
海河天津段微囊藻及其毒素的空间分布及与水环境因子的关系
2018-10-29 05:00王素炎陈裴裴李一鸣乔之怡
中国环境科学 2018年10期
霍 达,曹 琪,王素炎,陈裴裴,李一鸣,乔之怡*
海河天津段微囊藻及其毒素的空间分布及与水环境因子的关系
霍 达1,2,曹 琪1,王素炎1,陈裴裴1,李一鸣1,乔之怡1,2*
(1.天津农学院水产学院,天津 300384;2.天津市水产生态及养殖重点实验室,天津 300384)
采取荧光定量PCR技术及ELISA酶联免疫吸附法,以基因和-基因为靶基因对夏季蓝藻暴发期间海河天津市区段的产毒微囊藻种群丰度和毒素含量进行了研究.结果表明:夏季海河天津市区段微囊藻种群丰度具有明显差异性:产毒微囊藻种群丰度为1.16×104~2.48×107copies/mL占总微囊藻种群的4.25%~28.59%.藻毒素含量最高点为8号采样点天津站,每升水体中藻细胞共含毒素195.51μg,除去藻细胞水中毒素浓度为0.97μg/L.总的来说,海河天津市区段微囊藻总基因拷贝数较高,产毒微囊藻丰度在不同采样点间差异较大,影响水体中微囊藻丰度的最主要环境因素是pH值.海河水体中毒素含量没有超过安全阈值,但是单位体积水体中藻细胞内毒素含量很高,具有较高的毒素释放潜力,可能对下游河口生态系统造成潜在威胁.
微囊藻;微囊藻毒素;荧光定量PCR技术;ELISA酶联免疫吸附法;海河
微囊藻水华给淡水环境带来了极大的影响.一些种类的微囊藻能产生次生代谢物微囊藻毒素(MC),该毒素是富营养化水体中最主要的蓝藻毒素类型,可引发水生生物中毒甚至死亡,直接威胁水中鱼类及其它生物的生存.饮用水源中的微囊藻毒素会对人类的肝脏造成损伤,影响人类的健康和生存[1],同时,微囊藻毒素是强致癌剂,与人群原发性肝癌的高发关系密切[2].为保护人类身体健康,世界卫生组织规定的饮用水中微囊藻毒素安全浓度阈值<1.0μg/L[3].研究表明,微囊藻水华种群中同时含有有毒和无毒微囊藻基因型,两者种群丰度及其比例的动态变化直接影响了微囊藻水华毒素浓度,对微囊藻毒素浓度具有很好的指示作用[4-6].有毒和无毒微囊藻细胞形态一致,用传统的显微镜形态观察方法无法对两者进行区分和鉴定.有毒微囊藻细胞基因组中含有微囊藻毒素合成酶基因家族(mcy)[7-8],对毒素产生进行调控,而无毒微囊藻株中则不含有mcy 基因家族.因此可以应用PCR扩增mcy基因单个或多个成员检测和鉴别有毒和无毒藻株[9-11].
近些年来,随着分子生物学技术的发展,越来越多的研究人员对于微囊藻的产毒相关基因进行了定量的研究.Rinta-Kanto等[12]以基因和微囊藻16S rDNA基因为靶基因采用荧光定量PCR的方法分析了Western Lake Erie的产毒微囊藻构成及其丰度,发现了该湖中微囊藻存在产毒和非产毒共存的现象,在国内的研究中,学者们[13-15]对太湖及巢湖水华期间微囊藻种群丰度进行了研究,发现了相同现象在中国南方水体中的广泛存在.刘洋等[16]对南太湖入湖口的产毒微囊藻进行了研究,证明了该方法可以快速准确地检测水体中微囊藻产毒藻数量.
随着全球气温变化的影响,蓝藻暴发的范围慢慢由南方水体延伸至北方.北方水体具有冬季结冰,四季气温变化大等特征,与南方水体差异较大.已有研究表明在温度[17]、纬度[18]等因素影响下,微囊藻毒素的释放具有不同特征.海河贯穿天津市区,随着近年来两岸生活污水的排放,以及上游水质的恶化,海河市区内景观河道出现了大面积的微囊藻水华,对生活用水造成了较大的威胁.本研究将天津海河作为模型,从毒素合成调控基因和毒素两方面入手,研究北方缓速流动型水体的毒素释放特征,为水环境治理提供参考资料.
