
环境胁迫植物富集γ-氨基丁酸的研究进展
2018-10-16 09:53丁俊胄贾才华赵思明张宾佳熊善柏林亲录
食品工业科技 2018年18期
王 斌,丁俊胄,2,贾才华,*,赵思明,*,牛 猛,张宾佳,熊善柏,林亲录
(1.华中农业大学食品科学技术学院,湖北武汉 430070;2.伊利诺伊大学香槟分校食品与人类营养学系,美国伊利诺伊州厄巴纳市 61801;3.中南林业科技大学食品科学与工程学院,湖南长沙 410004)
γ-氨基丁酸(Gama-aminobutyric acid,GABA)是一种四碳非蛋白组成氨基酸,在神经系统中是一种重要的神经活性抑制剂,具有诸多功效,例如降低血压、治疗失眠、缓解焦虑、增强免疫力以及增加记忆力等作用[1]。我国卫生部2009年批准γ-氨基丁酸为新资源食品,建议成人每天摄入500 mg以内。随着人们年龄的增长,人体通过自身代谢生成GABA的能力降低,加之日常饮食中GABA含量摄入量不够,人体GABA含量较低。
GABA广泛分布于植物中,但含量普遍较低,当植物受到环境(低氧、低温、热、盐渗透、机械损伤)刺激时,会大量生成GABA。在食品生产中,可通过改变外界环境刺激植物组织,促进GABA的合成,制得高GABA食品,如茶叶、发芽糙米、豆芽等。
高GABA食品能够有效增加人体GABA含量,对于保持人体健康具有重要意义。本文将从GABA的代谢机制、环境胁迫对GABA积累的影响以及几种胁迫方式在高GABA食品研发中的应用做一个综述。
1 γ-氨基丁酸代谢途径
高等植物代谢积累GABA有两种途径。一种是以谷氨酸(L-Glutamine,Glu)作为底物,经谷氨酸脱羧酶(Glutamate decarboxylase,GAD,EC 4.1.1.15)催化生成GABA,该途径被称为GABA旁路或GABA支路[2]。另一种则是以多胺(polyamines)为底物,经过二胺氧化酶(DAO)、多胺氧化酶(PAO)催化,可将多胺(腐胺、亚精胺)转化为GABA,该途径被称为多胺降解[3-4]。
1.1 GABA支路
GABA支路广泛分布于真核生物和原核生物中,是GABA重要的代谢途径。如图1所示[1],在细胞质中,谷氨酸首先被GAD催化不可逆生成GABA,随后GABA由GABA转氨酶(GABA transaminase,GABA-T)催化转化为成琥珀酸半醛半醛酸(Succinic acid,SSA),琥珀酸半醛进入线粒体,由琥珀酸半醛脱氢酶(Succinate-Semialdehyde Dehydrogenase,SSADH)氧化生成琥珀酸(Succinic acid,SSC),随后琥珀酸进入三羧酸循环[2]。
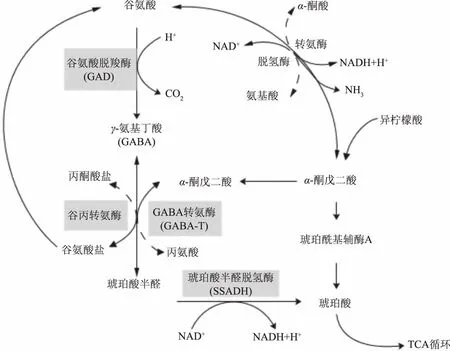
图1 GABA支路代谢路径图Fig.1 GABA shunt and its relationship to other metabolic pathways
1.2 多胺降解
