
低温下外源腐胺对黑皮果蔗拔地拉光合性能的影响
2018-11-30 06:43朱俊杰王天顺闫飞燕
西南农业学报 2018年10期
朱俊杰,王天顺,牙 禹,闫飞燕
(1.广西大学,广西 南宁 530004;2. 广西农业科学院农产品质量安全与检测技术研究所,广西 南宁 530007)
【研究意义】黑皮果蔗拔地拉 (SaccharumofficinarumL.cv. Badila)居目前我国果蔗种植面积的首位[1],由于是热带地区起源的原种,缺乏抗冷的遗传基础,引进到我国几十年来其抗冷性一直没有根本性的改善,严重影响其产量和品质。过去果蔗生产中曾广泛采用化学调控手段增加产量和改善其品质[2],但近年来人们对合成类的生长调节物质的使用有所顾忌和排斥。开发拔地拉内生调节物质显得尤为重要,不但可为果蔗化学调控提供新手段,还可为果蔗抗冷育种和抗寒栽培措施的制定供理论参考。【前人研究进展】多胺是生物体中一类低分子脂肪族含氮碱,主要有腐胺(Put)、亚精胺 (Spd)、精胺(Spm)、尸胺(Cad)及鲜精胺(Agm)等,具有阳离子性质[3],能够和核酸、多肽及蛋白质等带负电荷的大分子结合,对维持细胞酸碱平衡,稳定细胞结构起重要作用[4]。同时,多胺本身具有较高的生物活性,是植物内生的一类生长调节物质,具有参与植物信号转导[5],调控细胞分裂、生长、分化及衰老死亡[6],增强细胞对外界逆境胁迫的抗性等功效[7],长期以来都是植物生理、植物营养等学科领域内重要和活跃的研究领域[8]。C4植物通常比C3植物有更强的光合同化能力,在高温和缺水环境中更是如此,但在低温条件下,后者的光合优势更明显。因此,如何增强C4植物的低温光合耐受力长期以来一直是植物光合生理生态研究的重要问题。近二十多年来,随着光合气体交换和叶绿素荧光等技术的突破性进展,C4植物低温光合生理生态机制获得一些新认识:低温下C4植物量子效率明显低于C3植物[9],加之C4光合酶对低温敏感[10],更易遭受低温光抑制[11-12],整体光合效率低于C3植物;C4植物低温光合表现种间差异大,耐受机制也有差异[13],需结合特定环境开展相应的研究。如前所述,多胺具有活性高、功能多、成本低等优势,是植物内生调节物质选择时优先考虑对象之一。【本研究切入点】多胺对植物抗寒(冷)性的作用效果因植物种类的不同表现出明显差异,例如多胺可以增强一些植物的抗冷能力[14-15],但对部分植物却没有效果[16]。生产实践中使用的多胺通常包括腐胺、精胺和亚精胺,植物体经代谢反应可以从腐胺得到精胺和亚精胺。鉴于果蔗方面鲜见多胺抗冷(寒)性效果的报道,另外考虑到甘蔗中95 %以上的干物质来源于光合作用,因此本研究侧重于拔地拉低温光合对外源腐胺的响应研究。【拟解决的关键问题】探寻低温条件下外施腐胺对拔地拉光合作用的作用效果和机制,为C4植物低温光合抗性研究提供新资料,为果蔗生长调节物质选用提供新参考,同时也为果蔗抗性栽培和育种措施的制定提供理论借鉴。
