
紫苏花器官发生及发育的形态观察
2018-12-05 07:50王曼君
江苏农业科学 2018年21期
陈 娟, 王曼君, 项 俊
(黄冈师范学院生命科学学院/经济林木种质改良与资源综合利用湖北省重点实验室/大别山特色资源开发湖北省协同创新中心,湖北黄冈 438000)
紫苏[Perillafrutescens(L.) Britt.]是唇形科1年生草本植物,原产于东亚地区,因含有丰富的α-亚麻酸,而作为一种新型的油料作物成为研究的热点[1]。然而,从文献来看,目前的研究热点主要集中在紫苏的化学成分及营养和药用价值方面[2-9],对紫苏生长发育特性,尤其是花芽分化及花器官发育的研究很少。沈宗根等对栽培紫苏成熟花的解剖结构进行了描述,但对其花芽分化过程并未涉及[10]。梅虎等从生理角度研究了紫苏花芽分化期间核酸、内源激素以及营养物质含量的变化,但并未对花芽形态分化过程作出研究[1]。Dwivedi对紫苏作了较全面的胚胎学研究,但仍缺乏对各轮花器官的发生过程的报道[11]。植物花器官的发生规律及排列方式等在植物分类地位中具有重要意义[12],本试验通过扫描电子显微镜技术详细观察了各轮花器官原基的发生过程,旨在为紫苏杂交育种提供一定理论基础。
1 材料与方法
1.1 试验材料
选取湖北省大别山上野外采摘的紫苏种子,种植在黄冈师范学院教学研究实验基地。播种前施有机肥45 t/hm2、尿素150 kg/hm2、过磷酸钙600 kg/hm2、磷二铵300 kg/hm2,并于2016年3月28日播种,行距25 cm,株距30 cm。
1.2 紫苏花发育过程观察
从花芽肉眼可见开始,每周取样1次,采集紫苏花序,迅速置于FAA固定液中,用于石蜡切片制作观察和环境扫描电子显微镜观察。
1.2.1 石蜡切片的制作与观察 对经FAA固定液固定24 h后的紫苏花序进行乙醇梯度脱水,二甲苯逐级透明,石蜡包埋,Leica RM2145半薄切片机切片,苏木精-伊红染色,Nikon eclipse E200光学显微镜下内部结构,并用Tiger ProMedia系统摄像。
1.2.2 扫描电镜(SEM)观察的样品制作 取FAA固定液的紫苏花序,在解剖镜下剥去苞叶,暴露其须观察的部分,以乙醇和乙酸异戊酯梯度脱水。CO2临界点干燥,黏样,真空喷金镀膜。在电子扫描显微镜下观察并照相。
2 结果与分析