1 材料与方法
1.1 采样位点及化学指标测定方法
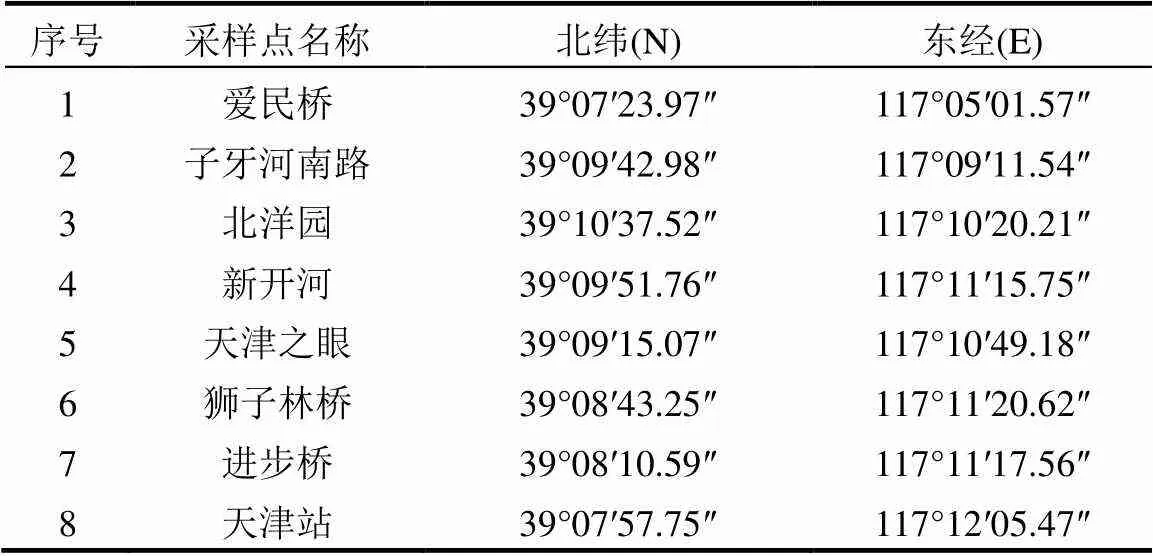
表1 采样点信息
2015年7月,在海河天津市区段(坐标)设置8个采样点(表1),分别为天津站、耳闸摩天轮、天津湾公园富民桥、狮子林桥.使用不锈钢采水器采取1.5m水柱,混合均匀.同时利用YSI多功能水质检测仪测定采样点水质基本参数如温度、pH值、氨氮、溶解氧等.TN、TP测定方法参照文献[14].
1.2 基因组DNA提取
取200mL混匀后的水样(根据微囊藻的量调整抽滤体积)使用Millipore玻璃纤维膜过滤.含藻滤膜置于-20℃保存.将膜剪碎后采用Rinta-Kanto等[12]的方法提取DNA.
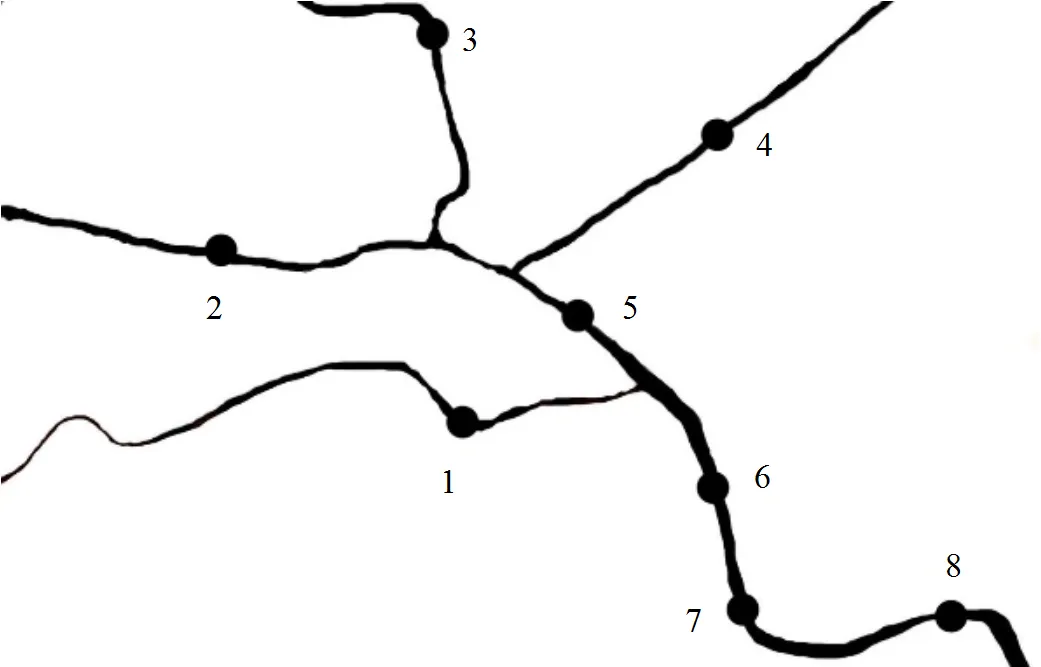
图1 采样点示意
1.3 构建荧光定量PCR 标准曲线
荧光定量PCR所选用的引物如表2所示,其中,引物188F和254R用于特异性扩增微囊藻藻蓝蛋白转录间隔区PC-IGS[4],引物F2和R2用于特异性扩增有毒微囊藻毒素合成基因[19].标准藻株为有毒微囊藻株FACHB 905,由中国淡水藻种库(FACHB)提供,在实验室内培养,培养条件(光照强度为30mmol protons/(m2×s),光照周期12h/12h (L:D),藻细胞浓度培养到对数生长期进行采收).采用全细胞PCR的方法扩增2个片段,将PCR产物进行琼脂糖电泳后回收.将纯化后的产物与pEASY®克隆载体连接,并注入大肠杆菌感受态细胞Trans1-T1中,于LB液体培养基中振荡培养;离心收集菌液,混匀,取适量涂布在含有LB(Amp+)的固体培养皿中过夜培养;挑取单个阳性菌落按上述条件进一步培养,再对培养后菌落中的质粒进行测序.利用质粒提取试剂盒提取质粒,用超微量核酸蛋白测定仪(eppendorf)测定质粒标准品的浓度.根据文献[20]计算拷贝数,将得到的质粒标准品用1×TE梯度稀释构建标准曲线.