1 材料与方法
1.1 供试材料与试验设计
以黑皮果蔗拔地拉为材料,选取中上部芽体完好茎段,单芽种植于塑料花盆中(直径15 cm),再置于培养箱内28 ℃ /20 ℃(白天/夜间),85 %相对湿度下萌发出苗。待所有幼苗第1片完全叶长出后开启人工白光光源,叶片表面平均光量子通量密度(PPFD)为700 μmol m-2s-1,光周期11 h,温度调节为25 ℃/20 ℃,相对湿度下调至70 %。至所有幼苗均长出4片完全展开叶时分为3组,每组5~10株。第1组为对照,一直保持在上述环境条件下继续生长8 d;第2组为低温处理组,在7 ℃/4 ℃生长4 d,其他环境条件同上,然后缓慢升温至25 ℃/20 ℃生长4 d,其他环境条件同上;第3组为腐胺结合低温处理,即在低温处理前用1 mmol L-1腐胺喷施于拔地拉苗叶片上,以叶片上下表面水滴均匀附着不下滴为度,转入低温处理时生长条件同第2组。此外,在第3组喷施腐胺的同时,第1~2组需喷施清水处理。上述处理后的第5和8天,每组选取长势均匀的植株3~5株测定+ 1叶的光合参数。整个试验从头重复2次,数据取2次重复的平均值。
1.2 光合气体交换和叶绿素荧光参数测定
蔗苗经常温和冷适应处理后,再经过一个夜晚的暗适应,于第二天早上5: 30和8: 00用于测定。早上5: 30 用2台LI-6400 (LI-Cor公司,美国) 光合测定系统仪 (带LCF 荧光探头,LI-6400-40) 测定最小初始荧光Fo和最大荧光Fm等参数,仪器自动计算Fv/Fm。早上8: 00 对照植株叶片在LED植物冷光源 (上海三澎机电有限公司) (PPFD,2000 μmol m-2s-1,红光/蓝光小灯数量组成为8∶1) 下活化25~30 min,低温适应处理植株活化45~75 min,气孔导度稳定后测定荧光光响应曲线和Ci-Pn响应曲线。光合气体交换参数和荧光参数同时测量。所用PPFD 梯度为2000、1600、1200、800、500、300、150、100、50、25 和0 μmol m-2s-1;最小等待时间为2 min,最大等待时间为5 min;叶室测定温度分别控制为(20 ± 1)℃;蓝光吸收系数为0.92,红光吸收系数0.87。Ci-Pn响应曲线CO2浓度梯度为400、300、200、100、50、400、400、600、800、1000和1200 μmol L-1,光照条件为2000 μmol m-2s-1,温度条件同上。测定完毕后,仪器自动记录的部分荧光参数需重新计算。