2.1 紫苏花序的形态
紫苏花序顶生。随着紫苏叶片数量的基本确定,茎尖生长点开始由营养生长转入生殖生长期,首先是顶端突起,然后突起部分不断地伸长,花序逐渐形成。
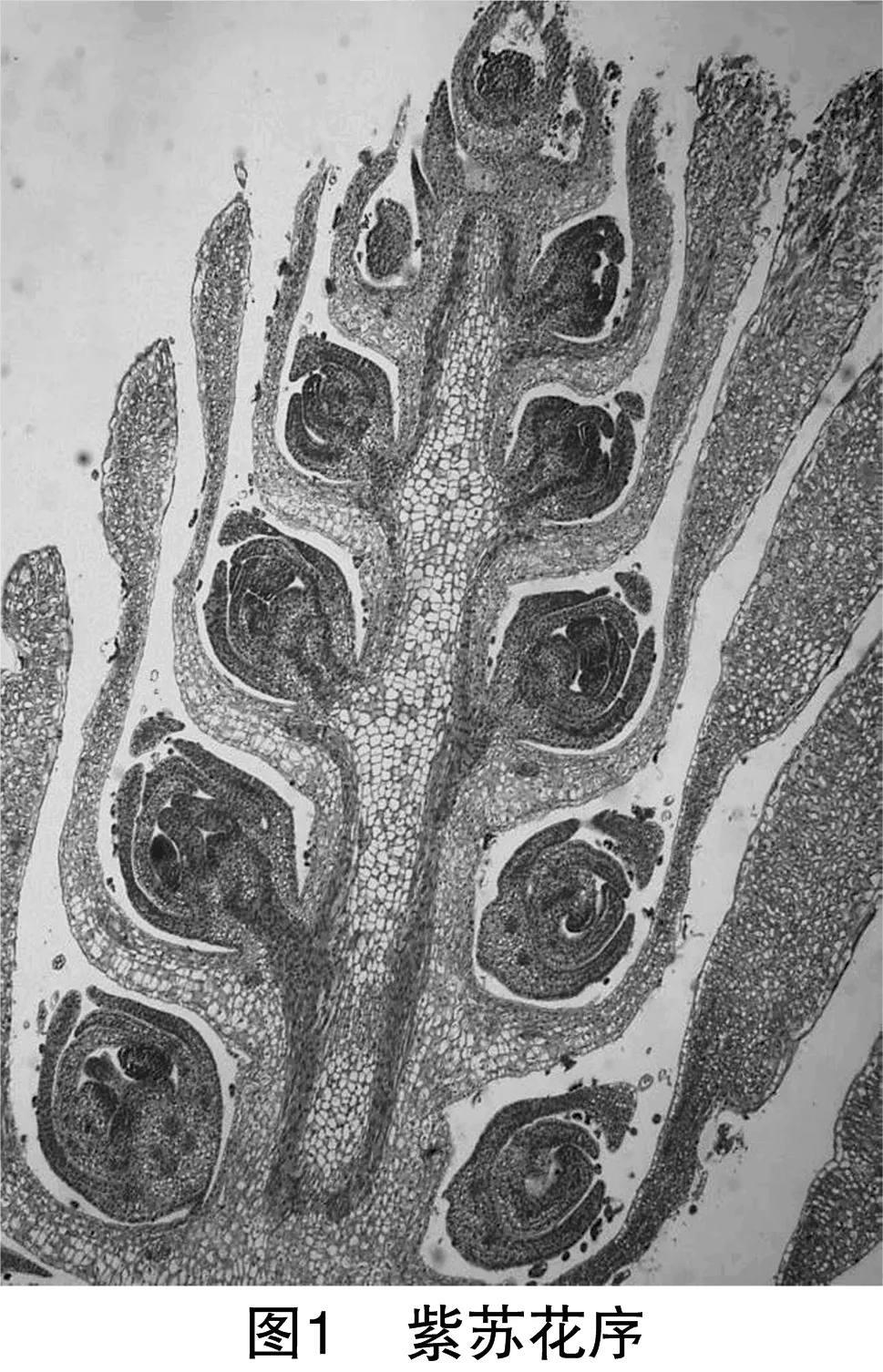
紫苏花在花序轴上呈轮伞状排列,并沿花轴由下及上方向形成无限花序,这种由下而上的排列方式类似于总状花序,顶生兼腋生(图1)。每朵花下方都有1个苞片,不同部位的花分化程度存在差异。
2.2 花器官的分化
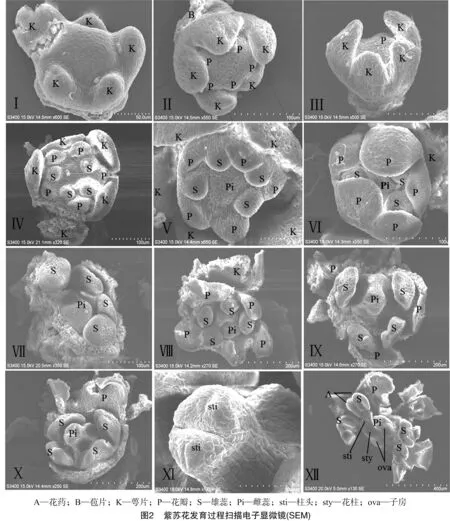
紫苏花为完全花,各花器官的分化次序为:苞叶原基形成期、萼片原基形成期、花瓣原基形成期、雄蕊原基形成期、雌蕊原基形成期(图2)。
2.2.1 苞叶原基形成期 紫苏生长进入生殖阶段时,茎尖逐渐膨大为圆凸的花序原基,花序原基不断向上伸长,并在周围形成小突起,此为苞叶原基(B)(图1、图2),此后,苞叶原基明显伸长并向内弯曲形成环抱状,此时,花序轴也明显伸长,并伴随着新的苞叶原基形成和伸长,整个花序不断伸长和变粗,形成类似总状花序的结构。
2.2.2 萼片原基形成期 随着苞叶原基的不断伸长,苞叶内侧形成1个小突起,此为花蕾原基,呈球形,顶部圆凸。花蕾原基不断增大,并其边缘产生5个小突起,此为萼片原基(K)(图2-Ⅰ)。5枚萼片原基发生同步,呈等五边形排列。接着这5枚萼片原基通过顶端生长和边缘生长,导致向上伸长和面积不断扩大(图2-Ⅱ至图2-Ⅳ)。
2.2.3 花瓣原基形成期 在萼片原基不断发育的过程中,花蕾原基顶部逐渐由圆凸转为扁平,此时在每2个萼片原基相邻处内侧的表面开始形成突起,此为花冠原基(P),共5枚。这5枚花冠原基与萼片原基一样,发生同步,呈等五边形排列(图2-Ⅱ)。
2.2.4 雄蕊原基形成期 在花瓣原基出现后不久,在其中4枚花瓣原基内侧的两两相邻处又出现4个小突起,此为雄蕊原基(S,图2-Ⅳ),4枚雄蕊原基呈类似于等腰梯形排列分布。随着这4枚雄蕊原基的不断发育和生长,逐渐出现分化,其中一对雄蕊生长速度明显快于另一对,即为紫苏的二强雄蕊(图2-Ⅶ,图2-Ⅷ)。
2.2.5 雌蕊原基形成期 雄蕊原基发育后不久,花蕾原基的最中心处即雄蕊原基内侧也逐渐隆起,形成雌蕊原基(Pi)(图2-Ⅴ)。此时,快速生长的5枚萼片的边缘已紧密愈合,将其他原基包住。随着雌蕊原基的不断伸长,花冠原基边缘基部也逐渐愈合,且花瓣的高度也超过雄蕊和雌蕊,并向中心方向伸展,逐渐覆盖雌雄蕊。
