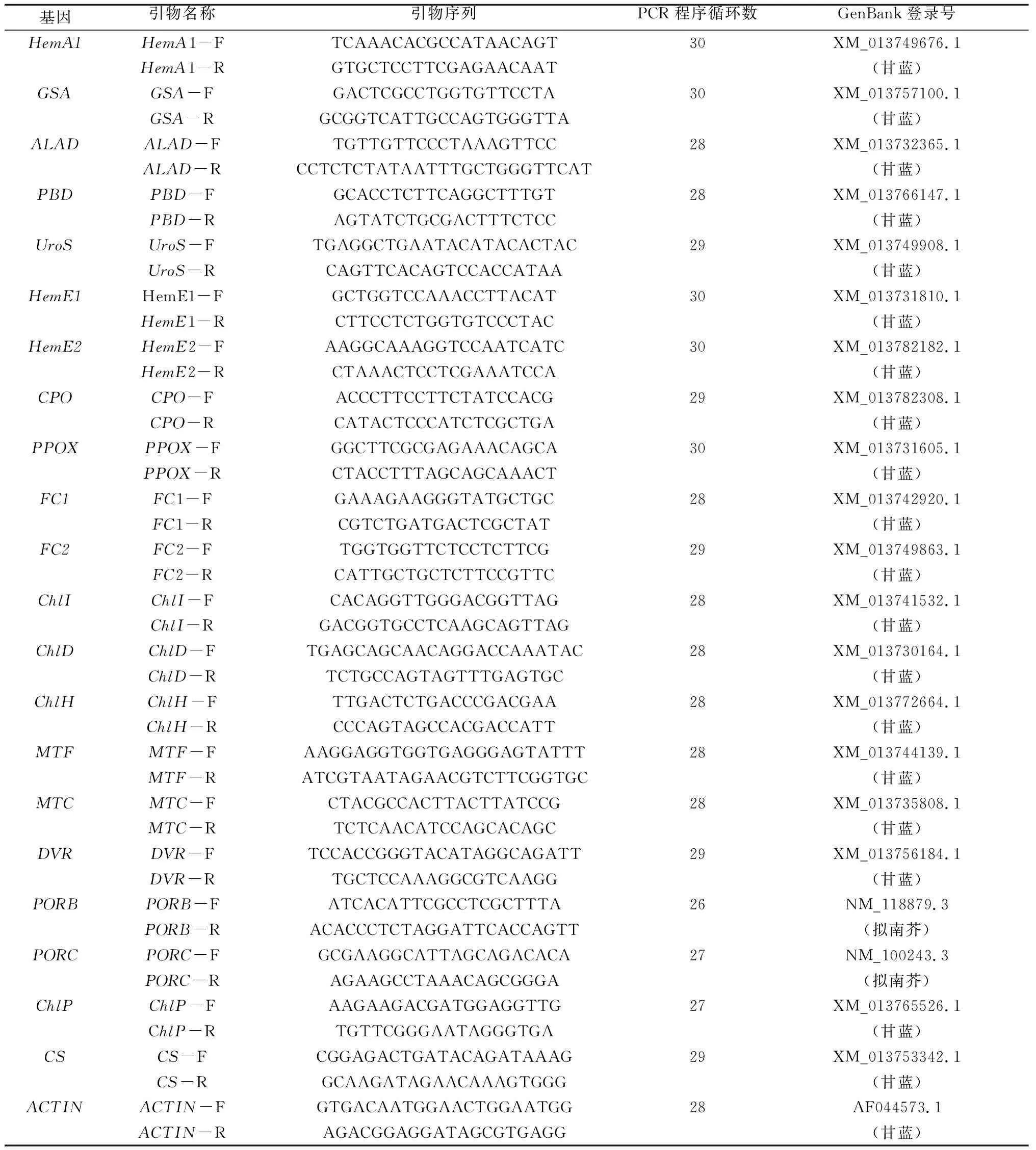
表2 半定量RT-PCR所用引物
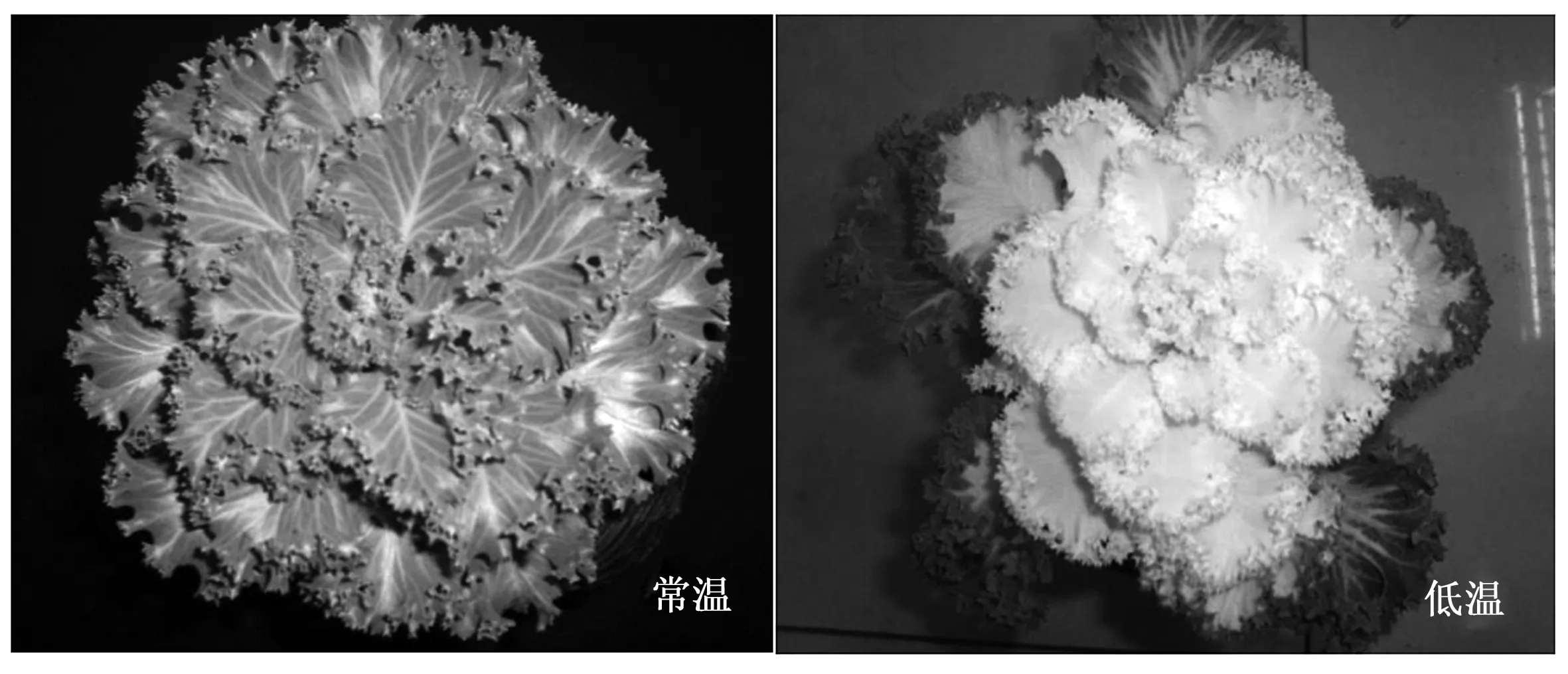
图1 温度对羽衣甘蓝白鸽叶色的影响
2.2 低温对羽衣甘蓝白鸽嫩叶中血红素含量的影响
血红素和叶绿素的前期合成过程完全重合,后期才发生分支,因此血红素合成过程对叶绿素合成有重要的调节作用。准确测定样品中的血红素含量对研究叶色突变体的叶绿素合成过程有重要意义。结果如图2所示,白鸽嫩叶中的血红素含量在低温条件下为(0.021 2±0.007 01)mg/mL,在常温条件下仅为(0.000 973±0.000 542 8)mg/mL,且差异极显著。
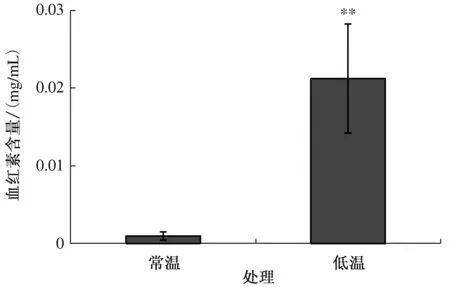
*表示差异显著(P<0.05);**表示差异极显著(P<0.01);下同
2.3 低温对白鸽嫩叶中四吡咯化合物共有合成基因表达的影响
在四吡咯化合物的合成过程中,共有9个基因编码的8种酶是叶绿素及血红素合成所共有的(表1)。如图3所示,9个四吡咯化合物共有合成基因的表达水平都在低温条件下略有下调,但其表达差异不显著。HemA1、GSA、ALAD、PBD、UroS、HemE1、HemE2、CPO、PPOX的表达水平在低温及常温条件下没有明显差异。低温及常温条件下相对表达量差异最小的是CPO,常温条件下为1 928.333 3±427.911 6,低温条件下为1 854.666 7±452.178 4,下调了3.82%;相对表达量差异最大的是GSA,常温条件下为2 050±485.077 3,低温条件下为1 726.666 7±382.796 7,下调了15.77%;但差异均未达到显著水平。