PSⅡ实际电子传递量子效率ΦPSⅡ= (Fm′-Fs)/Fm′,公式中Fm′和Fs均为仪器测定值[17];碳同化量子效率ΦCO2= (Pn-Rd)/(PPFD·а)[18],其中,Pn为净光合速率,Rd为光下暗呼吸速率,其值为光响应曲线和纵坐标的交叉点之值,а为吸光系数,其值取0.89(用叶绿素荧光成像系统大批量测定所获平均值);光化学淬灭系数qP=(Fm′-Fs)/(Fm′-Fo′) ,其中,Fm′、Fs和Fo′ 均为仪器测定值[19]。
1.3 数据处理
采用Microsoft Excel 2013和Sigmaplot 12.5软件进行数据处理及作图。
2 结果与分析
2.1 低温及其结合腐胺处理对拔地拉最大(潜在)光化学效率的影响
低温处理前3组拔地拉均表现出稳定的最大光化学效率 (Fv/Fm) 值,无光抑制迹象(图1)。和常温对照组相比,低温处理3 d后,低温处理组和低温结合腐胺处理组Fv/Fm急剧下降,表现出明显的光抑制,二者Fv/Fm的平均值虽有明显差异,但低温处理组标准差较大,经数理统计分析,低温处理组和低温结合腐胺处理组Fv/Fm值没有显著差异。低温处理结束后逐步回升至常温条件,经4 d适应后低温处理组及低温结合腐胺处理组Fv/Fm值基本恢复到对照组的水平,三者无显著差异。
2.2 低温及其结合腐胺处理对拔地拉光响应曲线特征的影响
常温下,拔地拉幼苗光响应曲线显示出典型的C4光合特性 (图2),没有明显的光饱和点,光补偿点低。从发育的角度看,成熟的+1叶经过8 d的生长,其光合净同化能力 (Pn) 没有明显变化,表明其光合特性已经相对稳定。低温处理后,叶片Pn大幅下降,光响应曲线趋于平缓,光补偿点降低 (数值未显示),冷敏性明显。在低光下,低温结合腐胺处理和低温单独处理相比,二者光响应曲线没有明显差别;在高光下,虽然两者Pn绝对值差别不大,但后者随光强梯度变化趋势明显优于前者,显示出更强的高光耐受能力。低温处理的蔗株经4 d常温恢复,Pn值和对照相比,仍有一定差别,部分光合机构很可能已经受到破坏,仍未完全修复;和低温单独处理相比,腐胺结合低温处理Pn恢复状况优势明显 (图2)。
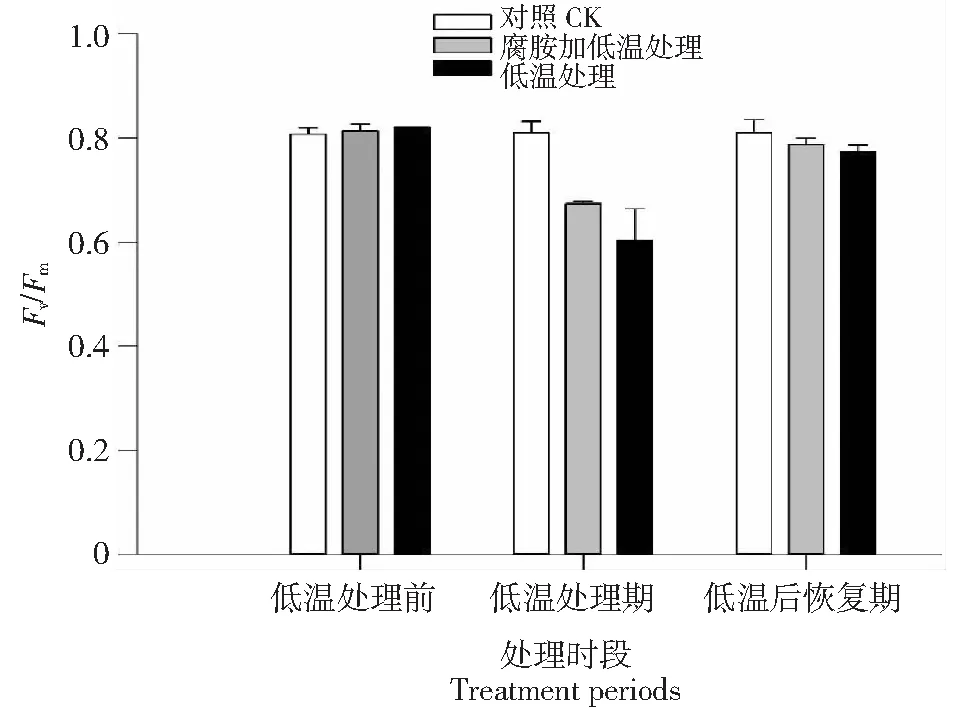
图1 常温、4 ℃低温处理4 d及常温恢复4 d条件下黑皮果蔗拔地拉凌晨最大光化学量子效率变化状况Fig.1 Changes of predawn maximal photochemical efficiency of PSII (Fv/Fm) in the chewing cane Badila under normal, chilling stress and recovery from chill conditions for detail see materials and methods