表2 mcyD基因和PC-IGS基因qPCR反应引物
1.4 定量PCR反应体系
定量PCR反应体系为20μL,其中含有10μL SYBR GREEN MIX,2pmoL引物,1μL模板,超纯水补足20μL, PCR反应在Bio-rad CFX Connet中进行.-基因扩增程序如下:95℃预变性2min, 95 ℃变性15s,58℃退火30s,72℃延伸30s,40个循环.基因定量PCR反应退火温度为56℃,其它条件与PC-IGS相同.
1.5 水中及细胞中微囊藻毒素检测
采用0.22μm一次性真空滤膜过滤5mL水样,保留含藻滤膜进行水中微囊藻毒素含量测定.细胞中的毒素使用GF/C膜过滤藻细胞并调节浓度后测定.所有毒素检测样本均使用中科院水生生物研究所研制的微囊藻毒素ELISA试剂盒进行检测.
1.6 数据统计
数据统计在R语言环境下进行,相关性计算采用Pearson相关性函数计算.数据展示采用平均值±标准差(mean±SD)形式.
2 结果与分析
2.1 海河水体理化参数
测得采样期间水体段理化指标见表3,采样期间海河水体的平均温度为29.2℃,4号采样点水温最高为30.4℃.水体呈弱碱性,5号采样点pH值最高.溶解氧变化范围较广,为4.77~17.15mg/L.总氮最高值为4号点3.55mg/L,最低值为1号点1.04mg/L.总磷最高点为2号点0.1mg/L,最低值为4号点0.05mg/L.

表3 海河水体主要理化参数
2.2 定量PCR标准曲线
利用质粒DNA为模板建立基因和基因定量PCR标准曲线.从图2和图3可以看出,循环阈值q与基因组拷贝数的对数呈显著线性关系.定量PCR反应中,序列定量PCR标准曲线方程为= -3.482+ 38.79 ,(2=0.993),扩增效率为93.8%.-序列定量PCR标准曲线方程为= -3.41+ 39.97,(2=0.999),扩增效率为96.5%.溶解曲线结果显示和-的扩增产物解链峰值温度分别为82.49℃(变异系数CV=0.09%)和84.41℃(变异系数CV=0.26%)

图2 mcyD基因荧光定量PCR标准曲线
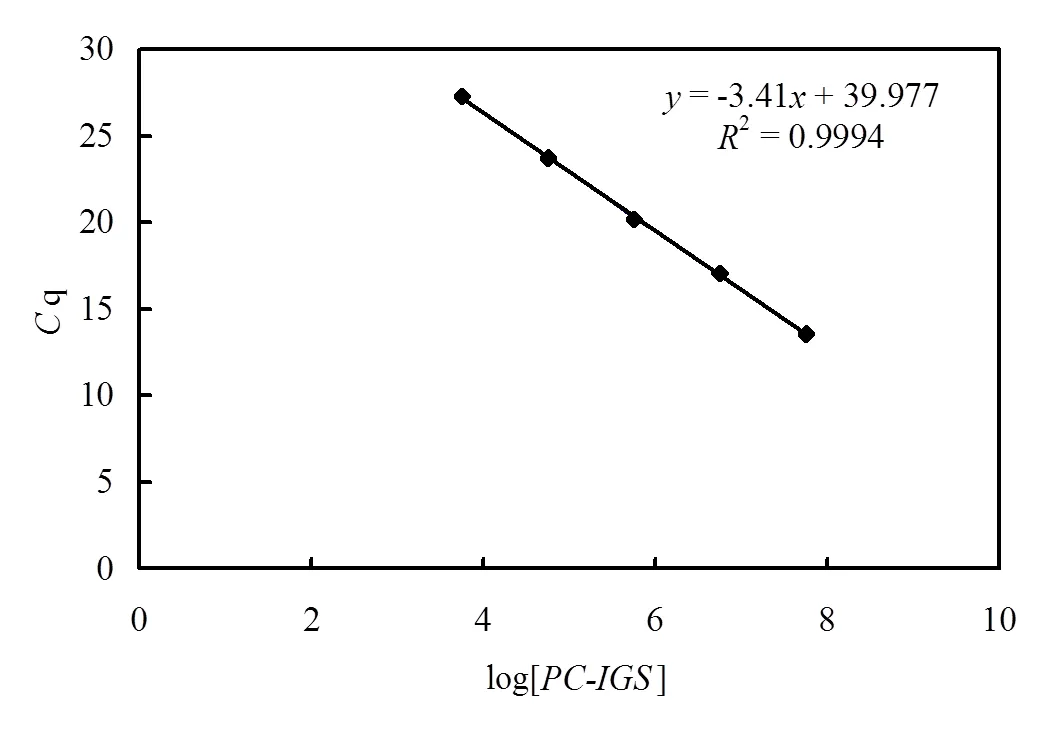
图3 PC-IGS基因荧光定量PCR标准曲线
2.3 产毒微囊藻和总微囊藻种群丰度的分布
定量PCR结果如图4所示,海河微囊藻种群丰度具有显著的空间分布差异.5号点天津之眼微囊藻种群丰度最高,每毫升水中微囊藻基因型拷贝数达到(1.53×108±1.13×107)copies/mL同时基因型拷贝数达到(2.48×107±7.12×106)copies/mL.3号点北洋园微囊藻种群丰度最低,微囊基因型拷贝数为(8.50×104±1.62×106) copies/mL,基因型拷贝数为(1.16×104±1.09×102)copies /mL.产毒微囊藻基因丰度与总微囊藻基因丰度比值为4.25%~28.59%平均比值为15.44%,各点之间差异较大.