多胺降解也是生成GABA的途径之一。相较于GABA支路,多胺降解产生的GABA含量较少,在以往的研究中,大部分都是以GAD活性作为关键因素来探究GABA在植物体内代谢情况。但最近的研究发现,在某些植物中多胺的含量与GABA含量有密切联系,通过多胺降解产生的GABA占有较大比例。Yang等[5]用氨基胍对大豆进行处理发现,氨基胍能显著增加多胺含量,但GABA含量下降了30%。另外,Wang等[6]用外源GABA处理甜瓜根,发现处理后甜瓜根的多胺含量显著增加,说明多胺代谢与GABA代谢有密切联系。
多胺降解产生GABA路径如图2所示,谷氨酸(L-Glutamic acid,L-Glu)经过转氨酶生成鸟氨酸(Ornithine,Orn)和精氨酸(Arginine,Arg),鸟氨酸和精氨酸经过鸟氨酸氧化酶(Ornithine decarboxylase,ODC)和精氨酸氧化酶(Arginine decarboxylase,ADC)氧化生成腐胺(Putrescine,Put),随后腐胺经亚精胺合成酶催化转化成亚精胺(Spermidine,Spd),亚精胺进一步转化为精胺(Spermine,Spm)。腐胺和亚精胺分别经二胺氧化酶(Diamine oxidase,DAO,EC 1.4.3.6)和多胺氧化酶(Polyamine oxidase,PAO,EC 1.5.3.3)氧化生成4-氨基丁醛,最后由氨基丁醛脱氢酶(4-aminobutyraldehyde dehydrogenase,ABALDH)转化为GABA[4,7-8]。
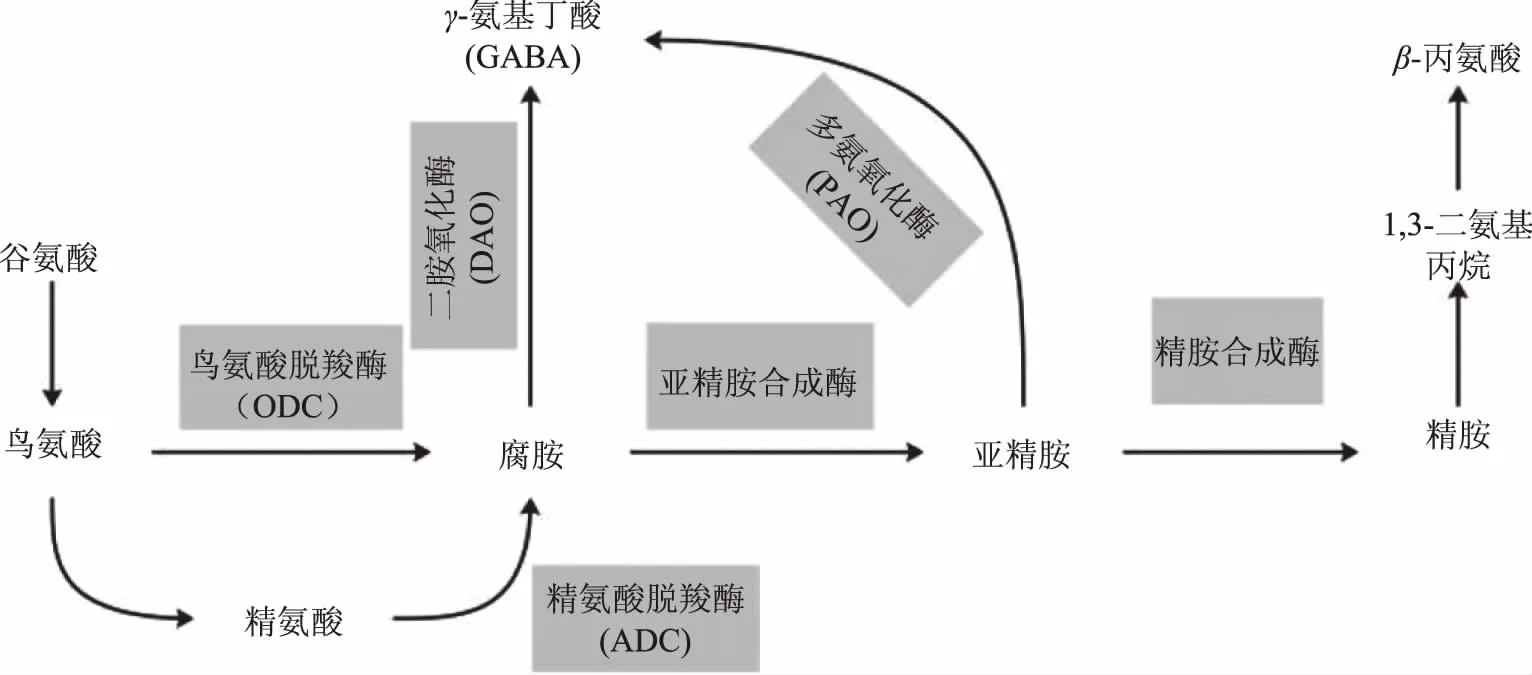
图2 多胺降解生成GABA代谢路径Fig.2 The production of GABA through polyamine degradation
1.3 环境胁迫促进GABA富集机理
当外界环境发生改变,不利于植物生长时,会激发植物内源酶活性,进而大量产生GABA,从而使植物适应外界的变化。GABA对于植物在逆境环境下的生存具有重要意义。
GABA在植物体内主要有以下作用:a、调节pH,当植物在无氧或受到机械损伤时,细胞质内会产生大量的H+,从而导致细胞质酸中毒,GAD在代谢的过程中需要消耗H+,减缓酸中毒,从而起到调节pH的作用[9];b、氮素储藏,GABA代谢的过程中会消耗大量的谷氨酸,谷氨酸中的氮素被临时转移到GABA,通过GABA支路代谢,氮素被重新用于蛋白合成[10];c、抵御干旱:GABA会使植物气孔减小,从而减少水分的流失,进而具有更强的抗旱能力[11];d、防止虫害,当受到虫害时,植物液泡中的有机酸被释放从而激发GAD的活性,促进GABA的产生,研究发现一定量GABA能够减缓幼虫的成长发育、降低其存活率[12-13];e、抵抗霉变,经GABA处理后的果实,其抗霉变相关的酶的基因表达及活性会增强,提高果实的抗霉能力[14]。
1.3.1 GABA支路 GAD是GABA支路代谢的限速酶,其活性与不同物种来源、环境条件差异等有密切联系[15-16]。GAD的活性主要由pH和钙离子/钙调蛋白(CAM)结合调控[17]。在受到环境刺激时(低氧、低温、酸刺激、高盐渗透等),植物体内H+和Ca2+会大量增加,H+和Ca2+对于GAD的活性均有促进作用。大部分GAD在酸性环境下具有较大活性,只有少部分GAD在碱性环境下能保持较大活性,微生物GAD的最适pH一般在3.8左右[18],而植物GAD的最适pH则在5.8左右[19]。