2.3 低温及其结合腐胺处理对拔地拉Ci-Pn响应曲线特征的影响
和光响应曲线变化趋势相似,常温下拔地拉叶片Ci-Pn响应曲线表现出典型的C4植物特征,CO2补偿点低 (图3),表明其光呼吸微弱。发育对叶片Ci-Pn响应曲线的影响不明显。和对照相比,低温处理后CO2补偿点略有提高 (数值未显示),但CO2饱和点大幅下降,超过CO2补偿点后的各Ci梯度下Pn值均大幅下降;腐胺结合低温处理和低温单独处理相比,叶片Ci-Pn响应曲线特征相似,CO2补偿点差别不大,CO2饱和点略有提高,Pn值有一定程度的增加,但差异不显著。经4 d常温恢复处理,低温单独处理和腐胺结合低温处理蔗株CO2饱和点和对照比仍有一定差别,腐胺结合低温处理比低温单独处理恢复状况更好。
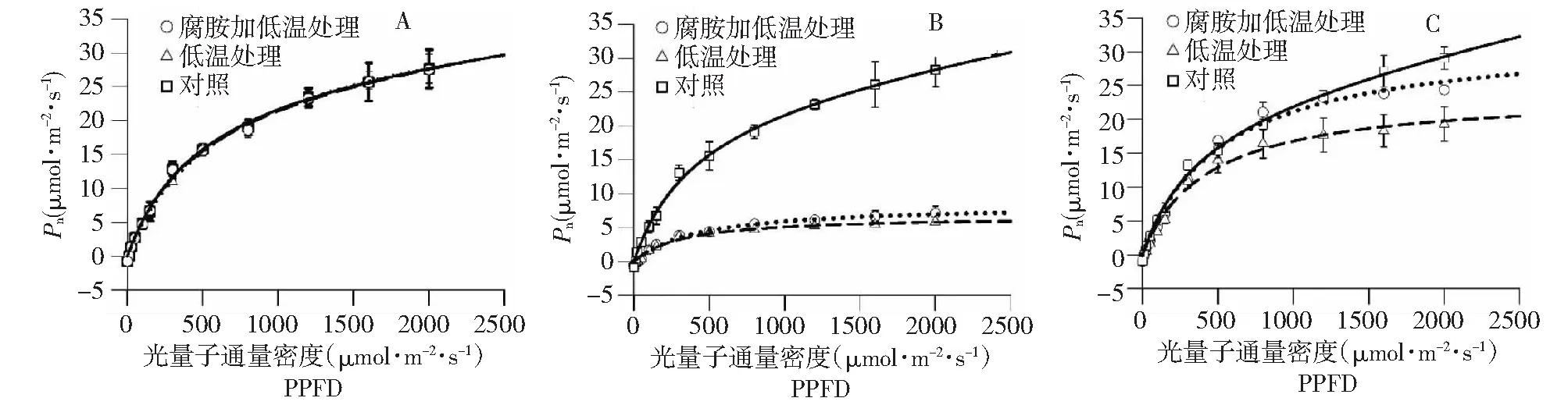
A:常温下;B低温下;C:低温结束后恢复,下同A. Normal condition; B. Chilling stress; C. Recovery from chilling stress, the same as below图2 不同处理条件下拔地拉光响应曲线的变化状况Fig.2 Changes in the light curves in the chewing cane Badila under different conditions
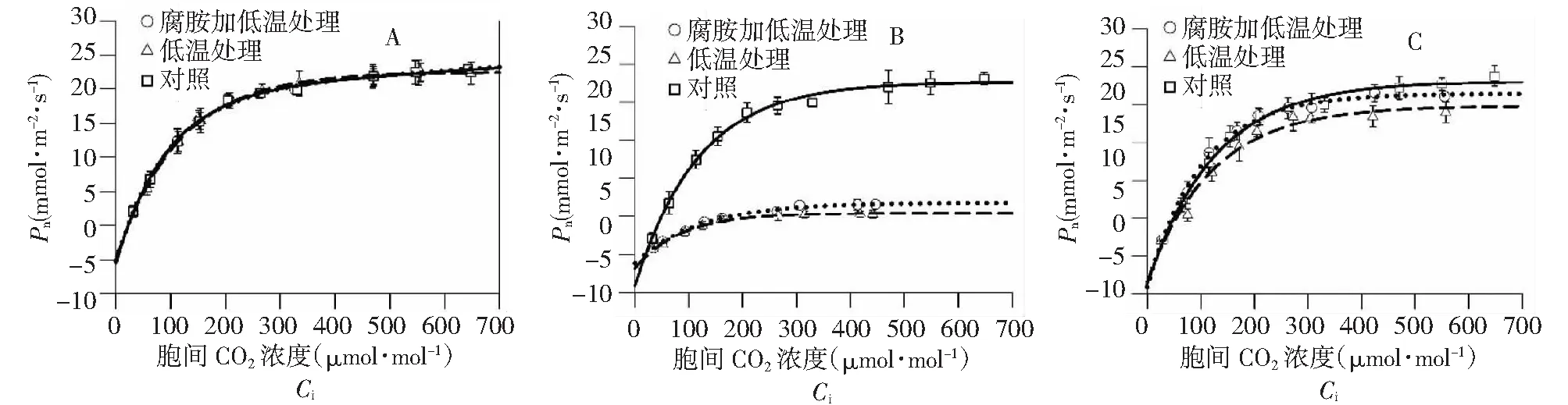
图3 不同处理条件下拔地拉胞间CO2响应曲线的变换状况Fig.3 Changes in the Ci-Pn curves in the chewing cane Badila under different conditions
2.4 低温及其结合腐胺处理对拔地拉ΦPSII光响应曲线特征的影响
光系统II电子传递量子效率ΦPSII值在常温下随光强的增加呈指数下降趋势,其值在8 d的叶片发育中变化较小 (图4)。4 ℃低温处理后4 d后,即使在低光条件下,ΦPSII值也出现大幅下降,在高光下逐渐下降至接近0;腐胺结合低温处理与低温单独处理蔗株叶片ΦPSII值没有明显差异。经过4 d常温下的恢复后,腐结合低温处理与低温单独处理蔗株叶片ΦPSII值均接近对照值,前者略优于后者。
2.5 低温及其结合腐胺处理对拔地拉NPQ光响应曲线特征的影响
叶片NPQ值在常温下随光强的增加呈幂函数增加,8 d的发育对其影响不大 (图5)。与对照相比,低温处理后,各个光强梯度下NPQ值均急剧增加,对耗散叶片多余激发能可能起了重要作用;与低温单独处理相比,腐胺结合低温处理蔗叶NPQ在低光阶段上升缓慢,但在高光下则增幅明显。经4 d常温恢复后,低温处理和腐胺结合低温处理蔗叶NPQ均回落,但仍明显高于对照,腐胺结合低温处理的回落幅度更大 (图5)。