图4 产毒微囊藻和总微囊藻种群丰度的空间分布
2.4 微囊藻细胞和水体中微囊藻毒素含量
ELISA酶联免疫反应检测结果如图5所示,藻细胞中毒素含量最高的点位为8号天津站为每升水中藻细胞含195.51μg毒素,最低3号为2.53μg.水中微囊藻毒素含量最高的也为8号天津站0.97μg/L.最低点3号0.18μg/L.4~8号点位显著高于1~3号点.通过线性回归得到细胞产毒和水中微囊藻毒素相关性2=0.95.
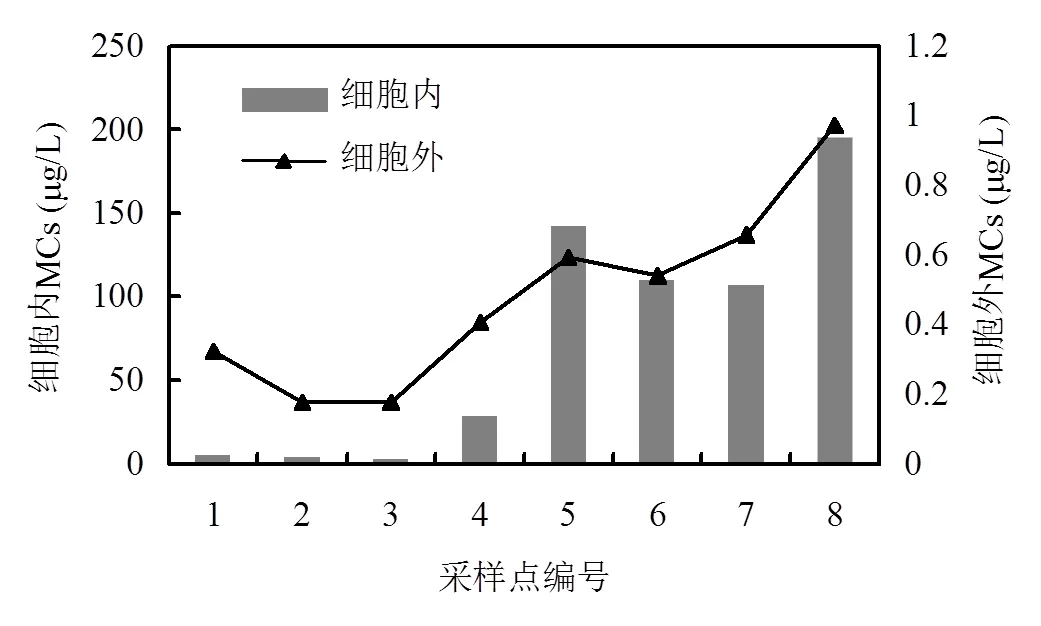
图5 每升水体中细胞内和细胞外的微囊藻细胞产毒素含量
3 讨论
3.1 基于荧光定量PCR结果的微囊藻基因丰度差异
近年来,夏季中的海河备受蓝藻困扰,蓝藻暴发对居民生活和城市景观带来较大影响.当前推测海河蓝藻暴发的原因有:海河生态用水不足,水体流动性差;海河水体中适合藻类生长的营养物质含量较高;天津夏季气候非常适应蓝藻生长等[21],但均缺乏定量的分析.此外对水体中微囊藻是否产毒,产毒微囊藻的丰富度如何也不甚清楚,缺乏相应的监测和报道.
近些年,荧光定量PCR在蓝藻水华的研究中得到广泛应用.这项技术能够对大批样品同时进行处理,对产毒微囊藻种群丰富度进行定量化的研究,快速对蓝藻水华是否产毒和毒性做出判断,可以预测蓝藻水华带来的生态风险[22].基因qPCR法具有灵敏性高、成本较低、所用时间短、所需检测样本少、操作简单等特点[23].qPCR法对于微囊藻产毒潜能的预测与毒素的直接检测结果具有高度一致性[24].研究中常选择单个基因作为目的基因进行qPCR扩增片段[25-30],也可选取多个基因作为目标基因进行扩增[31-33],本实验选取基因作为目的基因对水体中产毒微囊藻进行检测.-基因为特异性扩增微囊藻的基因片段,可检测出水体中总微囊藻基因丰富度.本实验通过建立基因和-基因的荧光定量PCR标准曲线,对海河水体中产毒微囊藻种群基因丰富度和总微囊藻种群基因丰富度进行定量分析.通过荧光定量PCR的结果表明,海河干流不同水系的微囊藻种群丰富度具有较大差异,北洋园(3号位点)产毒微囊藻丰富度与总微囊藻丰富度均最低;南运河(1号位点)和新开河(4号位点)产毒微囊藻丰富度与总微囊藻丰富度均较高,因此,海河干流中的微囊藻可能主要来源于南运河和新开河.
