
2种药用阿魏植物内生细菌及促生属性比较
2018-11-23 09:13刘永红郭建伟张永光祖丽皮亚木木沙尔OsamaAbdallaAbdelshafyMohamed董周焱古丽斯玛依艾拜都拉李文均
微生物学杂志 2018年5期
刘永红, 郭建伟, 张永光, 李 丽, 祖丽皮亚木·木沙尔,,Osama AbdallaAbdelshafyMohamed, 董周焱, 古丽斯玛依·艾拜都拉, 李文均,*
(1.中国科学院 新疆生态与地理研究所,新疆 乌鲁木齐 830011;2.中山大学 生命科学学院,广东 广州 510275;3.红河学院,云南 蒙自 661100;4.新疆大学 生命科学与技术学院,新疆 乌鲁木齐 830046;5.中国科学院大学,北京 100049)
新疆土地面积为166万平方公里,是我国面积最大的省级行政区,新疆的生物和矿产资源十分丰富,在国民经济发展中的地位相当重要。但新疆的绝大部分地区属温带大陆性干旱、半干旱气候,降水资源稀少、水分蒸发强烈,再加上风沙多,使得新疆的土地荒漠化、盐碱化严重,环境恶劣,许多普通植物不能在这种环境中生存。而植物内生菌具有固氮、溶磷、抗病和抗逆等功能,能够促进植物生长、提高抗逆性,产生活性次生代谢物等[1],在农业、医药等领域有着巨大的应用潜力[2]。内生菌可作为接种剂,改善植物在不良环境条件的生长,尤其是生长在盐碱和干旱等逆境胁迫下的植物,其相关细菌类群可能已经适应了逆境,并且能使宿主植物受益,很有必要对一些新疆特有植物及其内生菌开展相关研究。阿魏属植物在我国大约分布有 26 种 1 变种,其中分布于新疆的阿魏属植物有 20 种,9 种可作为药用,包括新疆阿魏、阜康阿魏、多伞阿魏和准噶尔阿魏等。阿魏在新疆少数民族传统民族药中的历史悠久,药用价值和经济价值高。如新疆阿魏的根和树脂能够消食健胃、通经止痛、祛风寒[3]。准噶尔阿魏可作为新疆阿魏的替代品使用,其药性与新疆阿魏相似,在哈萨克斯坦用于治疗头痛、感冒、胃病等[4]。近年来,放牧、过度采挖等人为因素造成阿魏生境的恶化,使得野生阿魏资源已越来越稀缺,有些甚至濒临灭绝。研究不同种阿魏和阿魏不同组织部位内生细菌的多样性,发掘具有特殊功能的内生细菌资源,并结合阿魏内生细菌的群落组成和物种分布,分析其内生细菌的优势种群,能够在一定程度上揭示阿魏的生存特点,探讨阿魏内生细菌的多样性形成机制,从而为内生细菌介导的阿魏生态适应性机制提供依据,对于从微生物角度研究干旱区植物抗病和抗逆等机制具有重要意义。
1 材料与方法
1.1 材料
1.1.1 样品采集 供试准噶尔阿魏样品于2015年6月采自新疆和布克赛尔县,新疆阿魏样品于2015年8月采自新疆石河子南山。基于不同生境采集两种阿魏的植株材料,每个采样点随机选择3株植物作为样本,每株植物间隔至少500 m。每个样点都采集其地上组织和地下根部,记录采样地点和植物生长地海拔高度(利用GPS),同时记录植物所处生境、植被类型等。