2.4 低温对白鸽嫩叶中血红素合成特有基因表达的影响
血红素生物合成途径特有的基因FC1和FC2的表达情况如图4所示。FC1的相对表达量在低温条件下为2 000±160.934 8,在常温条件下为1 440±170.587 2。FC1的表达水平在低温条件下明显上调,差异显著;FC2的相对表达量在低温条件下为1 650±213.775 6,在常温条件下为1 743.333 3±212.210 6,差异不显著。
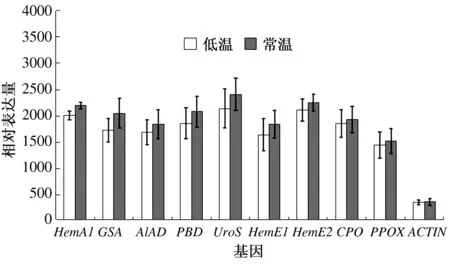
图3 不同温度条件下白鸽嫩叶中的四吡咯化合物共有合成基因表达
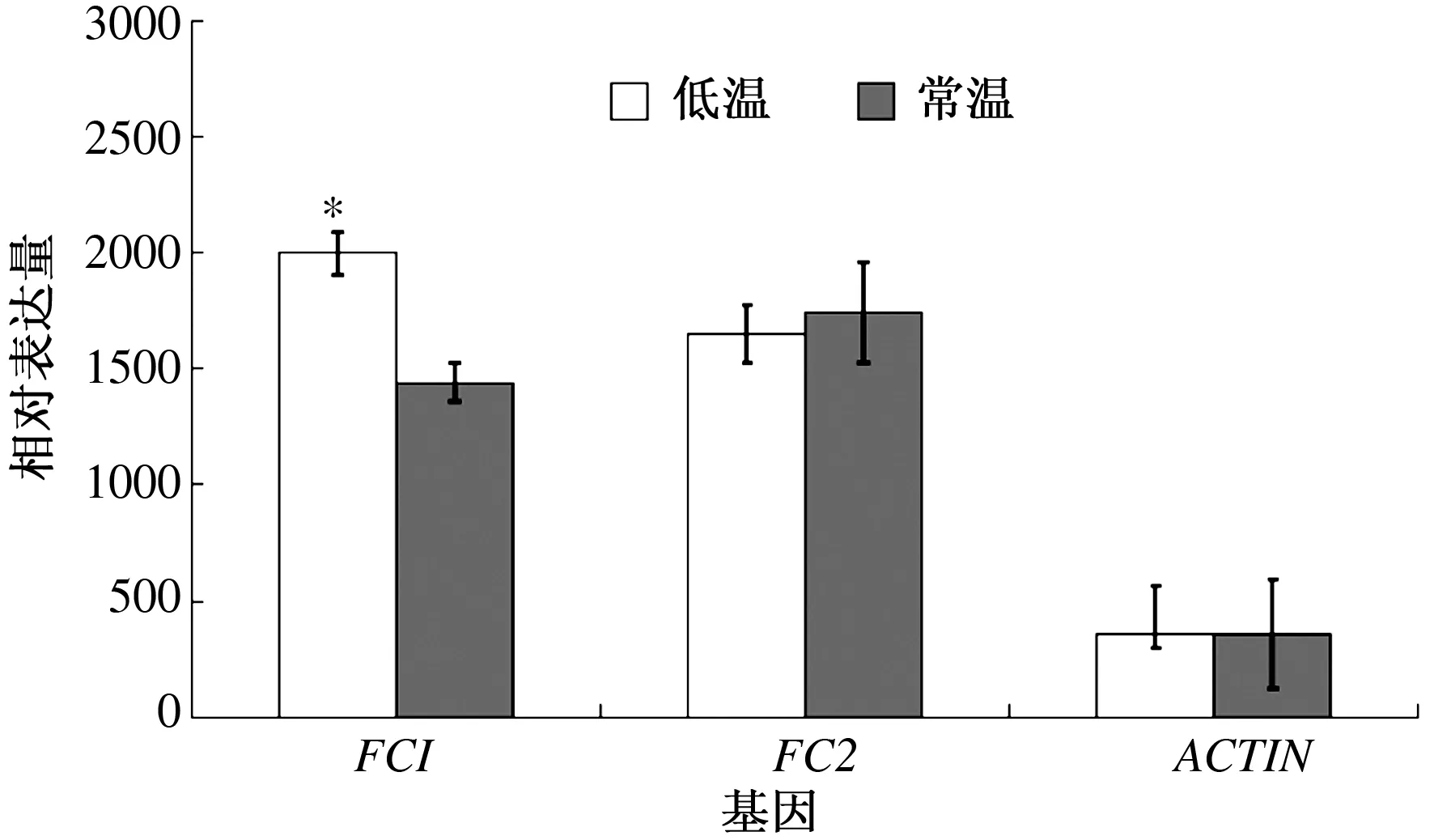
图4 不同温度条件下白鸽嫩叶中的血红素合成特有基因的表达
2.5 低温对白鸽嫩叶中叶绿素合成特有基因表达的影响
叶绿素生物合成途径特有的基因ChlI、ChlD、ChlH、MTF、MTC、DVR、ChlP、CS的半定量RT-PCR结果如图5所示。所有检测的叶绿素合成特有基因的表达在低温条件下都下调,特别是PORB、PORC、ChlP。PORB在低温下的相对表达量为1 973.333 3±153.079 5,在常温下的相对表达量为3 163.333 3±70.237 7,差异极显著;PORC在低温下的相对表达量为1 916±101.528 3,在常温下的相对表达量为2 950.666 7±105.457 7,差异极显著;ChlP在低温下的相对表达量为1 768.333 3±107.973 8,在常温下的相对表达量为2 270±175.214 2,差异显著。其他基因的表达水平在不同温度条件下差异不显著。