图4 不同处理条件下拔地拉光系统II电子传递量子效率响应曲线的变换状况Fig.4 Changes in the ΦPSII-PPFD curves in the chewing cane Badila under different conditions
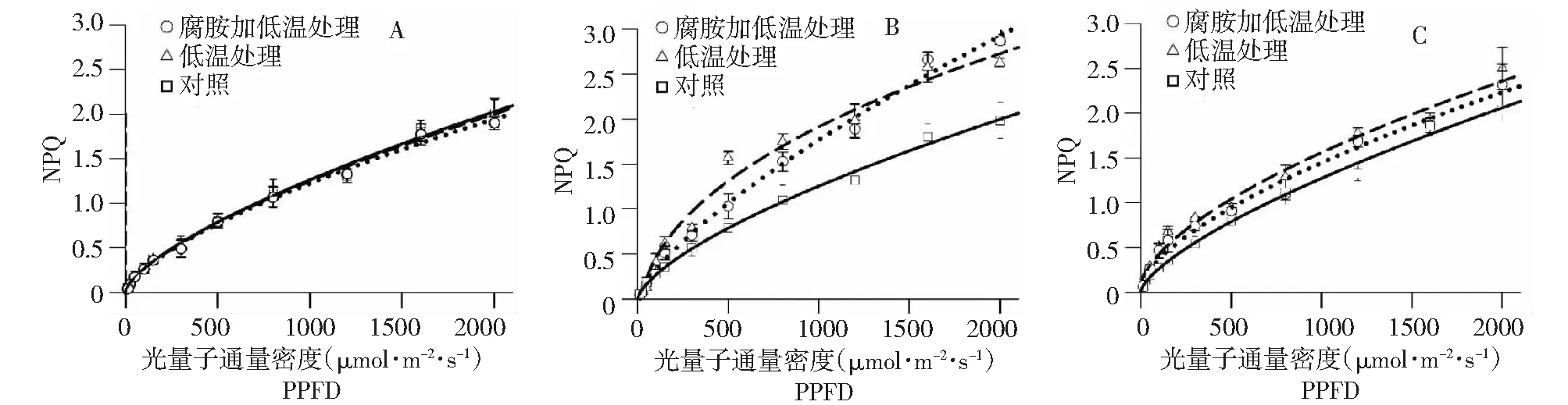
图5 不同处理条件下拔地拉NPQ响应曲线的变换状况Fig.5 Changes in the NPQ-PPFD curves in the chewing cane Badila under different conditions
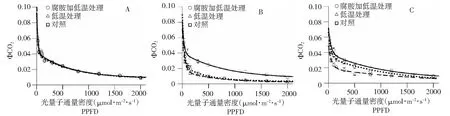
图6 不同处理条件下拔地拉CO2量子效率响应曲线的变换状况Fig.6 Changes in the ΦCO2-PPFD curves in the chewing cane Badila under different conditions
2.6 低温及其结合腐胺处理对拔地拉ΦCO2响应曲线特征的影响
拔地拉叶片CO2同化量子效率ΦCO2常温下随光强的增加呈指数衰减(图6),和ΦPSII的变化趋势相似(图4),同样,发育对ΦCO2值的影响不明显。低温处理后,ΦCO2值也是大幅下降;腐胺结合低温处理与低温单独处理相比,ΦCO2值差异不明显。常温恢复4 d处理后,低温单独处理和腐胺结合低温处理叶片ΦCO2值没有明显差异,两者均接近完全恢复状态。
3 讨 论
从低温处理前后光合气体交换参数值和叶绿素荧光参数值的变化状况看,拔地拉表现出明显的光合冷敏性。如前所述,拔地拉是热起源原种,没有抗冷的遗传基础,其光合机构也未能幸免。国内外已有关于冷敏植物光合特性的报道,低温可以影响光合机构从光能吸收转化、电子传递、碳同化到光合产物输出的所有过程[17-19],本研究发现低温下拔地拉的光化学效率、电子传递量子效率及碳同化能力都有所下降,与前人试验结果相符。进一步分析发现,低温下拔地拉光补偿点降低,表明其利用弱光的能力增强,而从光化学效率大幅下降和低温解除经一段时间恢复后光响应曲线未完全复原的情况看,光合机构的天线系统及2个光系统的化学计量关系很可能有一定程度的调节,当然这还需要进一步用生物物理和生物化学的手段加以验证。从低温前后Ci-Pn曲线的变化状况分析,低温严重削弱了Rubisco酶的羧化效率,对RuBP的再生影响也较大,而对磷再生限制的影响较小。从这些结果看,显然低温影响到光合酶的活性。也有关于其它一些冷敏感植物在低温下光合酶活性急剧降低的报道[20-22],这是其低温光抑制产生的一个重要原因。如甘蔗在干旱等逆境胁迫下光抑制造成光系统损伤[23],低温光抑制对拔地拉极可能有类似影响。拔地拉为了应付低温光抑制,其NPQ值在低温各个光梯度下都大幅增加,弥补了低活性的光化学途径耗散光合机构多余激发能较少的不足,这个结果也进一步印证了前述低温下光合机构结构调整的假设。