2.3 紫苏花器官原基的生长
随着各花器官原基的进一步生长,花萼最终发育成钟状,上唇3裂,下唇2裂;花冠成管状,上唇2浅裂,下唇3裂;雄蕊2对,一对相对于另一对稍长;花粉囊4室,微微分叉与花丝相连;子房上位,2个心皮,随着雌蕊原基的继续生长,深裂成为4室(ova)(图2-Ⅻ),花蕾原基中间突起部分继续向上伸长,形成花柱(sty)(图2-Ⅻ),柱头(sti)二裂(图2-Ⅺ)。
3 讨论与结论
紫苏花为两性完全花。一般认为,被子植物花器官具有向心发育的特性,即距离顶端最远的花器官分化发育最早,依次向内逐渐分化发育,但也存在一些植物的花器官具有离心生长的特性,如红木科(Bixaceae)、椴树科(Tiliaceae),可以发现花顶端停止生长,雄蕊原基向基或离心生长的现象[13-14]。少数植物中存在向心发育的现象如耧斗菜属(Aquilegia),雄蕊的发育是向心的,但是小孢子的发育是离心的,即离顶端近的花药发育更快[15],而马桑绣球(Hydrangeaaspera)的花萼和花瓣几乎同时发生,雄蕊和雌蕊也几乎同时发生[16]。南天竹(NandinadomesticaThunb.)的雄蕊和花瓣是经它们所具有的共同原基进行侧向分裂形成的[15]。欧洲油菜(Brassicanapus)的花器官发生顺序依次是花萼、雄蕊群、花瓣、雌蕊群,与一般的向心式发育有差别[17]。本研究发现,紫苏花器官的分化成花过程符合大多数被子植物花器官分化的一般规律,即花器官原基出现的顺序依次为萼片原基、花冠原基、雄蕊原基、雌蕊原基。
植物花器官在花托上的排列顺序有多种:螺旋式排列,如木兰科植物的花,这种花称为旋生花(spiral flower);轮状排列,即各轮花器官发生在花托的同一个或接近同一个水平上,一般大多数植物的花都是这种排列方式,称为轮生花(cyclic flower);另外还有一种半轮生花,花被轮生,雌雄蕊旋生,如某些毛茛科植物的花。在原始的被子植物花器官中,螺旋式和轮式排列方式共存,无一定秩序,而在较为进化的被子植物中,花器官数目逐渐减少并趋于定数,出现了花器官之间的合生。Endress认为,轮生排列是花器官数目减少而且固定的先决条件,也是合瓣、合生心皮以及雌蕊与其他器官合生的先决条件[18]。本研究表明,紫苏各轮花器官之间的排列顺序为轮生排列,这在各轮花器官原基发生时就已以发生的先后顺序表现出来。