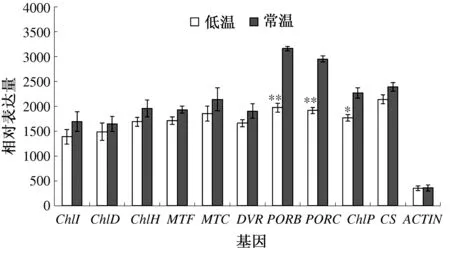
图5 不同温度下白鸽嫩叶中的叶绿素合成特有基因的表达情况
3 结论与讨论
血红素分子在调控叶绿素合成过程有重要作用[10-11]。细胞中的血红素还可作为核质信号调控一系列PhANGs的表达[6, 12]。叶绿素生物合成相关基因的表达模式差异极大,一部分基因在任何发育时期、任何外界环境下表达稳定,是组成型表达;而另一部分基因则容易受到发育阶段及外界环境影响,其表达水平在不同发育时期、不同外界环境下差异较大,是外界环境调控叶绿素代谢的位点。目前在叶绿素生物合成过程中已经发现了3个具有高度不同表达模式的酶,分别是:谷氨酸-tRNA还原酶(由HemA基因编码)、镁螯合酶(由ChlI、ChlD和ChlH基因编码的三亚基组成)和NADPH-原脱植基叶绿素氧化还原酶(由POR基因编码)。其中,镁螯合酶的活性非常容易受到外界环境的影响,编码NADPH-原脱植基叶绿素氧化还原酶的POR基因在不同发育阶段及不同光照条件下表达差异很大。因此,这些酶在植物应对外界和内部条件变化的过程中发挥着重要的作用。
本研究发现,白鸽嫩叶中的血红素含量在低温条件下远高于室温条件下。有研究[8]表明,白鸽嫩叶中的尿卟啉原III含量在低温与室温下没有明显的差异,粪卟啉原III的含量在低温条件下甚至略微增加,原卟啉IX和叶绿素含量则在低温条件下急剧降低。本研究半定量RT-PCR的结果表明,叶绿素和血红素合成共有的基因,其表达水平在低温和常温条件下差异不显著,这一结果与四吡咯合成共同前体尿卟啉原III及粪卟啉原III的含量在低温和常温条件下没有显著差异相一致[8]。