表1 阿魏样品采集信息Table 1 Description of the sampling sites
1.1.2 培养基 ①分离培养基(g/L):主要为以下9种[5],琼脂均为15。M1:丙酸钠2.0,L-天冬酰胺1.0,(NH4)2SO40.1,KCl 0.1,MgSO4·7H2O 30.0,FeSO4·7H2O 0.05;M2:柠檬酸0.12,柠檬酸铁0.12,NaCl 30.0,NaNO30.5,K2HPO40.4,MgSO4·7H2O 0.2,CaCl20.5;M3:Yeast 0.25,K2HPO40.5,NaCl 30.0;M4:琥珀酸钠 1.0,L-天冬酰胺1.0,KH2PO40.9,K2HPO40.6,NaCl 30.0,MgSO4·7H2O 0.1,CaCl20.2,KCl 0.3,FeSO4·7H2O 0.001;M5:微晶纤维素2.5,脯氨酸1.0,NaCl 30.0,KNO30.25,MgSO4·7H2O 0.2,K2HPO40.2,CaCl20.5,FeSO4·7H2O 0.01;M6:草酸钠2.0,酪蛋白水解氨基酸0.5,KH2PO40.3,Na2HPO4·12H2O 0.5,NaCl 30.0,ZnSO4·7H2O 0.02,CaCl20.5;M7:甘油10,L-天门冬酰胺1.0,NaCl 30.0,K2HPO41.0;M8:几丁质2.0,NaCl 30.0,K2HPO40.7,KH2PO40.3,MgSO4·7H2O 0.5,FeSO4·7H2O 0.1,ZnSO4·7H2O 0.001,MnCl20.001;M9:丙酸钠2.0,精氨酸1.0,NaCl 30,MgSO4·7H2O 1.0,KH2PO40.1,FeSO4·7H2O 0.05。②纯化及传代培养基(主要为ISP 2培养基):酵母提取物4 g, 葡萄糖4 g, 麦芽提取物10 g, 琼脂15 g,水1 L,pH 调至7.2,121 ℃ 灭菌 25 min。
1.2 方法
1.2.1 样品处理 将植物的地上和地下部分分开,用自来水将组织表面的泥土等冲洗干净,超声波清洗仪清洗数次,直至水质变清,再用吸水纸将组织上残留的水吸净。用75%的酒精浸泡植物根部和茎部1 min;将组织取出,用5%的次氯酸钠溶液浸泡8 min;将组织取出,无菌水漂洗5次;将组织取出,放入装有无菌滤纸的培养皿干燥2~3 d。最后一次冲洗植物组织的无菌水用内生细菌的分离培养基进行涂布培养,观察是否有菌落长出,以此验证表面消毒的效果。