3.2 水体理化指标与微囊藻产毒基因及毒素的空间分布关系
通过对8月份海河天津市区段水体理化指标分析,可知此时的水体处于富营养化,再根据qPCR结果印证了在富营养化状态下微囊藻的总丰度较高[34].在自然环境中,许多种环境因子都可能影响产毒微囊藻占总微囊藻种群的比例,在Yoshida等[35]研究Mikata湖泊中产毒微囊藻和非产毒微囊藻种群动态随时间变化规律时,发现硝态氮浓度与产毒微囊藻占总微囊藻种群的比值呈显著正相关.这与本实验中南运河(爱民桥 1号位点)硝态氮浓度很高,产毒微囊藻占总微囊藻种群的比值也高的实验结论相符合.从研究结果可以看出,产毒微囊藻占总微囊藻的比值范围4.25%~28.59%,平均值为15.50%.其中比值最高的点为狮子林桥(6号位点),比值最低的点为进步桥(7号位点).相关报道表明不同湖泊产毒微囊藻占总微囊藻种群的比值的波动范围较大[36-37],狮子林桥与进步桥的距离较为相近,但产毒藻株比例却差异很大,导致这种结果的原因还需要进一步研究.根据刘永定等关于中国藻类学的研究表明高pH (大于8)值有利于原核藻类生长[38],海河水体8月pH值均在8以上,呈现弱碱性,非常有利于微囊藻生长,导致蓝藻暴发.6号位点pH值显示为9.01,7号位点为8.01,推测藻类丰富度可能与pH值有关.5号位点pH为9.3,是pH值最高的点,5号位点也是基因丰富度和-基因丰富度最高的点.产毒微囊藻基因丰富度与总微囊藻基因丰富度的比值最高的点为狮子林桥(6号位点),比值最低的点为进步桥(7号位点),其中6号位点的溶解氧9.11mg/L大于7号位点的溶解氧4.77mg/L.新开河(4号位点)溶解氧为最高17.5mg/L,产毒微囊藻基因丰富度也最高为2.08×106copies/mL,溶解氧最低的点为进步桥(7号位点),产毒微囊藻基因丰富度的最低点为北洋园(3号位点),但溶解氧最低点不是产毒微囊藻基因丰富度最低点.这与李大命等在巢湖夏季和冬季有毒微囊藻和无毒微囊藻种群丰度研究中的实验研究结果[39]出现了差异性,具体原因有待进一步研究.
3.3 水体理化指标与微囊藻产毒基因及毒素的相关性分析
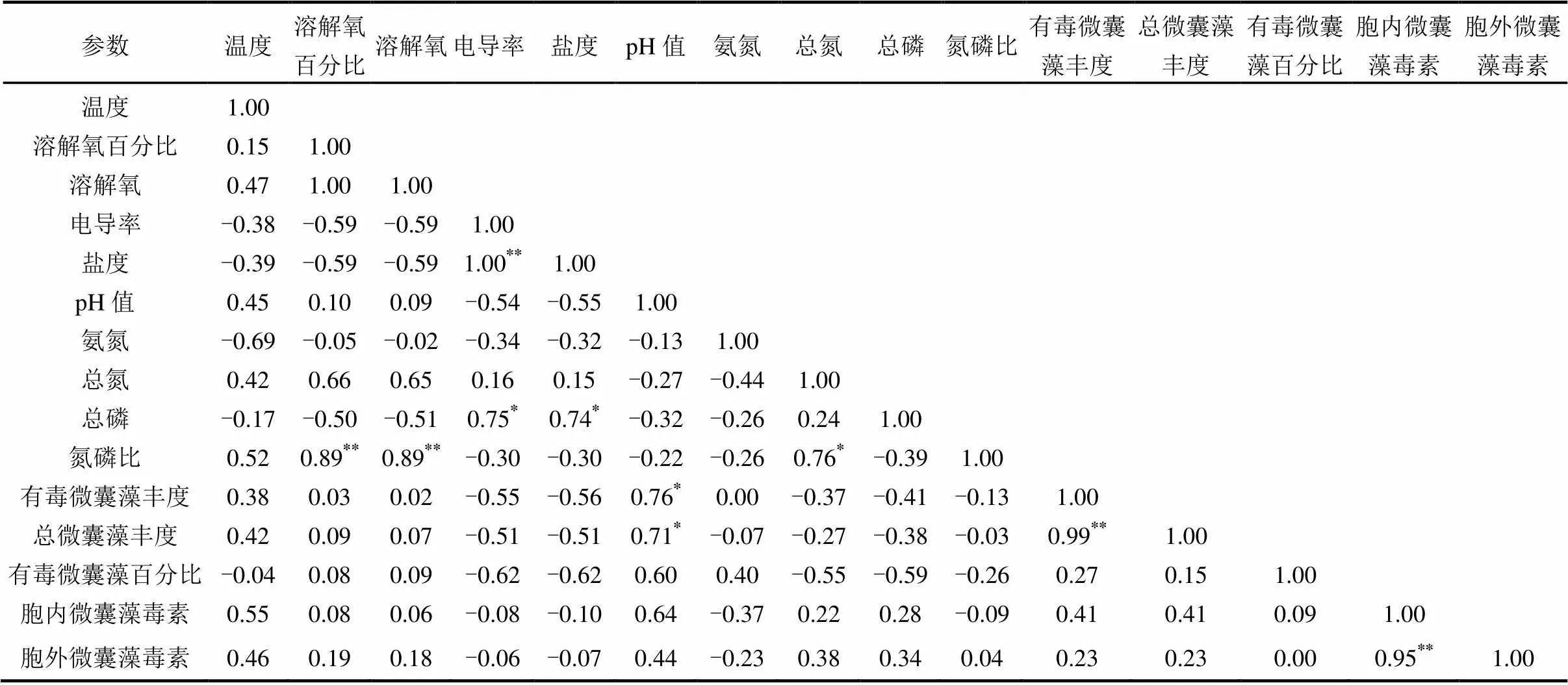
表4 环境因子与微囊藻和有毒微囊藻丰度之间的相关系数(n=8)
注: * :<0.05;**:<0.01.