低温条件下血红素合成途径特有的FC1表达水平显著提高,而叶绿素合成途径特有的PORB、PORC、ChlP的表达水平则显著降低,可以推测其共同的底物原卟啉IX流向了血红素合成途径,使低温条件下嫩叶中的血红素含量急剧增加,高水平的血红素可以对叶绿素合成进行负调控,进一步抑制了叶绿素合成及光合作用,导致白鸽嫩叶在低温条件下叶绿素含量急剧降低,变为白色。
在叶绿素合成过程中,NADPH-原脱植基叶绿素氧化还原酶催化原脱植基叶绿素还原为脱植基叶绿素,这是整个叶绿素合成过程中唯一需要光的步骤[13]。δ-氨基酮戊酸合成是叶绿素合成过程的限速步骤,δ-氨基酮戊酸合成率和光活性的原脱植基叶绿素的水平成反比[14]。在黑暗条件下,叶绿素合成过程只进行到形成原脱植基叶绿素,一旦原脱植基叶绿素的含量达到临界值,δ-氨基酮戊酸合成就会放缓[15]。NADPH-原脱植基叶绿素氧化还原酶通过与原脱植基叶绿素及NADPH组成三元复合物的方式来行使其功能[16]。结合在NADPH-原脱植基叶绿素氧化还原酶活性部位的原脱植基叶绿素是活化的,在光照下会立刻被转变为脱植基叶绿素[17]。而没有结合在NADPH-原脱植基叶绿素氧化还原酶活性部位的原脱植基叶绿素是非活化的,有可能诱导产生单态氧[18]。在拟南芥中发现了3种不同的NADPH-原脱植基叶绿素氧化还原酶基因,分别命名为PORA、PORB、PORC。其中,PORA在黄化幼苗刚刚被光照射时高效表达,在幼苗萌发时也可以检测到[19]。本研究中白鸽为成年植株,因此没有检测PORA的表达情况。