1.2.2 内生细菌的分离、纯化及保藏 ①内生细菌的分离 取部分组织块以无菌搅拌器打碎,采用稀释涂布法分离阿魏内生细菌。无菌条件下,将1 g打碎后的植物组织样品与9 mL无菌水充分混合,配制倍数为10-1的植物组织悬浮液,然后依次稀释为10-2、10-3和10-4共3个梯度的组织悬浮液待用。最后用无菌枪头分别吸取40 μL各梯度下的组织悬浮液,涂布于添加有50 mg/L制霉菌素的9种分离培养基上,每个处理重复3次,30 ℃培养。②内生细菌的纯化及形态鉴定:将所有分离平板30 ℃培养3~4周,待细菌菌落长出后,挑取并接种于1/2浓度的ISP 2培养基上,用四区划线法进行纯化。然后根据菌落大小、形状、质地和生长情况及颜色、可溶性色素的产生情况等特征对分离纯化的内生细菌菌株进行并菌、归类。③内生细菌的菌株保藏:用无菌竹签挑取已纯化的内生细菌单菌落,加入装有1.8 mL 20%无菌甘油的甘油管中,再用振荡器将甘油和菌体进行混合,于-80 ℃冰箱中保存备用。
1.2.3 内生细菌的系统发育分析 ①基因组DNA 提取:在装有菌体的1.5 mL EP管中加入480 μL 1×TE缓冲液和20 μL 2 mg/mL的溶菌酶,于37 ℃、160 r/min摇床培养 1~2 h,加入50 μL 20% SDS(sodium dodecyl sulfatesodium salt) 和5 μL 20 μg/mL蛋白酶震荡1 min,放于55 ℃烘箱保温60 min;加入V苯酚∶V氯仿∶V异戊醇=25∶24∶1溶液,震荡混匀,12 000 r/min离心10 min,取上清,转管(重复抽提2次);上清中加入80 μL 3 mol/L乙酸钠(pH 4.8~5.2)混匀后加800 μL无水乙醇,室温放置10 min以上;于12 000 r/min低温离心20 min,弃上清,加200 μL 70%乙醇洗1~2次,12 000 r/min离心10 min,弃乙醇,在37~55 ℃条件下干燥,然后加入30~50 μL的1×TE溶解DNA,-20 ℃保存。②菌株鉴定:采用上海生工生物公司合成的16S rRNA基因通用引物27F(5′-CAGAGTTTGATCCTGGCT-3′)和1492R(5′-AGGAGGTGATCCAGCCGCA-3′),以50 μL的PCR反应体系进行扩增。加入Reaction Buffer(10×): 5.0 μL,2×TaqPCR Master Mix: 4.0 μL,引物27F: 1.0 μL,引物1492R: 1.0 μL,模板DNA: 2.0 μL,补充水至50.0 μL。然后94 ℃预变性4 min,94 ℃变性45 s,55 ℃退火45 s,72 ℃延伸1 min,72 ℃最终延伸5 min,循环32次。最终取PCR产物3 μL与6×loading buffer混合点样,采用1%琼脂糖凝胶电泳检测,染色剂为Golden view EB染料,缓冲液为1×TAE,电压115 V,时间23 min。最后用凝胶成像系统检测扩增结果,检测出目的条带的样品送往上海生工生物公司进行测序。测序结果首先在EzTaxon中进行序列比对,选择和下载近缘种序列,用MEGA 6软件Neighbour-Joining 法构建系统进化树,再结合16S rRNA基因序列同源性大于98%为同一种[6]的原则确定菌株分类地位。