与单独低温处理的结果相对比,腐胺结合低温处理除了对ФPSII影响很小外,对其它光合参数值都有不同程度的提升,表明外源腐胺可增强拔地拉对低温的抗性。分析这些结果,不难看出,低温下腐胺的影响主要表现在碳同化方面,而不在电子传递上,该结果与其他植物中降低电子传递速率的报道相反[24],可能与植物种类的特异性有关,需进一步研究。有研究指出,逆境条件下,多胺可以和部分光合酶如Rubisco酶结合后增强其稳定性[25],这极可能是腐胺在低温下维持和提升甘蔗碳同化效率的主要机制。也有报道指出,逆境下多胺可以维持抗氧化酶的稳定,增加抗氧化酶活性[26],间接增进光合效率。从本研究结果中电子传递量子效率和CO2同化量子效率的变化趋势看,这种方式可能不是果蔗中腐胺作用的主要机制。
腐胺结合低温处理对拔地拉最显著的效果是促进了低温解除后光合效能的及时恢复,其它植物中类似报道还较少。从机制上分析,低温期间腐胺对光合器官及其组织结构完整性的维护,对光合酶活性的稳定等都有利于光合效能的保持,因此在低温解除后具有相对光合优势。此外,多胺是否对低温期间光合机构的渗透调节和气孔开闭也起到一定的作用还有待进一步验证。低温期间腐胺处理对电子传递和光能耗散作用并不明显,因此对急剧低温和霜冻后强光引起的低温光抑制效果可能并不理想,而在冬季自然温度逐步降低时使用腐胺效果可能更好,甘蔗生产中使用多胺时应根据不同的低温情况进行综合考虑。
4 结 论
外施腐胺对拔地拉低温光合性能有一定程度的稳定作用,且能够明显增强其碳同化能力,更重要的是,外施腐胺可以明显加速低温解除后拔地拉光合性能的恢复。该研究结果对果蔗栽培措施的制定和栽培策略的选择有理论参考价值,对果蔗低温抗性能力的提高提供了一种有潜力的参考调控方式。
猜你喜欢
出版人(2022年8期)2022-08-23
军事文摘(2021年16期)2021-11-05
装备制造技术(2020年1期)2020-12-25
今日农业(2020年23期)2020-12-15
英语文摘(2020年6期)2020-09-21
小星星·阅读100分(低年级)(2019年10期)2019-11-26
劳动保护(2018年8期)2018-09-12
山东工业技术(2016年15期)2016-12-01
Coco薇(2015年10期)2015-10-19
航空学报(2015年4期)2015-05-07