PORB在黑暗和光照条件下的表达几乎是恒定的[19-20]。PORC在光照条件下已经转绿的幼苗中表达增强,并主要在绿色植株中表达[21]。PORA、PORB、PORC的功能有部分重合,并在不同的环境条件下可以互补[22],说明NADPH-原脱植基叶绿素氧化还原酶是外界环境调控叶绿素合成过程的关键点,PORA、PORB、PORC的互补性可以帮助植物适应各种外界环境。PORA和PORB与前片层体的典型结构关系密切[23],在叶绿体发育过程中发挥着举足轻重的作用。最近的研究[24]发现,NADPH-原脱植基叶绿素氧化还原酶还参与了FLU蛋白对叶绿素合成途径的负反馈调控。
低温条件下,白鸽FC1的表达明显受到低温诱导,FC2的表达水平则没有明显差异,表明白色嫩叶中急剧增加的血红素主要由FC1催化合成。白鸽中的PORB和PORC表达水平在低温下显著降低,而在其之后的ChlP的表达水平也随之降低。这些结果表明FC1、PORB、PORC可能在白鸽低温白化的过程中起关键作用,彼此之间存在调控联系。
猜你喜欢
东坡赤壁诗词(2022年4期)2022-10-30
昆明医科大学学报(2021年5期)2021-07-22
江苏科技报·E教中国(2021年4期)2021-05-08
中国石油大学学报(自然科学版)(2018年1期)2018-03-16
吉林农业·下半月(2016年11期)2017-01-09
江苏农业科学(2016年7期)2016-10-20
医学研究杂志(2015年9期)2015-07-01
江苏农业科学(2014年6期)2014-08-12
食品工业科技(2014年13期)2014-03-13
食品工业科技(2014年18期)2014-02-27