已有研究表明,水体理化因子的差异可以显著的影响微囊藻毒素的释放.其中观点主要集中在温度、总氮、总磷、磷酸盐等[5,40-41]因素的影响,在本次研究中我们发现海河水体中微囊藻无论从总基因丰度到产毒基因丰度都与pH值显著相关(<0.05),这与本文在海河的另一项基于高通量测序得到的结果是一致的[42].这说明在高pH值情况下微囊藻无论在绝对丰度还是与其他原核生物共存的相对丰度水平上均能占据较大比例.在所有环境因子中,影响海河中微囊藻的丰度差异最重要的原因是pH值.然而目前的研究并不能给出pH值与微囊藻直接相关的明确原因,二者之间的因果关系尚不明晰.有一种推测是在微囊藻固碳过程中有一个中间形式为碳酸氢盐[43].蓝藻的集中暴发可能使碳酸氢根离子在水体中大量富集而后随着藻细胞衰败破裂而释放到水体中.微囊藻产毒基因、产毒藻株比例与各点位实际观测到的微囊藻毒素含量并没有显著相关性.这证明了虽然基因簇在微囊藻毒素合成中必不可少[44],但毒素的实际合成过程中受到转录因子调控或基因修饰影响显著,造成产毒藻株的丰度并不与毒素含量有实际关联.微囊藻毒素合成基因与微囊藻总16S rRNA基因丰度极显著相关(<0.01),这证明了在海河中微囊藻的产毒比例趋近一致,各采样点位产毒株所占比例大致相同.单位水体内胞外微囊藻毒素含量与胞内微囊藻毒素含量极显著相关,这说明在采样期间微囊藻毒素释放速率较为稳定,并没有发生大规模的胞内毒素释放事件.但值得注意的是,虽然藻细胞外水体中微囊藻毒素含量在各点位均未超过WHO规定的饮用水微囊藻毒素含量标准但是计算相同体积水体中藻细胞的毒素含量远超这一标准.这说明海河微囊藻具有较高的毒素释放潜力,这意味着当水体发生大规模藻细胞破裂事件时,水环境中毒素含量会远超过现有水平.一旦被饮用会对人体健康造成巨大威胁.在海河及与其类似的通海河道水体中,渗透压造成藻细胞破裂也可能会给河口生态系统造成巨大破坏.因此对于海河中微囊藻的控制是十分有必要的.
4 结论
4.1 夏季海河天津市区段微囊藻种群丰度具有明显空间差异产毒微囊藻种群丰度为1.16´104~2.48´107copies/mL,占总微囊藻种群的4.25%~28.59%.
4.2 藻毒素含量最高点为8号采样点天津站,每升水体中藻细胞共含毒素195.51μg,除去藻细胞水中毒素浓度为0.97μg/L.水体中微囊藻毒素未达到WHO规定的阈值.
4.3 产毒基因丰度及比例与环境中微囊藻毒素含量未呈现显著相关关系.pH值的大小与微囊藻总丰度和产毒基因型丰度呈现显著相关关系.
4.4 夏季海河中微囊藻胞内毒素含量远高于胞外含量,可能对下游的河口区域生态系统安全构成潜在威胁.
[1] Pouria S, Andrade A D, Barbosa J, et al. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. [J]. Lancet, 1998,352(9121): 21-26.
[2] Shun-Zhang Y U. Primary prevention of hepatocellular carcinoma [J]. Journal of Gastroenterology & Hepatology, 2013,95(4):427–434.
[3] Organization W H. Cyanobacterial toxins : Microcystin-LR [J]. Guidelines for Drinking-Water Quality, 1998,2:83-127.
[4] Kurmayer R, Kutzenberger T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp [J]. Applied & Environmental Microbiology, 2003,69(11):6723-6730.
[5] Davis T W, Berry D L, Boyer G L, et al. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms [J]. Harmful Algae, 2009,8(5):715-725.
[6] Joung S H, Oh H M, Ko S R, et al. Correlations between environmental factors and toxic and non-toxic Microcystis dynamics during bloom in Daechung Reservoir, Korea [J]. Harmful Algae, 2011,10(2):188-193.
[7] Dittmann E, Neilan B A, Erhard M, et al. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806 [J]. Molecular Microbiology, 1997,26(4):779-787.
[8] Tillett D, Dittmann E, Erhard M, et al. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: an integrated peptide-polyketide synthetase system. [J]. Chemistry & Biology, 2000,7(10):753-764.
[9] Baker J A, Neilan B A, Entsch B, et al. Identification of cyanobacteria and their toxigenicity in environmental samples by rapid molecular analysis.[J]. Environmental Toxicology, 2001,16(6):472-482.
