
毛蕊花糖苷对血小板衍生生长因子BB处理后大鼠肝星状细胞的影响
2018-12-07 03:16由淑萍张石蕾军马龙刘
癌变·畸变·突变 2018年6期
由淑萍 张石蕾 赵 军马 龙刘 涛*
(1. 新 疆医科大学公共卫生学院,新疆 乌鲁木齐830011;2. 新疆医科大学护理学院,新疆乌鲁木齐 830011;3. 新 疆维吾尔自治区药物研究所,新疆 乌鲁木齐 830004)
肝纤维化发生与发展的核心环节是肝星状细胞(hepatic stellate cells,HSC)激活后的迅速增殖,分泌过量的细胞外基质(extracellular matrix,ECM)。本课题组前期研究结果表明[1-3],肉苁蓉苯乙醇总苷(phenylethanol glycosides from Cistanche tubulosa,CPhGs)对牛血清白蛋白(bovine serum albumin,BSA)所致的肝纤维化大鼠及HSC细胞的活化均具有显著抑制作用,呈现出一定的抗肝纤维化及抗炎的生物活性。由新疆维吾尔自治区药物研究所植物化学室提取纯化的CPhGs(纯度为70%),经鉴定其主要成分为毛蕊花糖苷和松果菊苷,且毛蕊花糖苷诱导HSC细胞的凋亡最为显著[4]。同时,文献研究显示,血小板衍生生长因子(platelet derived growth factor,PDGF)是刺激HSC细胞最强的有丝分裂原,它有3种不同形式的二聚体:PDGF-AA、PDGFBB和PDGF-AB。目前认为,PDGF-BB是刺激HSC细胞活化,激活相关基因及蛋白的表达,对肝纤维化的形成最为有效的促有丝分裂因子。毛蕊花糖苷是否能够参与PDGF-BB因子诱导的肝纤维化并调节及抑制HSC细胞的活化仍然值得继续深入研究。本研究拟采用重组大鼠血小板衍生生长因子-BB(recombinant rat platelet derived growth factor-BB, rrPDGF-BB)刺 激HSC细胞,观察不同浓度的毛蕊花糖苷对PDGF-BB因子诱导的HSC细胞增殖的抑制作用,及其对HSC细胞迁移能力和肝纤维化形成过程中的标志性基因表达的影响,如HSC细胞活化标志基因α-SMA、凋亡效应分子caspase-3、最重要的促增殖和凋亡的MAPK信号通路中的关键基因ERK1/2、P-ERK1/2、Akt、P-Akt等,以探讨毛蕊花糖苷对HSC细胞的调控作用及可能的作用机制,以期为抑制HSC细胞活化、寻找防止肝纤维化的新靶点提供实验依据。
1 材料与方法
1.1 材料与试剂
肉苁蓉苯乙醇总苷及毛蕊花糖苷,由新疆和田帝辰生物医药有限公司提供,以DMEM(高糖)完全培养液将毛蕊花糖苷配置成3种混悬液,浓度分别为1.5、3.0、6.0 mg/L;HSC细胞购自中国武汉普诺赛(Procell)公司;大鼠Ⅰ型胶原ELISA试剂盒(CUSABIO)购自武汉华美生物工程有限公司;重组大鼠PDGF-BB(rrPDGFBB)购自美国Peprotech公司;p44/42 MAPK(Erk1/2)、phospho-p44/42 MAPK(p-Erk1/2)、 Akt、 P-Akt抗 体 购自美国CST公司;α-SMA抗体购自美国Abam公司;caspase-3抗体、β-actin抗体购自中国武汉三鹰(Proteintech)公司。
1.2 HSC细胞培养及分组