Ca2+也是影响GAD活性的重要因素,GAD是一种钙调素协调蛋白,含有CAM结合蛋白,能够与Ca2+结合形成Ca2+/CAM。GAD的氨基酸组成中C端含有22~25个氨基酸残基,形成的空间结构能够与Ca2+/CAM结合,起到激活GAD的作用[20],进而促进GABA的合成。但Ca2+仅在与CAM共同作用下才会激发GAD的活性。植物组织受到外界刺激(冷激、热激、机械损伤、渗透压胁迫等)时,细胞质中的Ca2+会大量增加,激活GAD,促进GABA合成。
1.3.2 多胺降解 多胺降解生成GABA的代谢酶包括ADC、ODC、DAO、PAO以及ABALDH等[21-22]。多胺降解生成GABA的过程可以分为两个步骤:多胺的生成和多胺的降解。多胺的生成主要由ADC和ODC来调控。ADC能催化精氨酸生成鲱精氨,进而转化为腐胺。ADC在植物界中分布广泛,已发现的编码基因有ADC1和ADC2[23]。ADC在所有的植物组织中均有表达,在受到冷激作用下会促进ADC1的表达,而ADC2仅在植物组织受到外界刺激(冷冻、干燥、高盐渗透、机械损伤等)时,会促进其表达[3]。ODC是一种磷酸吡哆醛依赖酶,能够直接催化鸟氨酸生成腐胺,主要分布于动物组织内,在植物中表达程度较低,合成的基因有ODC1和ODC2,其中ODC2仅存在于人体。ODC在逆境胁迫下活性变化较小,其主要在细胞分裂及发育的过程中发挥作用[24]。受到外界刺激时,会促进ADC和ODC的表达,提高ADC和ODC的活性,进而促进多胺的积累[25]。
多胺的降解主要受到PAO和DAO的调控。DAO和PAO可分别将腐胺和亚精胺转化为氨基丁醛,然后经ABALDH催化生成GABA[4]。通常,DAO在植物体内的表达程度较低,但是当受到外界环境刺激时会促进DAO的表达增加其活性,将积累的多胺转化为GABA[25]。
2 环境胁迫处理在富集GABA中的应用
2.1 低氧胁迫
低氧条件下,GABA被认为是一种信号分子,能够调节细胞质pH,抵抗氧化性损伤[26-27]。低氧是胁迫植物富集GABA最常用的手段之一,具有操作简单、成本低、效果明显等优势,在植物富集GABA中有广泛的应用。针对不同原材料,低氧处理方式获得GABA含量虽有较大的差异,但较未处理组,GABA含量可提高5~30倍不等。杨润强等[28]先用pH3.5的营养液浸泡蚕豆培养36 h,随后低氧联合NaCl胁迫培养4 d,蚕豆GABA含量可达1.06 mg/g(Dw),较空白GABA提高了7.57倍。尹永祺等[29]以玉米为原材料,在通气量为1.0 L/min的条件下,胁迫培养3 d,GABA含量提高了12.2倍。黄亚辉等[30]分别用液体浸泡和真空充氮的方式胁迫茶叶富集GABA,茶叶的GABA含量分别达到4.01 mg/g和2.88 mg/g,提高了14倍和30倍。除了营养液的方式进行低氧胁迫,丁俊胄等[31]用CO2和N2真空充气的方式对糙米进行低氧胁迫,发现CO2的胁迫效果要优于N2,且胁迫介入的时期及时间长短对富集GABA的效果也有显著差异。Ding等[32]在使用“黑糯”和“先恢”两个高GABA水稻亲本研究中,发现常规发芽72 h后GABA含量是未发芽的2.8倍和6.4倍,而在66~72 h期间经CO2低氧处理后的样本GABA含量分别是未发芽糙米的38.5倍和37.6倍。