[10] Hui P, Song L, Liu Y, et al. Detection of hepatotoxic Microcystis strains by PCR with intact cells from both culture and environmental samples. [J]. Archives of Microbiology, 2002,178(6):421-427.
[11] Ouahid Y, Pérez-Silva G, Campo F F D. Identification of potentially toxic environmental Microcystis by individual and multiple PCR amplification of specific microcystin synthetase gene regions [J]. Environmental Toxicology, 2005,20(3):235-242.
[12] Rinta-Kanto J, Ouellette A, Boyer G, et al. Quantification of toxic Microcystis spp during the 2003and 2004blooms in western lake Erie using Quantitative Real-Time PCR [J]. Environmental Science & Technology, 2005,39(11):4198-4205.
[13] 李大命,叶琳琳,于 洋,等.太湖水华期间有毒和无毒微囊藻种群丰度的动态变化[J]. 生态学报, 2012,32(22):7109-7116.
[14] 李大命,孔繁翔,张 民,等.太湖和巢湖夏季蓝藻水华期间产毒微囊藻和非产毒微囊藻种群丰度的空间分布[J]. 应用与环境生物学报, 2011(4):480-485.
[15] Li D, Kong F, Shi X, et al. Quantification of microcystin-producing and non-microcystin producing Microcystis populations during the 2009and 2010blooms in Lake Taihu using quantitative real-time PCR [J]. Journal of Environmental Sciences, 2012,24(2):284-90.
[16] Liu Y, Peiru H U, Sisan M A, et al. Detection of microcystin- producing Microcystis cells at the entrance of rivers to southern Lake Taihu by real-time fluorescence quantitative PCR [J]. Journal of Lake Sciences, 2016,28(2):246-252.
[17] Walls J T, Wyatt K H, Doll J C, et al. Hot and toxic: Temperature regulates microcystin release from cyanobacteria [J]. Science of the Total Environment, 2017,610-611:786.
[18] Mantzouki E, Lürling M, Fastner J, et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins [J]. Toxins, 2018,10(4):156.
[19] Kaebernick M, Neilan B A, Börner T, et al. Light and the Transcriptional Response of the Microcystin Biosynthesis Gene Cluster [J]. Applied & Environmental Microbiology, 2000,66(8):3387- 3392.
[20] Neilan B A, Jacobs D, Del D T, et al. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis [J]. Int. J. Syst. Bacteriol., 1997,47(3):693-697.
[21] 于洪利,杜庆有.天津海河2013年蓝藻暴发过程及相关因素分析[J]. 海河水利, 2017,s1:30-33.
[22] Baxa D V, Kurobe T, Ger K A, et al. Estimating the abundance of toxic Microcystis in the San Francisco Estuary using quantitative real-time PCR [J]. Harmful Algae, 2010,9(3):342-349.
[23] 谢数涛,龙思思,韩博平,等.微囊藻毒素合成酶基因的PCR检测方法 [J]. 热带亚热带植物学报, 2005,13(4):315-318.
[24] Fergusson K M, Saint C P. Molecular phylogeny of Anabaena circinalis and its identification in environmental samples by PCR. [J]. Applied & Environmental Microbiology, 2000,66(9):4145-4148.
[25] Baker J A, Entsch BNeilan B A, Mckay D B. Monitoring changing toxigenicity of a cyanobacterial bloom by molecular methods [J]. Applied & Environmental Microbiology, 2002,68(12):6070-6076.
[26] Tooming-Klunderud A, Mikalsen B, Kristensen T, et al. The mosaic structure of the mcyABC operon in Microcystis [J]. Microbiology, 2008,154(7):1886-1899.
[27] Tillett D, Parker D L, Neilan B A. Detection of toxigenicity by a probe for the microcystin synthetase A gene(mcyA) of the cyanobacterial genus Microcystis, comparison of toxicitieswith 16S rRNA and phycocyanin operon (phycocyanin intergenic spacer) phylogenies [J]. Applied & Environmental Microbiology, 2001,67(6):2810–2818.
[28] Bittencourt-Oliveira M D C. Detection of potential microcystin- producing in Brazilian reservoirs with a mcyB molecular marker [J]. Harmful Algae, 2003,2(1):51-60.
[29] Kurmayer R, Dittmann E, Fastner J, et al. Diversity of microcystin genes within a population of the toxic cyanobacterium Microcystis spp. in Lake Wannsee (Berlin, Germany). [J]. Microbial Ecology, 2002, 43(1):107-118.
[30] Jaana V, Anne R, Katrianna H, et al. Quantitative real-time PCR for determination of microcystin synthetase e copy numbers for microcystis and anabaena in lakes [J]. Applied & Environmental Microbiology, 2003,69(12):7289-97.
[31] Youness O, Gonzalo P S, Campo F F D. Identification of potentially toxic environmental Microcystis by individual and multiple PCR amplification of specific microcystin synthetase gene regions [J]. Environmental Toxicology, 2005,20(3):235-242.
[32] Oberholster P J, Myburgh J G, Govender D, et al. Identification of toxigenic Microcystis strains after incidents of wild animal mortalities in the Kruger National Park, South Africa [J]. Ecotoxicology & Environmental Safety, 2009,72(4):1177-1182.
[33] Hotto A, Satchwell M, Boyer G. Molecular characterization of potential microcystin-producing cyanobacteria in lake Ontario embayments and nearshore waters [J]. Applied & Environmental Microbiology, 2007,73(14):4570-4578.