1.2.4 内生细菌群落多样性及促生属性分析 采用优势度指数(Dominance)、香农指数(Shannon)、均匀度指数(Evenness)、丰富度指数(Margalef)和辛普森指数(Simpson)5个指标评价内生细菌群落的多样性,具体分析方法参考文献[7]所述,利用PAST软件(Version2.03)进行整理和分析。内生细菌的固氮、溶磷、产吲哚乙酸和产铁载体等促生属性的检测方法参考Liu等[8]已发表的内容进行。
2 结果与分析
2.1 表面消毒结果检测
采用印迹法对表面消毒效果的验证显示,平板上无菌落长出,因此证明所有的表面消毒程序有效,植物表面消毒彻底,后续分离得到的菌株确实是来自于植物组织内部。
2.2 最佳分离培养基探索
为探索阿魏内生细菌分离的最佳培养基,对每种培养基不同浓度中分离到的内生细菌种类和数量进行统计。由图1可知,M5和M7培养基各分离到13个属的内生细菌,其次为M6和M1培养基,分别分离到11个属和10个属的内生细菌。由此可以初步判断,M5、M7、M6和M1四种培养基为较理想的阿魏内生细菌分离培养基。从所得菌株数量来看,M2和M7培养基所分离到的内生细菌较多,M2培养基分离到29株内生细菌,M7培养基分离到25株内生细菌;其次为M1和M5培养基,各分离到21株内生细菌。因此,综合考虑每种分离培养基所分离的内生细菌种类和数量,M7为分离阿魏内生细菌实验的最佳培养基,M2和M5培养基也较适于分离阿魏内生细菌。
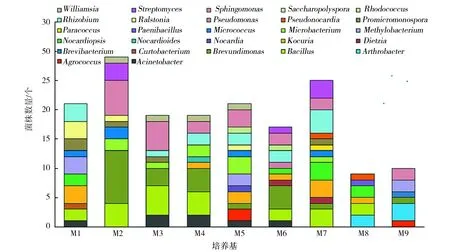
图1 不同培养基分离效果比较Fig.1 Comparison of endophytic bacteria isolated from different media
2.3 2种植物的内生细菌组成和多样性比较
2.3.1 2种阿魏内生细菌组成 由表2可知,新疆阿魏植株样品共获得125株内生细菌,分属于3个门、13个目、23个科、 29个属,其优势度指数为0.07、香农指数为2.91、均匀度指数为0.63、丰富度指数为5.81、辛普森指数为0.93;准噶尔阿魏植物样品共分离获得170株内生细菌,分属于3个门、14个目、20个科、 27个属,其优势度指数为0.08、香农指数为2.83、均匀度指数为0.63、丰富度指数为5.06、辛普森指数为0.92。可以看出,2种阿魏植物内生细菌的优势度和均匀度无明显差异,但新疆阿魏内生细菌群落的多样性指数和丰富度指数更高。
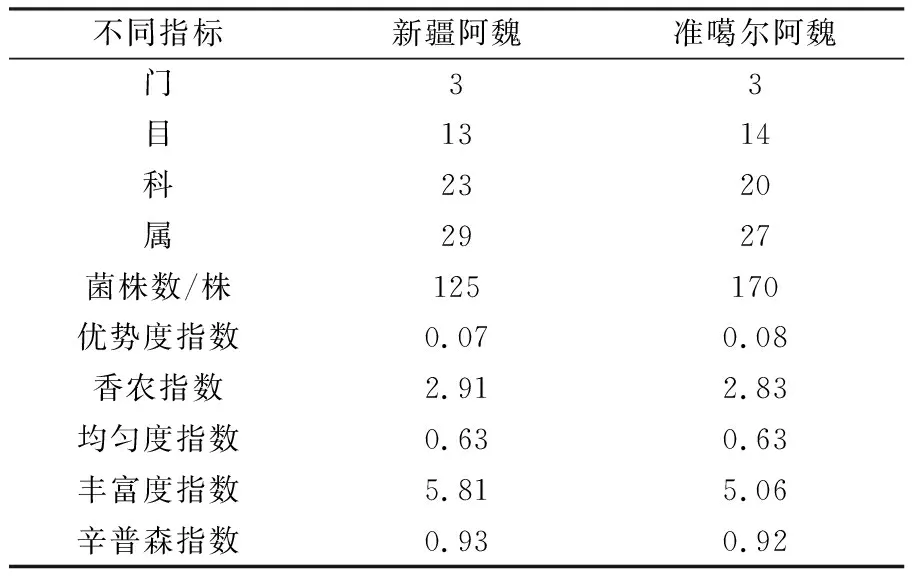