被子植物花芽分化时期有长有短,分化的时期也不一致,如山毛榉科的板栗雄花序从第1年6—8月便出现雄花序原基,至来年3月才分化成熟,开始开花[19],可谓历时很长,而紫薇科的黄金树花芽分化则集中在3月下旬至5月上旬之间[20],可谓历时较短。笔者观察到紫苏整个花器官的分化期处在7月中旬至9月初,相比而言,分化速度较快,分化时期较短。植物花芽分化生理活动旺盛,须消耗大量养分,紫苏作为一种重要的油料作物,此阶段适时进行花期管理,及时施以速效肥,可以补充养分供应,促进花芽分化,促使其多开花结实,获得高产。
有些文献表明,紫苏为自花授粉植物。本研究也发现,虽然紫苏雌雄蕊原基的分化产生不同步,雄蕊原基的分化早于雌蕊原基,但在随后的发育过程中,雌雄蕊的发育又逐渐趋于同步。而张大勇曾指出,植物雌雄蕊成熟的特点在一定程度上影响植物的授粉方式,如雌雄异熟可以避免自花授粉;雌雄蕊在空间上的分离可以使植物更倾向于异化授粉等[21]。紫苏雌雄蕊同步成熟可以保证其自花授粉。然而,紫苏雄蕊为二强雄蕊,其特点符合唇形科植物虫媒授粉的特性[22-25];另外,紫苏作为一种芳香植物,其分泌出的独特香气也具有吸引昆虫帮其传粉的功能,根据沈宗根的研究显示,紫苏花蜜腺的泌蜜量丰富,诱蜂力强[24,26]。因此,笔者认为,紫苏应该属于自花兼具异花授粉的植物,这样既可以通过异花授粉保证其后代的质量,又可以避免不良环境造成虫媒授粉不良时,通过自花授粉保证后代的数量。
紫苏二强雄蕊原基与另外1对短雄蕊原基发生时间一致,产生长短的分化是在这2对雄蕊原基发育一段时间之后,其机理有待进一步研究。
猜你喜欢
经济林研究(2022年2期)2022-07-20
课外生活·趣知识(2022年6期)2022-07-14
现代农业科技(2022年4期)2022-03-08
锦绣·上旬刊(2020年10期)2020-12-14
中学课程辅导·教师教育(上、下)(2018年15期)2018-10-10
山西农业科学(2018年9期)2018-09-12
生物学教学(2017年6期)2017-02-18
中国马铃薯(2015年3期)2015-12-19
华南师范大学学报(自然科学版)(2014年6期)2014-08-28