以往大部分的研究认为,低氧胁迫下,植物主要是通过 GABA支路的方式富集。植物在低氧胁迫下,细胞质中的H+大量增加,促进了GAD的代谢转化。尹永祺等[33]以发芽玉米为实验材料,检测低氧胁迫下玉米发芽过程中GAD、DAO、AMADH的活性,发现发芽过程,GAD活性先上升后下降,始终高于对照组,而DAO总体呈下降趋势,AMADH呈稳定趋势。玉米发芽过程中,多胺降解酶始终处于较低的活性,而GAD则处于较高的活性,表明在该过程中是GABA支路起主要作用。但最近的研究发现由多胺代谢所产生的GABA占了相当大的比例,氨基胍对多胺氧化酶有特异性抑制作用,郭元新等[34]在低氧胁迫下用2.5 mmol的氨基胍抑制大豆中DAO的活性,而对GAD等酶无抑制作用,其富集GABA的含量降低了32%,说明大豆富集的GABA中有32%来自于多胺降解。
2.2 高盐渗透
高盐渗透也是富集GABA的手段之一,植物组织在培养的过程中添加高浓度盐溶液,可以促进植物富集GABA。NaCl是高盐胁迫中常用的胁迫剂。盐胁迫的研究主要集中在高盐对GABA支路的影响。盐胁迫能够提高植物体内GAD的活性,曾晴等[35]用不同浓度的NaCl处理大豆,发现高盐渗透条件下,GAD活性和GABA含量都随着NaCl浓度的增加而升高,当NaCl浓度为100 mmol/L,GABA含量可达269.93 mg/100 g。同时,盐胁迫作用在玉米、糙米,苦荞等植物中也能起到类似的促进作用[36-39]。盐胁迫除了能提高GABA支路的积累量,对一些植物的多胺降解途径也有一定程度的提高,Xing等[40]以大豆为原材料,通过盐胁迫处理,发现随着NaCl浓度的提高,DAO的活性增强,多胺(Put、Spd和Spm)含量降低,GABA含量提高了17倍。
2.3 低温胁迫
低温胁迫是指在较低的温度下储藏处理植物,在该过程中会增加植物组织中GABA的含量。植物为了增强其抗冻能力,通过GABA支路提供能量,减少活性氧积累[27]。低温胁迫是一种操作简单、高效的处理方式,一般不会单独使用,会在植物种子发芽前后或联合其他胁迫方式共同使用。马丽等[41]采用低温胁迫后的糙米进行发芽实验,结果显示经低温胁迫后的发芽糙米GABA含量高,为原材料(未冷冻处理未发芽组别)的2.58倍,是发芽组(未冷冻处理)的1.19倍。白青云等[42]将马铃薯用营养液浸泡后,用-24 ℃和-68 ℃冷冻,随后微波解冻,结果显示-24 ℃处理要比-68 ℃处理GABA含量更高,GABA含量可达51.37 mg/100 g,为原材料5.61倍。低温胁迫与其他胁迫方式联用可以进一步增加植物中GABA的含量,尹永祺等[43]将低氧处理后的发芽玉米,分别置于5、-5、-18 ℃进行低温胁迫处理,解冻4 h后测定GABA含量,发现在-18 ℃条件下GABA含量最高,约为原材料(未低氧未低温胁迫处理)GABA含量的30倍,同时较、单一低氧处理GABA含量提高36%。Wang等[44]用NO处理冷藏香蕉,发现低温储藏条件下,NO可以增加DAO酶活性同时降低GABA-T活性,促进GABA的积累。综上,低温胁迫对两种代谢途径均有作用,可通过增加GAD活性、DAO活性或降低GABA-T活性等方式促进植物中GABA的积累。