HSC细胞分为5组[4]:对照组、PDGF-BB组、毛蕊花 糖 苷 (1.5、 3.0、 6.0 mg/L)组 。 将HSC细 胞 以 5×104/mL接种于6孔板,含10%胎牛血清DMEM高糖培养基培养24 h后,对照组不施加任何干预,PDGF-BB组仅用rrPDGF-BB刺激(10 ng/mL),作用24 h。3个毛蕊花糖苷组均加入rrPDGF-BB(10 ng/mL)刺激24 h,再分别加入(1.5、3.0、6.0 mg/L)的毛蕊花糖苷处理。
1.3 划痕实验检测毛蕊花糖苷对HSC迁移能力的影响
将消毒好的直尺置于6孔板上,用200 μL无菌枪头沿直尺在培养板底部划3条平行直线,分别于培养0、24 h后采集图像,拍照图像采集点尽量保持前后一致。Image J分析细胞迁移距离,划痕宽度用l表示,迁移率=(l0h- l24h) /l0h×100%。
1.4 ELISA法检测Ⅰ型胶原(Col-I)含量
采用ELISA法定量测定不同组别的细胞培养上清液中Col-I含量。用大鼠Col-I抗体包被微孔板,制成固相载体,依次加入标准品、生物素化的抗Col-I抗体、HRP标记的亲和素,TMB显色(TMB在过氧化物酶的催化下转化成蓝色,并在酸的作用下转化成最终的黄色)。颜色的深浅和样本中的Col-I呈正相关。用酶标仪下测定吸光度D(450)值,并计算样本浓度。
1.5 Western blot法 检 测 α-SMA、 caspase 3、ERK1/2、P-ERK1/2、Akt、P-Akt蛋白表达
HSC细胞培养、分组同上,48 h后收集细胞并分别提取各组细胞总蛋白,BCA法测定蛋白含量。取各组细胞总蛋白20 μg进行SDS-聚丙烯酰氨凝胶电泳实验,根据目的蛋白的大小将包含目的蛋白的凝胶切下,以恒压100 V、60~90 min进行转膜,5%脱脂奶粉的1×TBST溶液封闭1 h,根据目的蛋白分别加入一抗,具体的一抗及浓度为:ERK1/2、P-ERK1/2、Akt和P-Akt均为1∶1 000,α-SMA、caspase-3和β-actin均为1∶5 000,4 ℃过夜。TBST洗膜3次,每次10 min。加入碱性磷酸酶标记的抗兔/鼠二抗(Invitrogen),孵育2 h,β-actin作为内参显色剂显影,凝胶成像系统分析结果。
1.6 统计学方法
2 结 果
2.1 毛蕊花糖苷高效液相图分析
采用高效液相法测量毛蕊花糖苷的纯度,结果显示毛蕊花糖苷纯度为98.89%,见图1。
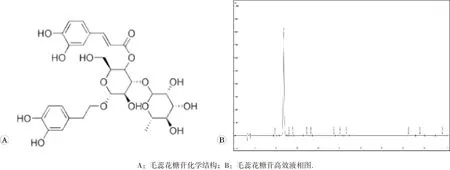
图1 毛蕊花糖苷高效液相图分析
2.2 毛蕊花糖苷对HSC迁移能力的影响
细胞划痕法检测毛蕊花糖苷对HSC迁移能力影响的结果显示,正常对照组细胞迁移率为(38.05±1.95) %,PDGF-BB组细胞为(93.16±3.28)%,可见与对照组比较,PDGF-BB能够促进HSC的迁移(P<0.01);毛蕊花糖苷(1.5、3.0、6.0) mg/L组细胞的迁移率分别为(33.11 ±1.20)%、(42.12±1.24)%、(66.55±1.51)%, 与PDGFBB组比较,毛蕊花糖苷1.5、3.0、6.0 mg/L组能明显抑制HSC的迁移(P<0.01)(图2);且毛蕊花糖苷抑制HSC的迁移有明显的剂量-效应关系(r=0.894,P=0.038)。

图2 毛蕊花糖苷对HSC迁移能力的影响
2.3 毛蕊花糖苷对HSC中Col-I含量的影响