[34] Ye C, Yang Y, Xu Q, et al. Volatile organic compound emissions from Microcystis aeruginosa under different phosphorus sources and concentrations [J]. Phycological Research, 2018,66:15-22.
[35] Yoshida M, Yoshida T, Takashima Y, et al. Dynamics of microcystin- producing and non-microcystin-producing Microcystis populations is correlated with nitrate concentration in a Japanese lake [J]. Fems. Microbiology Letters, 2007,266(1):49-53.
[36] Beversdorf L J, Chaston S D, Miller T R, et al. Microcystin mcyA and mcyE Gene Abundances Are Not Appropriate Indicators of Microcystin Concentrations in Lakes [J]. Plos. One, 2015,10(5): e0125353.
[37] Hotto A M, Satchwell M F, Berry D L, et al. Spatial and temporal diversity of microcystins and microcystin-producing genotypes in Oneida Lake, NY [J]. Harmful Algae, 2008,7(5):671-681.
[38] 刘永定,范 晓,胡征宇.中国藻类学研究 [M]. 武汉:武汉出版社, 2001:245.
[39] 李大命,孔繁翔,阳 振,等.巢湖夏季和冬季有毒微囊藻和无毒微囊藻种群丰度研究[J]. 环境科学学报, 2011,12:2672-2680.
[40] Vézie C, Rapala J, Vaitomaa J, et al. Effect of nitrogen and phosphorus on growth of toxic and nontoxic Microcystis strains and on intracellular microcystins concentrations [J]. Microbial Ecology, 2002, 43(4):443-454.
[41] Rintakanto J M, Konopko E A, Debruyn J M, et al. Lake Erie Microcystis: relationship between microcystin production, dynamics of genotypes and environmental parameters in a large lake [J]. Harmful Algae, 2009,8(5):665-673.
[42] Huo D, Chen Y X, Liu P, et al. Molecular detection of microbial communities associated with Microcystis vs Synechococcus dominated waters in Tianjin, China [J] Journal of oceanology and limnology, 2018,4(36):1145-1156.
[43] Sandrini G, Ji X, Verspagen J M, et al. Rapid adaptation of harmful cyanobacteria to rising CO2[J]. Proc. Natl. Acad. Sci., 2016,113(33): 9315-9320.
[44] Rantala A, Fewer D P, Hisbergues M, et al. Phylogenetic Evidence for the Early Evolution of Microcystin Synthesis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2004,101(2):568-73.
[45] 杨晓红,蒲朝文,张仁平,等.水体微囊藻毒素污染对人群的非致癌健康风险 [J]. 中国环境科学, 2013,33(1):181-185.
致谢:感谢天津农学院基础学院的刘萍老师在水化学分析中提供的帮助.
The spatial distribution ofand microcystin and its relationship with environmental factors in Haihe Tianjin City.
HUO Da1,2, CAO Qi1, WANG Su-yan1, CHEN Pei-pei1, LI Yi-ming1, QIAO Zhi-yi1,2*
(1.Department of Fishery Science, Tianjing Agricultural College, Tianjing 300384, China;2.Tianjin Key Laboratory of Aqua-ecology and Aqua-culture, Tianjin 300384, China)., 2018,38(10):3897~3903
The spatial distribution ofand microcystin in Haihe River during water bloom were investigated using quantitative real-time PCR (qPCR) and ELISA techniques. Microcystin synthetase gene () and-fragments were used as the target the total microcystin population and the potential microcystin-producing subpopulation. The results showed that microcystinexisted as a mixed population of potentially toxic and non-toxic genotypes in Haihe River. There was significantly spatial variation in the abundance of microcystin-producingThe abundance of microcystin-producingsubpopulation was ranged from 1.16×104to 2.48×107copies/mL and the ratio of its abundance to total microcystinvaried from 4.25% to 28.59%. The peakvalue of microcystinwas site point 8, which 1mL water contained 195.51μg microcystininalgae cells and 0.97μg/L in water. To sum up, total abundance ofremained a high level in Haihe Tianjin region. The toxin ofshowed various among different sampling sites. The most significant impact factors forabundance was pH value. However, the content of microcystin is not upon the value of WHO’s threshold. But the intercellular toxin in a given volume of water remains a high level, which may cause the potential risk to the ecosystem of down flow estuary.
;microcystin;quantitative real-time PCR;ELISA techniques;Haihe River
X522
A
1000-6923(2018)10-3897-07
霍 达(1993-),男,内蒙古呼伦贝尔人,天津农学院硕士研究生,主要从事分子生态学方向的研究.发表论文8篇.
2018-03-23
国家国际科技合作专项(2013DFA71340)
* 责任作者, 副教授, qiaozhiyi7070@163.com
猜你喜欢
中南药学(2022年8期)2022-11-19
海河水利(2021年6期)2021-12-28
考试与评价·高二版(2021年3期)2021-09-10
食品安全导刊(2021年21期)2021-08-30
建材发展导向(2021年23期)2021-03-08
江苏农业学报(2019年1期)2019-09-10
天然产物研究与开发(2018年8期)2018-09-10
中国环境科学(2017年5期)2017-05-23
中南大学学报(自然科学版)(2016年2期)2017-01-19
支点(2016年3期)2016-03-21