表2 2种阿魏内生细菌的物种组成和多样性比较Table 2 Comparison of endophytes diversity and composition from two kinds of Ferula
2.3.2 2种阿魏内生细菌优势种比较 图2和图3从物种的属水平分别展示了新疆阿魏和准噶尔阿魏内生细菌的优势菌。可以看出,新疆阿魏内生细菌中芽胞杆菌属(Bacillus)为优势菌,占分离菌株总数的5.6%,其次为链霉菌属(Streptomyces)、微杆菌属(Microbacterium)和节杆菌属(Arthrobacter),分别占新疆阿魏内生细菌的4.8%、4.0%和4.0%。此外,新疆阿魏内生细菌还存在许多稀有种,例如,包括类芽胞杆菌(Paenibacillus)、紫杆菌属(Porphyrobacter)、副球菌属(Paracoccus)、德沃斯氏菌属(Devosia)等在内的17个属,仅占分离总数的0.8%;准噶尔阿魏内生细菌的优势菌也是芽胞杆菌属(Bacillus)和链霉菌属(Streptomyces),二者分别占分离菌株总数的15.51%和8.62%,其次为节杆菌属(Arthrobacter)和鞘氨醇单胞菌属(Sphingomonas),分别占分离总数的6.9%。可以看出,2种阿魏植物内生细菌群落的优势菌具有很高相似性,芽胞杆菌属(Ba-cillus)、链霉菌属(Streptomyces)和节杆菌属(Arthrobacter)在这两种植物中都占有极其重要的生态位。通过比较还可以看出,2种阿魏的内生细菌群落具有差异性,如Porphyrobacter,Amycolatopsis、Bradyrhizobium等只在新疆阿魏中分布,而Saccharopolyspora只在准噶尔阿魏中分布。
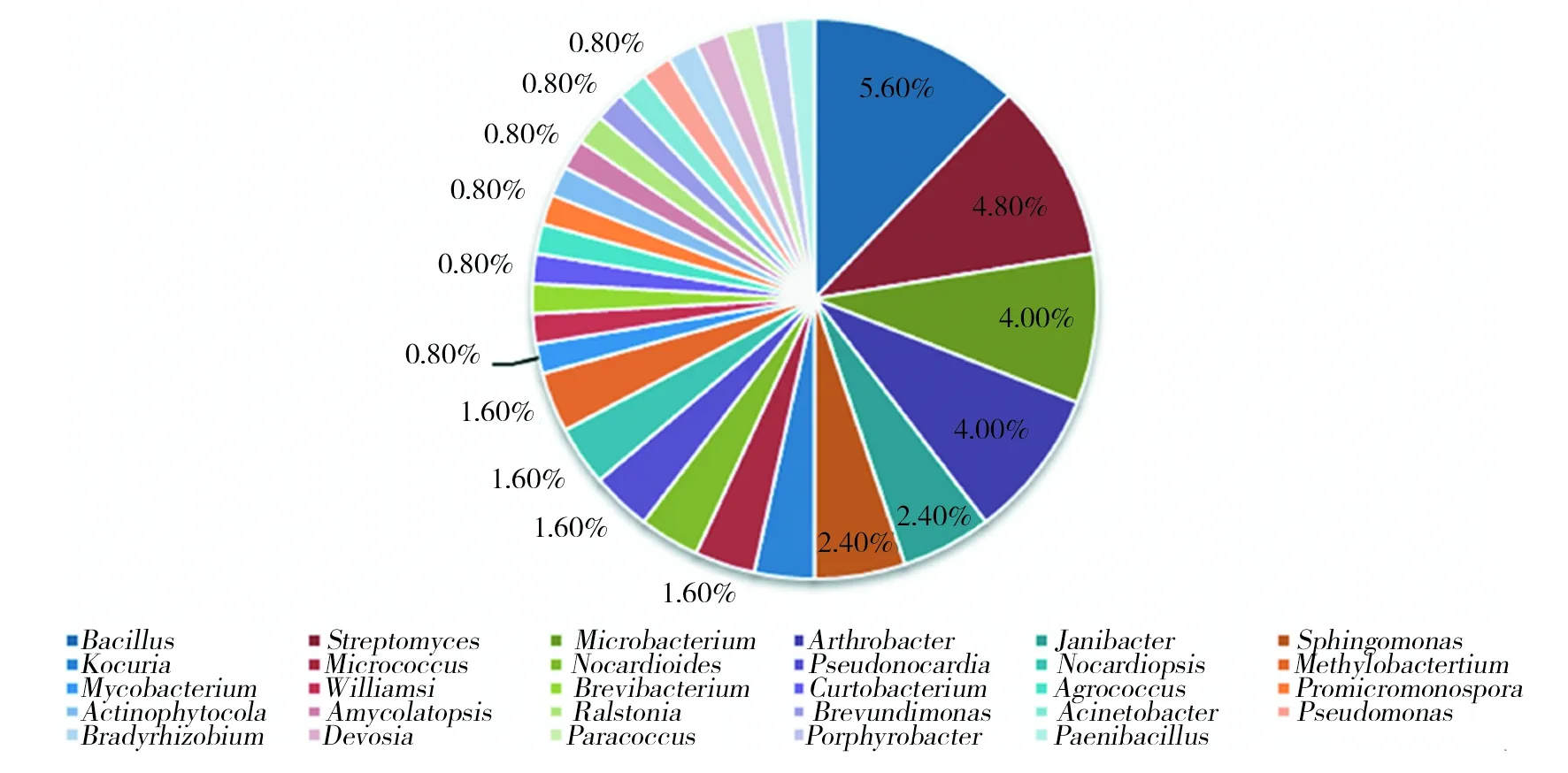
图2 新疆阿魏内生细菌不同属的分布情况Fig.2 Distribution of endophytes from F. sinkiangensis at genus level
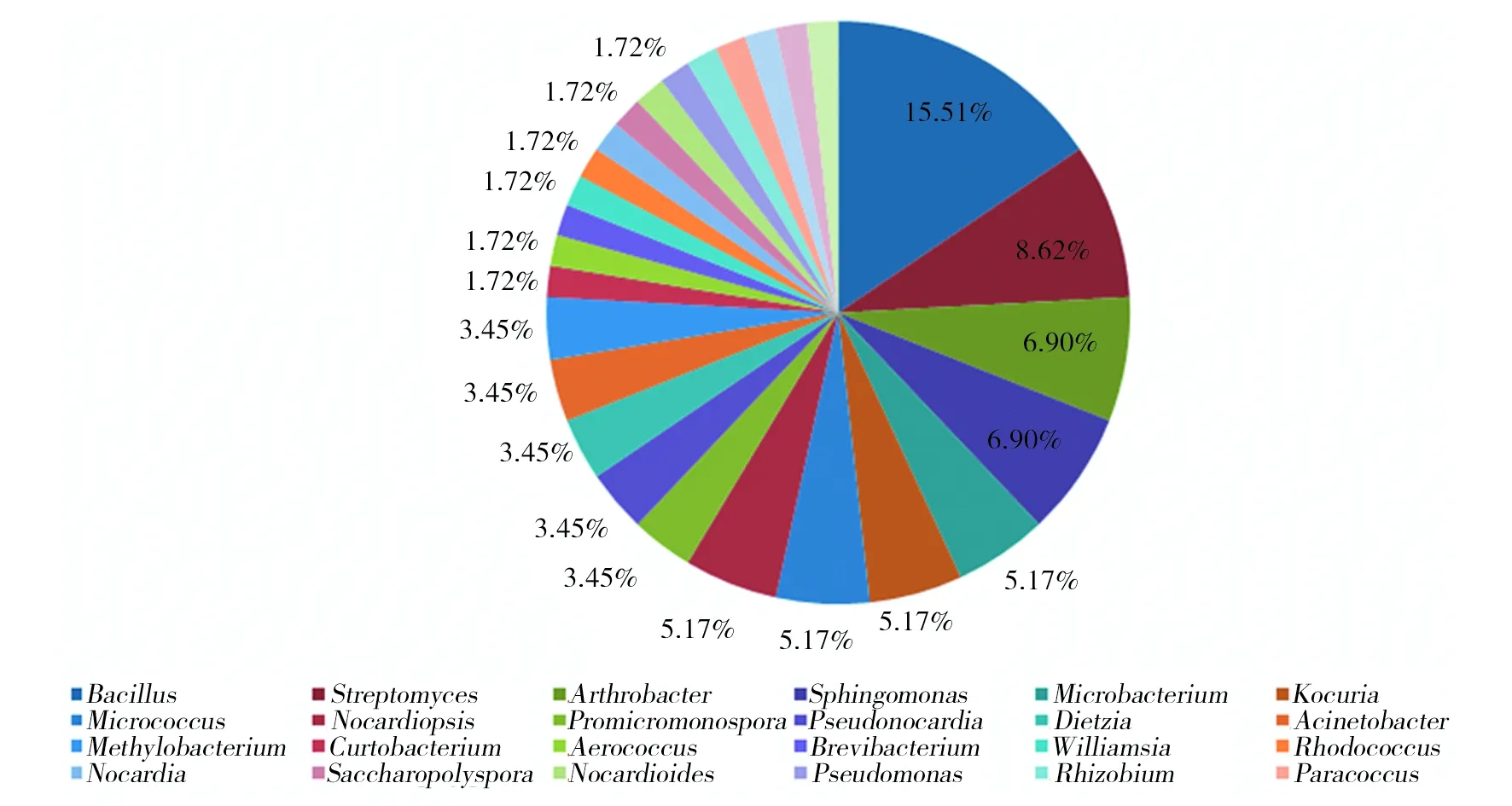
图3 准噶尔阿魏内生细菌不同属的分布情况Fig.3 Distribution of endophytes from F. songorica at genus level
2.4 不同组织部位植物内生细菌的分布
表3通过多样性指数比较了2种阿魏不同组织部位内生细菌的多样性。结果显示,新疆阿魏地上组织中的55株内生细菌被分为13个分类单元,其优势度指数为0.14,香农指数为2.25,丰富度指数为3.10;地下组织根部的70株内生细菌被分为27个分类单元,其优势度指数为0.06,香农指数为3.07,丰富度指数为6.23。这些结果表明,新疆阿魏根部内生细菌群落具有明显的多样性,但其地上组织中优势菌的优势度更明显;准噶尔阿魏地上组织中的82株内生细菌被分为21个分类单元,其优势度指数为0.08,香农指数为2.70,丰富度指数为4.54;地下组织根部的88株内生细菌被分为22个分类单元,其优势度指数为0.09,香农指数为2.68,丰富度指数为4.69。可以看出,准噶尔阿魏内生细菌群落中,其地上组织和地下组织的优势度、丰富度和均匀度都无明显差异,多样性和优势菌的优势度也不明显。