肝纤维化时大量ECM在Disse间隙沉积以胶原为主,尤以Col-I为主,这些间质成分的改变不但激活HSC,也影响肝细胞的功能,因此我们通过ELISA法对Col-I的含量进行测定,结果显示:与正常对照组比较,PDGF-BB组HSC中Col-I含量增加,与PDGF-BB组比较,毛蕊花糖苷(1.5、3.0、6.0 mg/L)组Col-I含量均有一定程度的减少,差异具有统计学意义(P<0.01);且随着毛蕊花糖苷浓度的增加,胶原分泌呈现逐渐降低的趋势,毛蕊花糖苷各剂量组间呈现剂量-效应关系(r=0.397,P=0.027),见图3。提示毛蕊花糖苷能够抑制HSC的ECM过度沉积及胶原的形成。
2.4 毛蕊花糖苷对HSC活化标志物α-SMA蛋白表达的影响
α-SMA作为HSC激活的标志,因此我们通过Western blot法对α-SMA蛋白表达进行测定,结果发现:rrPDGF-BB作用于HSC细胞后,α-SMA蛋白的表达较对照组明显增强;与PDGF-BB组比较,毛蕊花糖苷(1.5、3.0、6.0 mg/L)组α-SMA蛋白的表达有所降低,差异具有统计学意义(P<0.01);同时,毛蕊花糖苷(1.5、3.0、6.0 mg/L)组α-SMA蛋白表达呈现剂量-效应关系(r=0.975,P<0.01),见图4。由此结果预测,毛蕊花糖苷可以抑制HSC由静止表型向表达α-SMA的肌成纤维细胞样表型转化,抑制HSC在肝损伤部位的移行和增殖。
2.5 毛蕊花糖苷对HSC凋亡效应分子caspase-3表达的影响
Caspase-3被视为最重要的凋亡效应分子,Western blot法对caspase-3蛋白表达进行测定,结果发现:与对照组比较,PDGF-BB组呈低表达;毛蕊花糖苷干预HSC后,与PDGF-BB组比较,caspase-3 蛋白的表达量升高,并且随着毛蕊花糖苷的浓度增加,其表达量逐渐升高(r=0.982,P<0.001),呈现药物剂量-效应关系。见图5,说明毛蕊花糖苷可以激活caspase-3的活性,促进caspase-3发挥凋亡执行作用。
2.6 毛蕊花糖苷对ERK1/2、Akt信号通路的影响
ERK1/2、Akt激活可促进HSC细胞增殖,PDGF激活的信号通路中ERK1/2、Akt信号通路存在着广泛的“cross talk”,具有相互协同或抑制作用。Western blot结果表明:PDGF-BB作用于HSC 48 h后可以使ERK1/2、P-ERK1/2、Akt、P-Akt蛋白表达明显增强,而毛蕊花糖苷可以明显抑制ERK1/2、Akt的活化,PERK1/2、P-Akt蛋白表达水平明显降低,且不同浓度间呈现药物剂量-效应关系(0.826 图4 毛蕊花糖苷对HSC活化标志物α-SMA蛋白表达的影响 图5 毛蕊花糖苷对HSC凋亡效应分子caspase-3表达的影响 肝纤维化时HSC是ECM过度沉积的关键细胞,并在肝纤维化的启动过程中发挥重要作用,ECM的合成累积循环变化能活化HSC[5],其中大量的ECM成分以胶原和PDGF等细胞因子为主[6];而PDGF又可进一步激活HSC,使其演化为肌成纤维母细胞并表达α-SMA,因此,诱导活化的HSC凋亡并抑制其活化、增殖是防治肝纤维化的有效途径[7-9]。 图6 毛蕊花糖苷对ERK1/2、Akt信号通路中ERK1/2、P-ERK1/2、Akt、P-Akt蛋白表达的影响 α-SMA与HSC关系密切[10-12],被广泛用于HSC活化的标志。本研究结果显示,rrPDGF-BB刺激HSC细胞48 h后,迁移能力显著增加,α-SMA蛋白的表达明显增强,同时,酶联免疫ELISA定量法检测HSC中Col-I含量也呈现明显增加趋势,表明HSC活化的过程常常伴随着侵袭能力的增强;而不同浓度的毛蕊花糖苷组干预后,与PDGF-BB组比较,HSC迁移能力明显下降,α-SMA蛋白表达明显降低,Col-I含量显著减少,证实毛蕊花糖苷可以抑制HSC的侵袭能力、减少PDGF产生、降低HSC活化、降低胶原纤维含量、阻遏肝纤维化的形成。 