2.4 超声波胁迫
超声波技术常用于提取植物天然活性成分,提高植物种子发芽率,食品的清洗灭菌等。郑艺梅等[45]采用28、40 kHz的超声波分别处理发芽糙米,能够显著提高其GABA含量,约为原材料的3倍。Yang等[46]报道了浸泡过程中超声波处理可提高黄豆芽中的GABA含量,使用频率为40 kHz、功率300 W超声波处理30 min的样品组GABA含量比对照组高出43.39%。张祎等[47]报道了在糙米发芽17 h时采用频率30 kHz的超声波处理16 min,GABA含量达到77.56 mg/100 g。Ding等[48]研究25 kHz超声波处理不同发芽时段糙米中GABA积累量,发现发芽时间0、66 h的糙米,经5 min超声处理后均可显著提高GABA含量,分别较未超声处理组提高60.87%和69.24%。
2.5 其他胁迫方式
除了低氧、高盐、低温等胁迫方式,其他方式如高温热激、酸性刺激等处理方式均能有效增加植物GABA的含量。Young-Su Youn等[49]采用低氧联用热处理方式,低氧处理后经120 ℃处理30 s,胁迫两种小麦发芽富集GABA,结果表明联用胁迫方式处理的,两种小麦GABA含量较、单一低氧处理组分别提高了1.82 mg/100 g和2.23 mg/100 g。Li等[50]采用弱酸性电解水处理粟谷发芽,结果显示弱酸性电解水可促进粟谷GABA积累,较原材料GABA含量提高21%。Hyun Soo Kim等[51]采用低氧、酸刺激等方法联用处理糙米、米糠,均可以促进GABA的合成,相较未处理组,GABA含量提高了17倍,达523 mg/100 g。
3 结论
GABA支路和多胺降解均是植物GABA代谢的重要途径,对植物在环境胁迫下生长起着至关重要的作用。与GABA支路途径相比,多胺降解途径生成的GABA含量虽然较低,但对于植物在环境胁迫条件下积累GABA研究具有重要意义。此外,不同胁迫方式对植物GABA含量及代谢机制均有较大差异,研究不同胁迫方式对植物积累GABA的影响,可用于研发高GABA食品。联用不同胁迫方式可进一步促进GABA积累,但其涉及的不同代谢途径及其相互作用机理尚不明确,有待进一步研究。
猜你喜欢
粮食与饲料工业(2022年5期)2022-10-28
西安石油大学学报(自然科学版)(2022年5期)2022-10-08
新疆农业科学(2022年2期)2022-04-01
蔬菜(2018年11期)2018-01-16
电信科学(2016年9期)2016-06-15
江苏农业科学(2016年4期)2016-06-14
电测与仪表(2016年13期)2016-04-11
电工技术学报(2014年7期)2014-11-15
食品工业科技(2014年13期)2014-03-11
中国饲料(2013年7期)2013-01-25