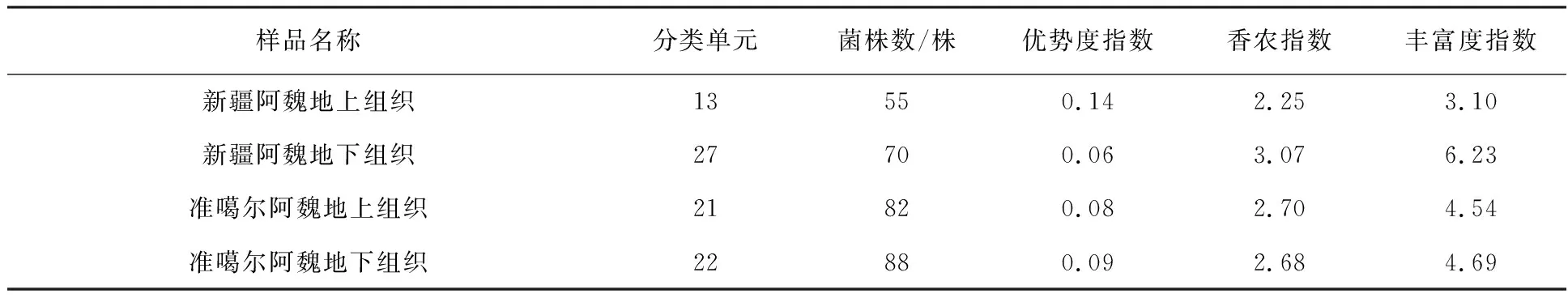
表3 2种阿魏不同组织部位内生细菌的多样性Table 3 Endophytic diversity from different tissues
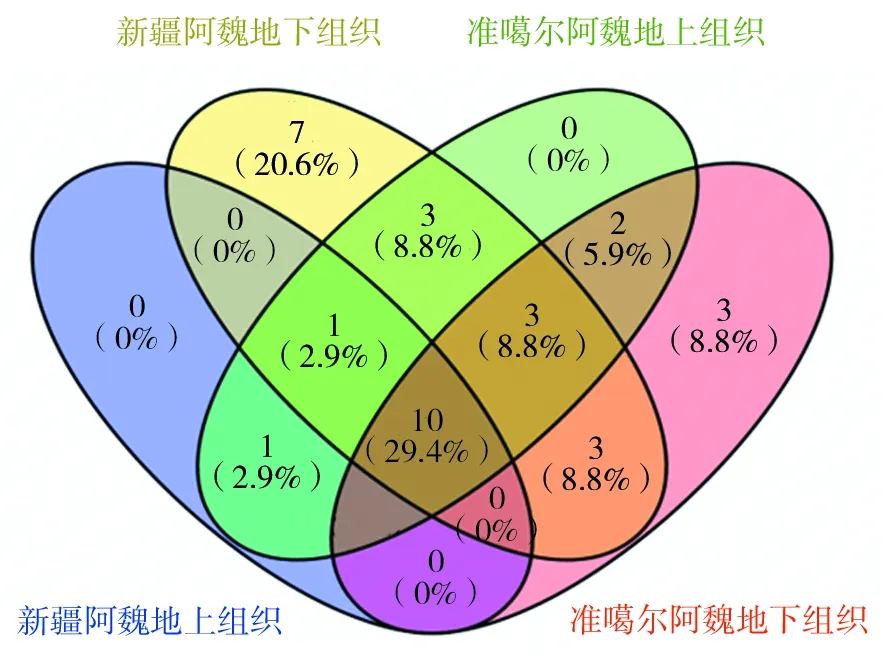
图4 2种阿魏不同组织部位内生细菌的韦恩图分布Fig.4 Venn distribution of endophytes from different tissues
如图4所示,通过韦恩图分布进一步比较2种阿魏不同组织部位内生细菌的分布情况。新疆阿魏内生细菌的29个属中,有2个属的内生细菌仅存在于地上组织,有16个属的内生细菌仅存在于地下组织,另外11个属的内生细菌在地上组织和地下组织中都有分布;准噶尔阿魏内生细菌的27个属中,有5个属的内生细菌仅分布于地上组织,有6个属的内生细菌只分布于地下组织,还有16个属的内生细菌在地上组织和地下组织中都有分布。通过比较2种阿魏不同组织部位的内生细菌发现,有13个属的内生细菌同时存在于新疆阿魏和准噶尔阿魏的地上组织,16个属的内生细菌同时存在于2种阿魏的根部组织。另外,还有10个属的内生细菌在新疆阿魏和准噶尔阿魏的地上和地下组织中同时存在。由此可以看出,2种阿魏的内生细菌在物种组成和种类分布上具有很高的相似性。
2.5 阿魏植物内生细菌的潜在新物种
表4为2种阿魏植物内生细菌的潜在新物种。可以看出,新疆阿魏(F.sinkiangensis)中总共有5株菌可能为内生细菌潜在新种,这5株菌分布在Porphyrobacter、Paracoccus、Amycolatopsis、Actinophytocola和Devosia。其中,菌株SX2RGS8从16S rRNA基因序列和系统发育分析结果可初步判定为潜在新属;准噶尔阿魏(F.songorica)中只发现1株Nocardioides潜在新种。值得一提的是,这6株潜在新内生细菌都来自植物的根部组织。
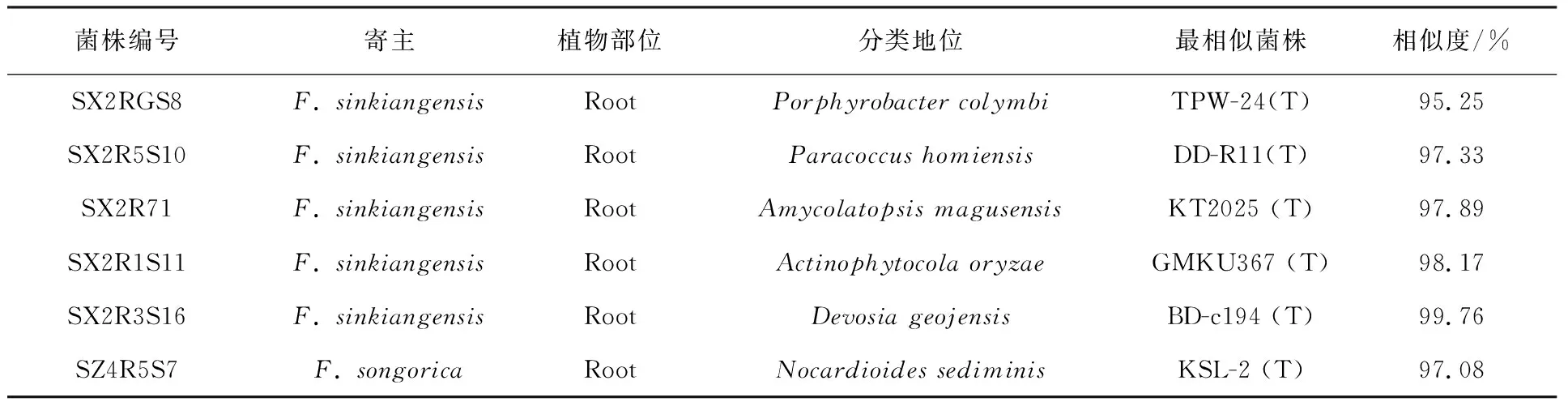
表4 阿魏植物内生细菌的潜在新分类单元Table 4 Potential novel species
2.6 阿魏植物内生细菌的促生属性