研究指出,通过药物防治来促进活化的HSC凋亡或将成为具有前景的抗肝纤维化手段[13]。天冬氨酸特异性半胱氨酸蛋白酶(caspase)的激活是细胞凋亡发生机制的关键元件,在级联反应中能够呈现放大效应,caspase抑制剂可逆转肝纤维化的发生。在caspase 家族介导的凋亡信号传导通路中,caspase-3是执行型caspase,处于各条通路的枢纽环节,被视为最重要的凋亡效应分子,活化后可特异性地裂解底物并切割下游的蛋白激酶、核酸酶以及细胞骨架等,诱导细胞凋亡。本研究结果显示,rrPDGF-BB诱导HSC活化后,caspase-3蛋白的表达量显著降低,毛蕊花糖苷干预后,其表达量增加,且剂量越高caspase-3蛋白的表达越高,提示毛蕊花糖苷参与了HSC细胞的凋亡,增加caspase-3的活性可促进活化的HSC凋亡。 MAPK信号通路是细胞内最重要的促增殖和凋亡通路之一[14],在HSC的增殖和凋亡中能够影响其下游细胞周期调节蛋白、凋亡相关蛋白等效应分子的活性[15],caspase-3被视为最重要的凋亡效应分子;caspase-3的活化可调控ERK1/2、Akt信号通路,这其中,PDGF-BB的作用尤为突出;PDGF-BB与受体PDGFR β偶联后诱导酪氨酸残基的自体磷酸化,引起受体相关蛋白的募集和激活,进一步激活如ERK、Akt、P38这3个MAPK家族中的主要亚群[16-17],调节细胞的凋亡、免疫反应、增殖、基因表达等多重功能。本次实验结果证实了PDGF-BB对HSC的增殖、活化及促肝纤维化作用的同时,也发现毛蕊花糖苷能使caspase-3激活,减少ERK1/2、Akt的磷酸化水平,阻断ERK1/2、Akt信号通路,进而抑制HSC中ECM过度沉积及胶原形成,减少肝纤维化的发生。 毛蕊花糖苷是一类生物活性很强的化合物,具有消炎、增强免疫、抗缺氧、清除自由基、DNA碱基修复等多种生理作用,同时,肝纤维化是由一类复杂的细胞因子和生长因子系统调节的慢性、渐进的病理过程,具有可逆性,其病理发生机制与毛蕊花糖苷的药理学防治机制具有很强的相关性。而对于肝纤维化,至今仍然没有公认的有效治疗方案,因此深入研究毛蕊花糖苷抑制HSC的迁移作用,使ECM分泌减少,促进ECM降解,抑制HSC的增殖并诱导其凋亡可为毛蕊花糖苷的抗肝纤维化作用提供更多的理论依据。 综上,本研究表明,毛蕊花糖苷可以抑制PDGFBB诱导的HSC的迁移,降低I型胶原的分泌,抑制α-SMA蛋白的表达降低HSC的活化,并且可以激活HSC最重要的凋亡效应分子caspase-3的表达,及通过抑制ERK1/2、P-ERK1/2、Akt、P-Akt蛋白的表达发挥抗肝纤维化的作用。

3 讨 论

猜你喜欢
材料与冶金学报(2022年2期)2022-08-10
中国马铃薯(2022年2期)2022-07-05
中草药(2022年11期)2022-05-31
中草药(2022年9期)2022-05-06
当代陕西(2022年5期)2022-04-19
皮革科学与工程(2022年1期)2022-01-15
科学与财富(2021年33期)2021-05-10
现代装饰(2021年1期)2021-03-29
山西农业科学(2020年11期)2020-11-19
食品工业科技(2014年13期)2014-03-11