2.6.1 新疆阿魏内生细菌促生属性 如表5所示,新疆阿魏内生细菌中具有溶磷属性的菌株较少,且只分布在Microbacterium、Acinetobacter和Ralstonia这3个属,其中Microbacterium总共检测29株菌,但只有3株菌具有溶磷活性;Acinetobacter总共检测4株菌,且这4株菌都具有溶磷活性。相比之下,具有固氮潜力的菌株较多,共有9个属的内生细菌检测到该活性,其中Microbacterium的29株被检测菌株,有24株具有固氮潜力。另外,还有14个属的内生细菌具有产吲哚乙酸能力,不过产吲哚乙酸的菌株大多(23株)分布在Microbacterium,本研究中被检测的Bacillus菌株都具有产吲哚乙酸能力。产铁载体能力表明,共有11个属的内生细菌具有产铁载体能力,且在Microbacterium中分布最多,共检测到17株。
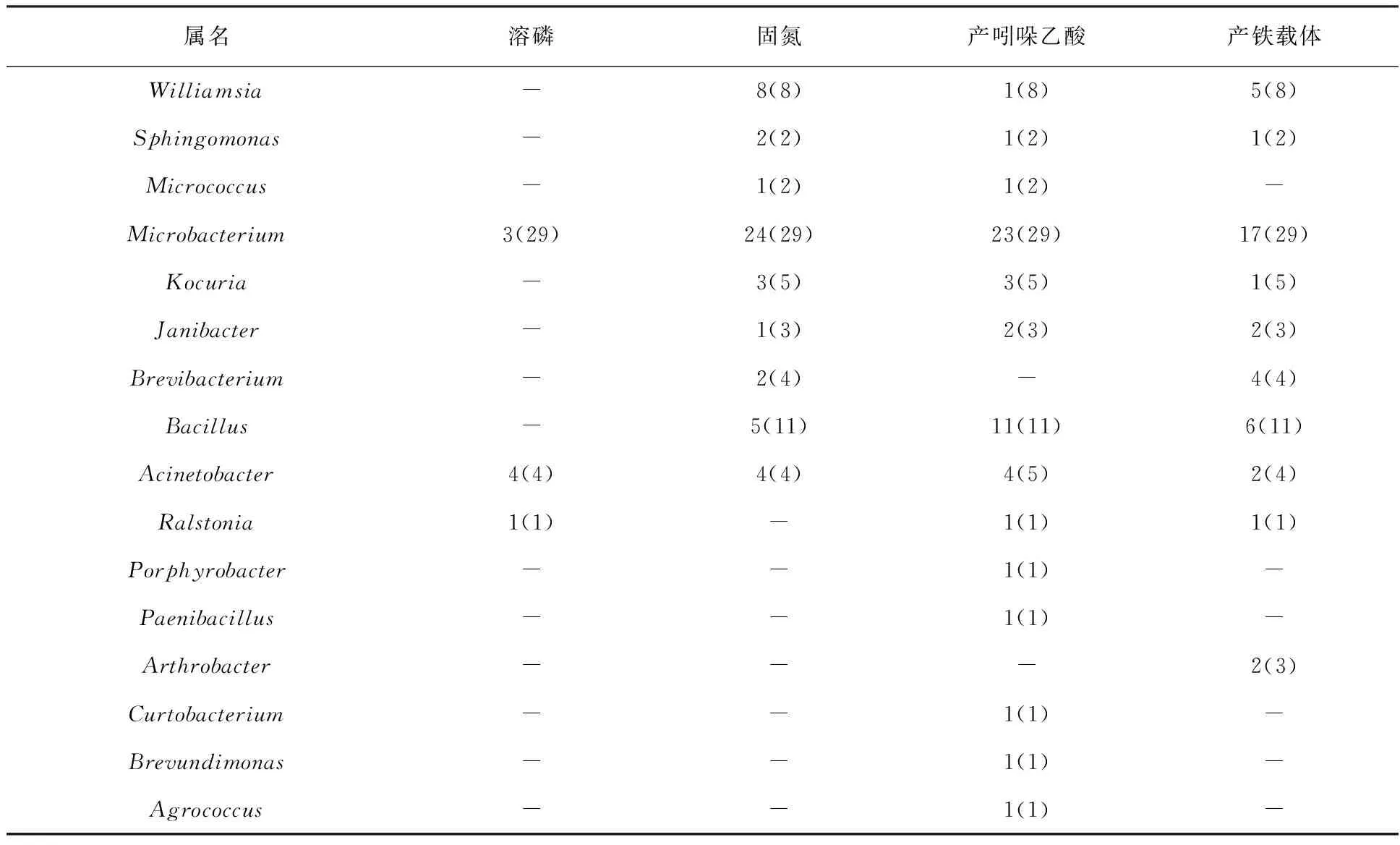
表5 新疆阿魏内生细菌的促生属性检测结果Table 5 Positive strains tested for growth promotion traits associated with F. sinkiangensis
2.6.2 准噶尔阿魏内生细菌促生属性 如表6所示,准噶尔阿魏内生细菌中共有7个属的内生细菌表现出溶磷潜力,17个属的内生细菌具有固氮能力。具有产蛋白酶能力和产纤维素酶能力的内生细菌分别为8个属和9个属。同样可以看出,具有溶磷潜力的菌株较少,具有固氮潜力的菌株最多。另外,Bacillus、Curtobacterium和Microbacterium中的一些菌株同时具有溶磷、固氮、产蛋白酶和产纤维素酶4种能力。
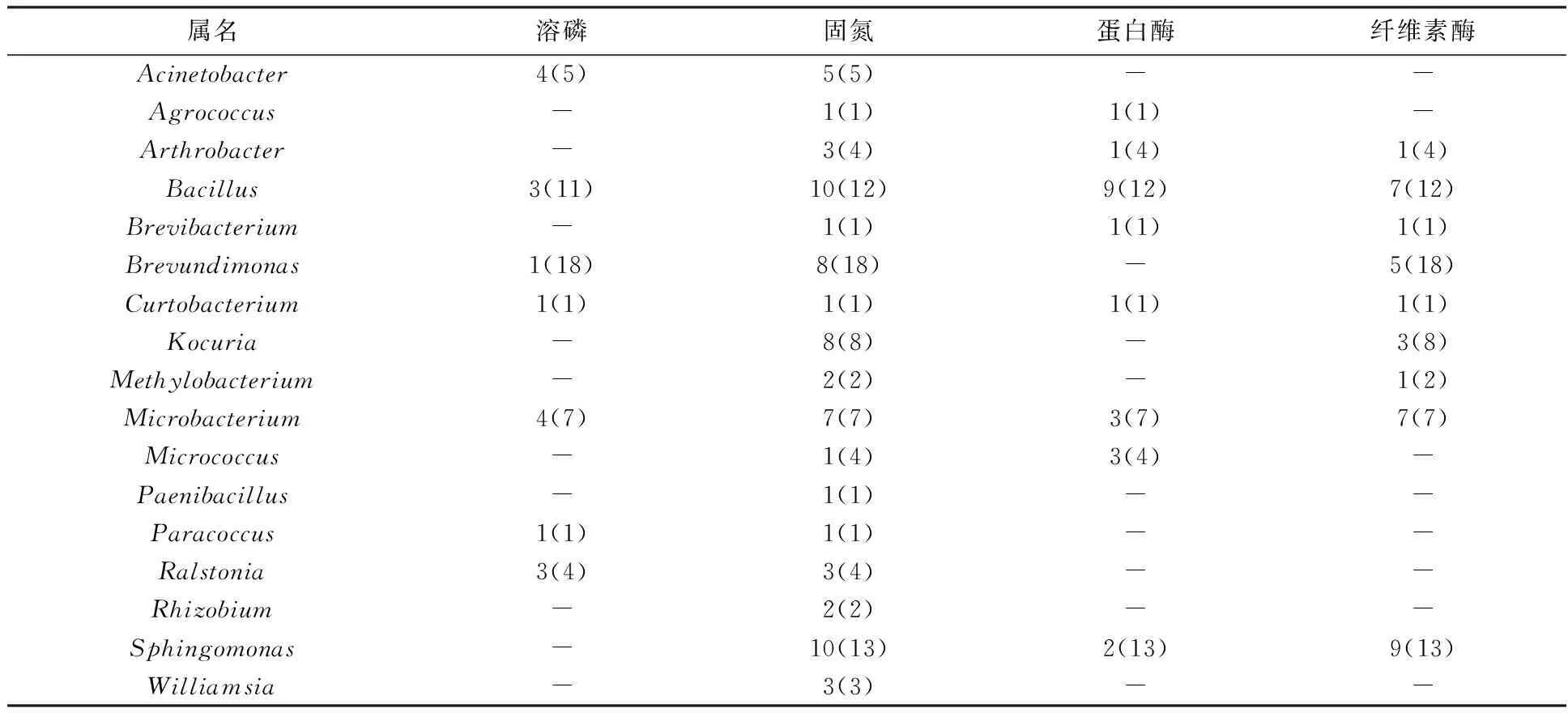
表6 准噶尔阿魏内生细菌的促生属性检测结果Table 6 Positive strains tested for growth promotion traits associated with F. songorica
3 讨 论
内生细菌是植物微生态系统中的重要组成部分,在植物中的分布具有普遍性。研究表明,每一种植物体内都至少有一至多种不同的内生菌[9],植物根、茎、叶、果实、种子、块茎、胚珠及豆科根瘤中都有内生菌存在[10]。本研究以2种新疆特有的阿魏植物为研究对象和材料,分别从其地上组织茎部和地下组织根部分离内生细菌。通过比较发现,2种阿魏植物内生细菌群落组成具有差异性,但更多的表现出相似性。差异性表现在Saccharopolyspora只在准噶尔阿魏中有所分布,Mycobacterium、Janibacter和Promicromonospora等只在新疆阿魏中分布;相似性:①两种阿魏内生细菌的优势菌均为芽胞杆菌属(Bacillus)和链霉菌属(Streptomyces);②两种阿魏根部的内生细菌多样性都高于茎部;③有10个属的内生细菌在新疆阿魏和准噶尔阿魏的地上和地下组织中同时存在。
关于内生细菌多样性形成的影响因素已有诸多报道。植物内生细菌的分布除受时间和空间变化影响外,也受植物组织部位的影响[11-12],根部内生细菌多样性较高是因为根部环境相对较稳定[13],其内生细菌群落受到的影响也相对较少。王宏飞[14]在研究不同样地中盐生植物内生菌的多样性变化时发现,土壤性质是影响植物内生菌群落组成的主要因子之一,且随着土壤盐浓度的增加,内生菌群落多样性降低;另外,内生菌群落的优势菌也会随着土壤盐浓度的变化而变化,如当土壤盐浓度增加时,链霉菌属的数量明显减少,相反,芽胞杆菌属的数量却呈上升趋势。因此可以推断,本研究中土壤性质在一定程度上造成了2种阿魏内生细菌群落的差异性,但由于寄主植物都是阿魏且药用成分相似[15],使得这2种植物内生细菌群落更多地表现出了相似性分布特点。
植物内生菌在改良植物生长、提高植物适合度、增强植物生物和非生物胁迫抗性上具有重要作用[16-17]。尤其在极端环境条件下,内生菌能够改变渗透调节、控制离子吸收、增加气孔导度[18]等减轻环境胁迫对植株造成的不利影响,提高植物生长能力[19]。本研究所分离到的新疆阿魏和准噶尔阿魏内生菌资源中大部分具有固氮、产生长素和产铁载体等能力。具有固氮能力的内生菌能够帮助植物固氮以提供生长所需的氮素营养,甚至可以产生生长素等生物调节物质,从而促进植物的生长[20]。产铁载体能力表明内生菌可以为生长在铁胁迫条件下与病原菌竞争铁源,从而抑制病原微生物的生长以促进宿主植物根系的抗病能力[21-22]。有研究表明,芽胞杆菌属和链霉菌属是许多植物内生菌的优势菌株,且这2个属的菌株具有生防和促生功能[23-24]。本研究进一步证实芽胞杆菌属的菌株具有固氮、溶磷、产铁载体、产吲哚乙酸、产蛋白酶及纤维素酶等多种促生属性,一些芽胞杆菌同时具有多种促生活性,这些研究结果对于促进植物生长和病虫害的生物防治具有重要意义[25]。
总之,本研究中分离到的植物促生菌及内生细菌新资源为进一步研究荒漠地区盐生植物的生态适应性提供了一定的理论基础,同时为微生物的资源收集和保育工作带来了更多的可能。通过进一步研究2种阿魏植物内生细菌的相似性分布特点和形成机制,对于如何利用内生细菌与植物的共生特性对植物资源进行开发和保护,以及如何依赖内生细菌进行药用植物珍稀活性成分的生产等极其重要。
猜你喜欢
食品安全导刊(2021年20期)2021-08-30
书屋(2020年9期)2020-09-23
湖北农机化(2020年4期)2020-07-24
卫拉特研究(2020年0期)2020-01-19
世界农药(2019年4期)2019-12-30
今日农业(2019年11期)2019-08-15
蒙古学问题与争论(2019年0期)2019-03-29
乡村地理(2018年2期)2018-09-19
新疆农垦科技(2018年5期)2018-07-09
新疆农业科技(2014年6期)2014-03-03
